江苏云台山山樱花种群结构及点格局分析
2021-07-28彭智奇朱淑霞董京京钟育谦翟飞飞郑爱春王贤荣伊贤贵
彭智奇,董 鹏,朱 弘,朱淑霞,董京京,钟育谦,翟飞飞,郑爱春,王贤荣,伊贤贵*
(1.南京林业大学生物与环境学院,南方现代林业协同创新中心,樱花研究中心,江苏 南京 210037;2.江苏省野生动植物保护站,江苏 南京 210036;3.南京市浦口林业站,江苏 南京 210032)
山樱花(Cerasusserrulata),隶属蔷薇科(Rosaceae)樱属(Cerasus)[1],广泛分布于我国大部分地区的山谷林中,是暖温带森林里的常见树种[2]。山樱花是‘松月’(C.serrulatavar.lannesiana‘Superba’)、‘关山’(C.serrulatavar.lannesiana‘Kanzan’)等众多观赏樱花的亲本[3],花形秀美、花色明丽、花期长及秋叶红黄色等特征使其拥有不菲的观赏价值,其分布区域之广、适应性之强,使其近年来在国产樱属种质资源里备受关注[4-6]。
植物群落的植物社会学分析是研究任何植物的首要基础,是了解群落结构及其构成的先决条件。它能够提供关于种群结构和内外在影响、生物对群落内其他物种的变化产生的反应、种间相互作用、关键物种的演替和退化生境的生态恢复等的信息[7]。群落结构受种群结构的直接影响,群落的发展和演变趋势与种群结构息息相关[8]。种群龄级结构是种群数量随时间变化的客观反映,能界定该种群在群落中的地位和作用, 是植物种群的重要结构指标之一,当位于同一环境时,径级可以成为龄级最直观的体现[9];静态生命表和存活曲线是种群数量的动态展现,能帮助了解种群数量变化的潜在机制、阐明种群生存的客观规律[10];点格局分析和时间序列预测也是近年来研究植物种群结构与动态的常用方法[11]。群落动态反映了群落在时间、空间上的变化规律,区域尺度的群落动态研究可客观反映群落种群之间以及种群与环境之间的关系,并预测该地区种群的发展和演替趋势[12-13]。由此可见,上述方法对探索种群特性、了解生态系统结构、阐明群落演替规律、保护生物多样性都有重大意义[14]。另外,对样地内植物群落的研究不仅可以为植物区系分析提供可靠依据,也可以更好地了解种群特性与群落结构以及森林系统结构、功能和演化趋势[15-16]。
国外关于樱属的研究主要集中在组织培养、无性系繁殖、种群和生态因子间的联系及种群恢复上,以非洲李(Prunusafricana)为例,其药用价值自从被认识到以后其野生种群数量大幅减少,因此也被列入IUCN、CITES附录[17]。国外学者对其分布区域的生态特征,群落丰富度、覆盖度、生长率、死亡率和繁殖参数展开了调查,并观察其完整的生命周期找到影响种群生存的主要影响因子[18]。在1998—2006年间种群结构不同于寿命较长的植物种典型的“J型”曲线,大约有一半的个体在研究期间死亡,主要是由于过度采伐、过度放牧、火灾等人为因素。随后对村民和森林管理人员进行无性繁殖技术和苗圃实践培训,为该植物种可持续发展提供了新的可能性[19]。最近有关Prunuskorshinskyi的研究加入了种群与土壤微生物和植物光谱的联系[20],能更好地解释植物群落结构及环境的相关性、客观地揭示种群现状,但关于种群的发展趋势及保护对策的研究并不深入。国内关于山樱花的研究目前主要集中在组织培养[2,15]、种子萌发[21]、基因组[22]、遗传多样性[23]、谱系地理学[24]等方面,但是对其种群生态学的研究鲜见报道。为进一步增加对该植物种在群落中状况的了解,本研究根据1 hm2的固定样地监测数据,对云台山保护区山樱花群落结构、种群结构和点格局进行分析,通过时间序列进行预测,以期掌握该地区山樱花种群结构特征与空间分布格局,了解其在所处植物群落的地位,从而为山樱花的资源保护、引种培育以及利用等提供理论依据,同时也为云台山自然保护区地带性植被恢复与林相改造提出参考建议。
1 材料与方法
1.1 研究区自然概况
云台山(119°6′~119°30′E,34°31′~34°47′N)是泰山山脉沂蒙分脉的一支余脉,最高海拔625 m,总面积为16 676.3 m2,具有低山谷地貌,是江苏省内有代表性的主要山地之一[25]。云台山地处暖温带和北亚热带过渡区域,受海洋气候影响,又形成了类似亚热带的小气候区域,特殊的地理位置也让其植物构成明显具有南北兼容的特点。因此云台山植物种类丰富度极高,仅次于宜溧山脉,在江苏省内排名第二[25]。山区土壤为棕壤,周围平原区分布着次生盐土、盐土及滨海盐土[26-28]。最新的植物调查结果显示,云台山野生植物资源相当丰富,不乏极具地域代表性的物种和珍贵的古树名木。云台山现存植被中的优势种主要有杉木(Cunninghamialanceolata)、化香树(Platycaryastrobilacea)、青冈(Cyclobalanopsisglauca)等,构树(Broussonetiapapyrifera)、野花椒(Zanthoxylumsimulans)、刺楸(Kalopanaxseptemlobus)等次生先锋树种同样占据优势。虽然先后有野生乔灌木、蕨类、草本植物、果树、药用植物[29-34]等的群落学报道,但是对该区域代表性的物种,尤其是木本植物缺乏较全面的认识。
本研究拟通过对江苏云台山省级自然保护区山樱花种群的研究,了解其在所处植物群落的地位,掌握其种群结构特征与分布格局,为山樱花的保护与利用以及地带性植被恢复等提供理论依据。
1.2 样地设置和数据收集
在江苏云台山省级自然保护区内,典型暖温带落叶阔叶林内设置100 m×100 m的固定监测样地(119°27′36.98″E,34°43′0.67″N),如图1,总面积10 000 m2。将样地西南角设置为坐标系原点,x轴东西走向,y轴南北走向,再将样地划分为400个5 m×5 m的工作样方,并依次编号。
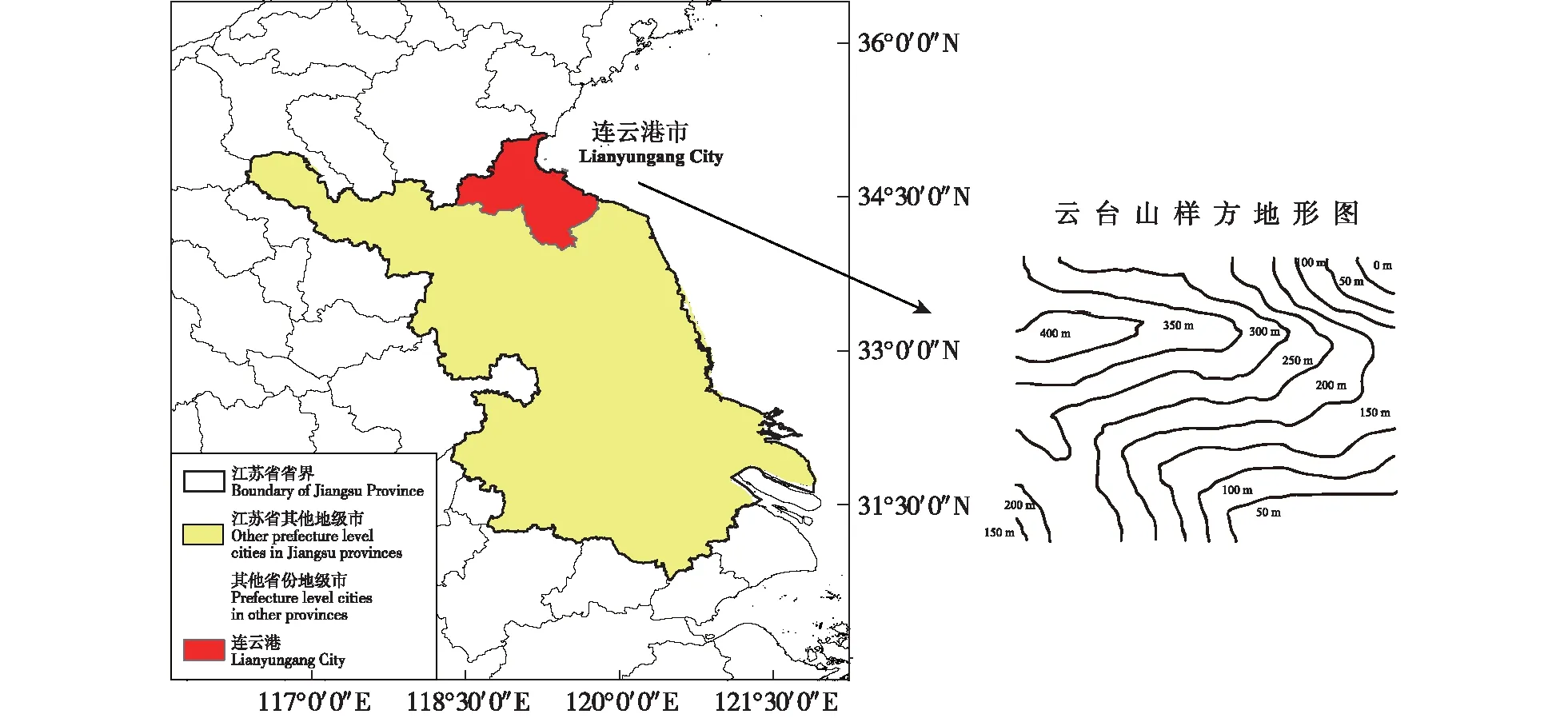
审图号:GS(2020)4632号
以小样方为调查单元,记录符合条件样本的样方编号、植物主干编号、中文名及其坐标、胸径、树高、枝下高、冠幅等指标。采用“样方法”对云台山样方中所有的群落个体进行调查:乔木层树种测量胸径、高度、株数,灌木层测量高度、株数,草本层测量高度、盖度。同时采用“实测法”调查每个样方内山樱花个体(含幼苗)的坐标、树高、胸径等,同一树桩萌发的无性系分枝记录胸径最大的一株。小样方的坐标原点也设置为西南角,小样方内植株的空间坐标用投影距离来表示。
1.3 分析方法
1)种群径级结构划分。通过汇总胸径数据,胸径在2.0 cm以下的山樱花幼苗设置为第Ⅰ径级,≥2.0~4.0 cm划定为第Ⅱ径级,以此类推,每隔2 cm为一个径级,第Ⅰ径级为幼苗,胸径不统计。该样方内山樱花最小胸径2.1 cm,最大胸径15.8 cm,最终将该种群分成8个径级,并统计各径级山樱花株数。
2)静态生命表的编制。静态生命表通过这种数量关系反映出种群在出生率、死亡率等相关方面的信息,为综合判断种群现状提供了有效的手段。生命表最关键的原始数据为ax,其他相关指标由以下函数式求得:
lx=ax/a0×1 000;dx=lx-lx +1;qx=dx/lx;Lx= (lx+lx +1)/2;
式中:x代表龄级;ax代表x到x+1龄级区间内现有个体数;lx代表x龄级初始状态标准化存活个体数;dx代表x到x+1龄级区间内标准化死亡数;qx代表x到x+1龄级间隔期死亡率;Lx代表从x到x+1龄级间隔期还存活的个体数;Tx代表从x龄级到超过x龄级的个体总数;ex代表进入x龄级平均寿命;Kx代表各龄级消失率[35-36]。
3)死亡曲线与存活曲线绘制。分别绘制山樱花种群存活曲线和死亡率、消失率曲线,以径级为横坐标,纵坐标分别为标准化的存活个体数的自然对数和死亡率[37]。
4)时间序列预测。研究采用的时间序列预测模型如下:
式中:t表示龄级;Mt为t龄级内有活的个体数;n代表预测n年后t龄级的个体数;xk代表经过n个龄级时间后未来t龄级的种群大小。本次只对未来2、4、6个龄级时间后种群各龄级的个体数量进行预测[38]。
5)点格局分析。采用R-3.6.1,在Rstudio操作平台上分析山樱花种群及各龄级在不同尺度下的点格局。通过模拟计算99%的置信区间,当g(r)值位于上下包迹线之内,符合随机分布:在上包迹线之上为聚集分布;在下包迹线之下为均匀分布[39]。
2 结果与分析
2.1 群落结构特征
乔木层和灌木层主要树种(重要值前8位)概况见表1,其中灌木层的乔木树种为该树种的小苗。统计可知,典型样地内的树种共计63种,5 301株,其中短柄枹栎(Quercusserrata)2 626株,山樱花550株,占总体的10.38%。另外,样地内有灌木39种,草本、藤本合计37种,植物多样性指数较高,其主要群落为短柄枹栎+满山红(Rhododendronmariesii)+海金沙(Lygodiumjaponicum)。
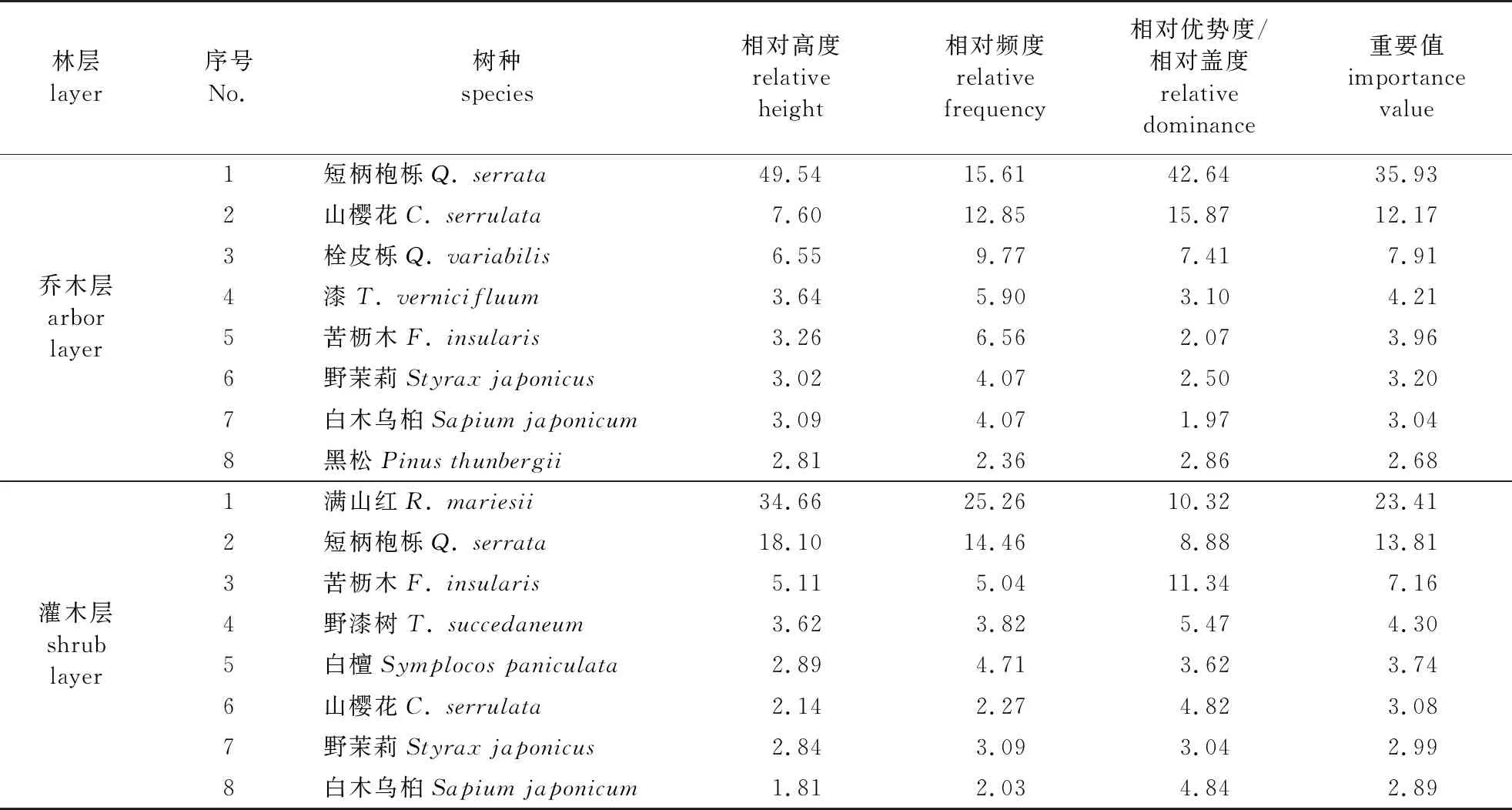
表1 群落乔木层、灌木层主要种概况
壳斗科株型高大,冠型开展,在此情况下不利于山樱花生长,但山樱花重要值仅次于短柄枹栎,是该群落重要的组成成分之一。短柄枹栎种群已在落叶阔叶林乔木层中占有绝对的优势,其重要值高达35.93,乔木层重要值>2.0的从大到小依次是短柄枹栎、栓皮栎(Q.variabilis)、山樱花、漆(Toxicodendronvernicifluum)、苦枥木(Fraxinusinsularis)、野茉莉(Styraxjaponicus)、白木乌桕(Sapiumjaponicum)、黑松(Pinusthunbergii)、化香树(P.strobilacea)、黄连木(Pistaciachinensis);灌木层重要值>2.0的从在厔小依次是满山红、短柄枹栎、苦枥木、野漆树(T.succedaneum)、白檀(Symplocospaniculata)、山樱花、野茉莉、白木乌桕、漆、野柿(Diospyroskaki)、枫香(Liquidambarformosana)、化香、山胡椒(Linderaglauca);草本层重要值>2.0的从大到小依次是海金沙、渐尖毛蕨(Cyclosorusacuminatus)、铁角蕨(Aspleniumtrichomanes)、救荒野豌豆(Viciasativa)、泽漆(Euphorbiahelioscopia)、蒲公英(Taraxacummongolicum)、铁苋菜(Acalyphaaustralis)、酸模(Rumexacetosa)。
2.2 山樱花种群年龄结构分析
云台山样地内山樱花种群为550株(图2),Ⅰ—Ⅷ龄级数量依次为147、141、114、78、40、18、8、4株。由图2可知,山樱花种群年龄结构呈不规律的“拓展型”,幼龄级个体数量多,老龄级个体则相反。

图2 山樱花龄级结构及数量分布图
山樱花种群个体数波动性较大,幼苗(第Ⅰ龄级)占总数的26.73%,幼龄级(Ⅱ—Ⅲ)个体占总数的46.18%,中龄级(Ⅳ—Ⅵ)个体占总数的24.91%,三者共占总个体数的97.82%;老龄级(Ⅵ—Ⅷ)个体比例仅为2.18%,其中最大胸径为30 cm。种群龄级结构表现为年轻个体在数量上占绝对优势,幼龄树和中龄树的数量非常丰富,而老龄级所占比重极少。说明云台山山樱花种群龄级结构为“J型”,属于增长型种群,自然更新能力强。
2.3 山樱花种群静态生命表及存活曲线
由山樱花种群静态生命表(表2)可知,从种群标准化存活个体数(lx) 上来看,随龄级的增加,山樱花种群lx逐渐减少,且个体减少的速度先增大再减小;第Ⅰ龄级山樱花生命期望(ex)最高,代表其生存能力最强;第Ⅶ龄级个体最弱,且平均生存能力一直随龄级增大而减弱,说明低龄级个体的生存能力较强,但随着龄级的增长而呈现不断减弱的趋势。不同阶段的生命期望变化可能和不同龄级对环境的适应性有关,也会直接影响群落内环境资源的分配,从而影响群落内各植物的种间关系。
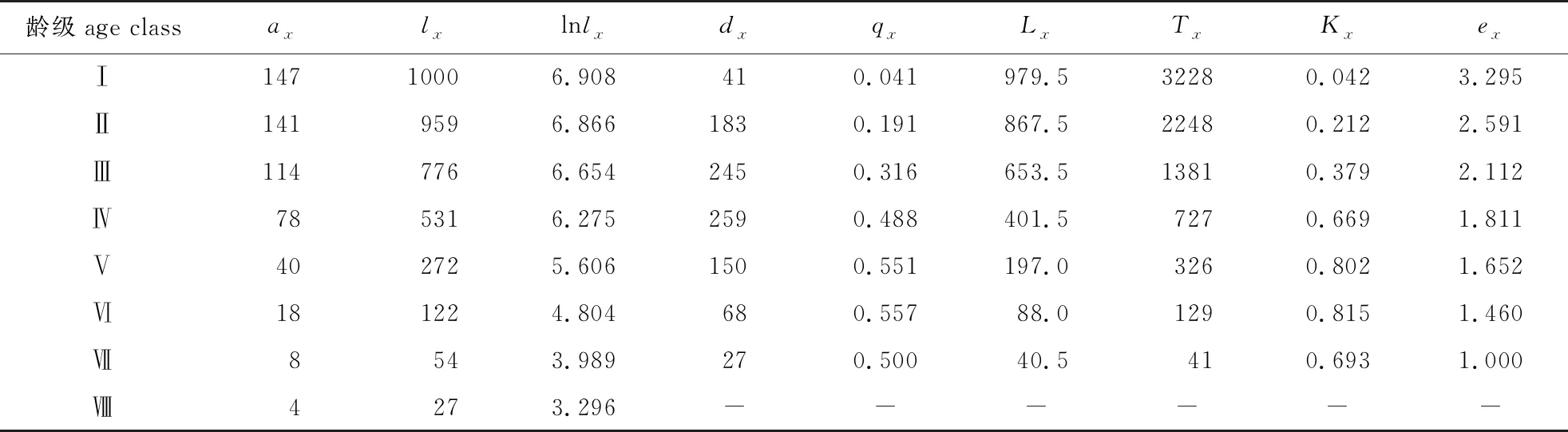
表2 山樱花种群静态生命表
存活曲线共分为3种类型,分别是Deevey-Ⅰ(凸型),表示接近生理寿命前只有少数个体死亡;DeeveyⅡ(对角线型)表示接近生理寿命前死亡率相等,DeeveyⅢ(凹型)型表示接近生理寿命前幼年期死亡率很高[40]。
由山樱花种群存活曲线(图3A)可见,其呈现出“凸-凹”的变化趋势,具体表现为Ⅰ—Ⅳ龄级可以认定为凸曲线即Deevey-Ⅰ型,代表该种群绝大多数山樱花个体都是能活到第Ⅳ龄级,Ⅰ—Ⅳ龄级死亡率较低;Ⅴ—Ⅷ龄级可以认定为凹曲线,代表该种群山樱花个体到达第Ⅴ龄级后,逐渐开始大面积凋亡,且山樱花种群存活曲线关于第Ⅳ龄级和第Ⅴ龄级的中点中心对称。由图3B可见,山樱花种群死亡率曲线qx和消失率曲线Kx走势雷同,随龄级增加先增大再减小,Ⅵ龄级时死亡率、消失率分别为0.557、0.815,均为最大值,死亡率和消失率曲线在Ⅰ—Ⅵ龄级呈上升态势,但经过Ⅵ龄级时间后反有大幅降低的趋势。在7个龄级中,只有Ⅰ、Ⅱ两个龄级的致死率(Kx)和死亡率(qx)均小于50%,即有50%以上的个体能存活至下一龄级。
存活曲线的下降趋势(图3A)与死亡率和消失率的上升趋势(图3B)可以互为佐证。由此可以推测,山樱花的第Ⅵ龄级在其生活史中是重要的转折期。幼龄级的山樱花种群存活率较高且变化趋势幅度较小,说明山樱花幼苗不需要大量的阳光,可以良好生长;随着龄级的增长,山樱花为强阳性树种,需要越来越多的生存资源和空间,但是在与群落建群种(栓皮栎和短柄枹栎)的竞争中处于弱势,因此其存活率随着龄级的增长呈现下降趋势;在第Ⅴ龄级后,山樱花个体可能突破林冠,通过环境的筛选可以获得足够的生存资源,不易死亡,个体达到比较稳定的状态。
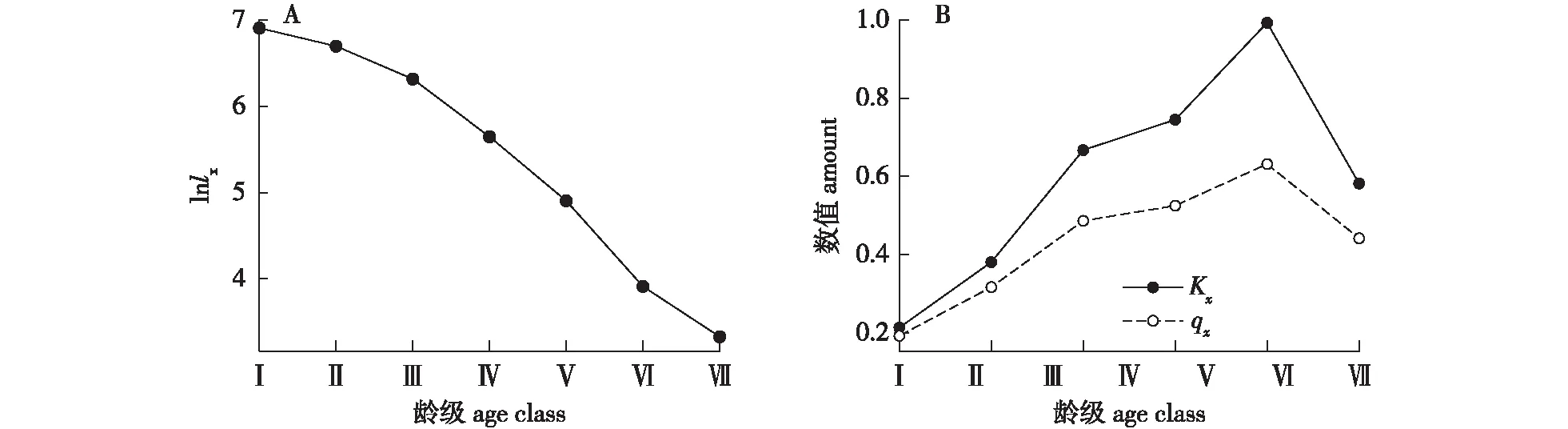
图3 山樱花种群存活曲线(ln lx)和死亡率(qx)、消失率(Kx)曲线
2.4 山樱花种群时间序列预测
在各龄级原始数量的基础上,利用依次移动平均法建立未来2、4、6、8个龄级时间后山樱花时间序列模型(表3)。经过2个龄级时间后,第Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ、Ⅶ、Ⅷ龄级涨幅分别为2.13%、12.28%、23.08%、47.50%、66.67%、62.50%、50.00%;经过4个龄级时间后,第Ⅳ、Ⅴ、Ⅵ、Ⅶ、Ⅷ龄级分别增长了53.85%、132.50%、250.00%、350.00%、350.00%;经过6个龄级时间后,第Ⅵ、Ⅶ、Ⅷ龄级分别增长了400.00%、737.50%、1000.00%;经过8个龄级时间后,第Ⅷ龄级个体数量已经在原始数量的基础上翻了15番。经历同等时间后,龄级区间内的个体数量增速越来越快,成活率也越来越高,即山樱花幼苗及低龄级植株过渡到成年的个体数也越来越多。综上所述,云台山山樱花种群保持着良好的增长状态,虽然根据时间序列预测,老龄级
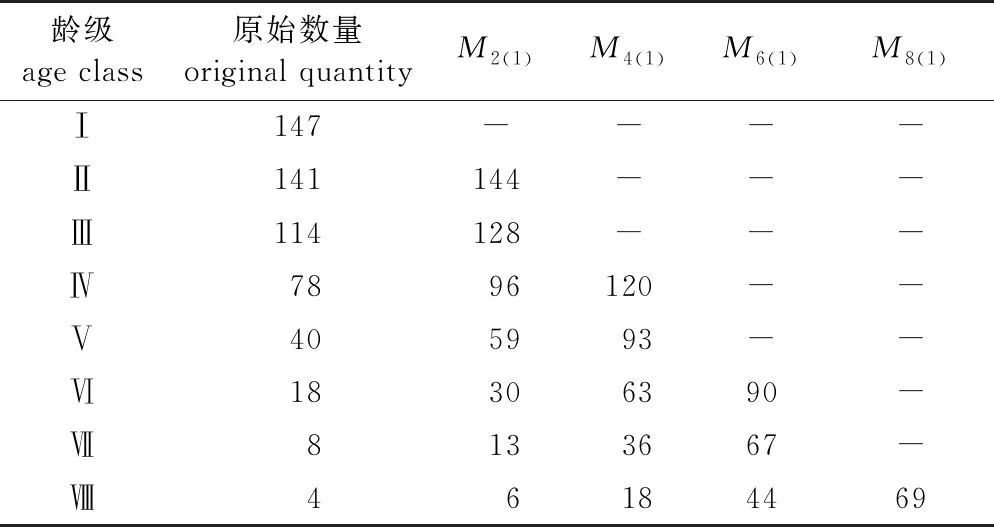
表3 山樱花种群龄级结构的时间预测
个体得到大量补充,但其幼龄级个体生命力旺盛,储备丰富;随发展趋势,低龄级、中龄级个体数量充足,完全可以满足后期龄级的更新,保持良好的种群结构。因此,研究样地植被演替过程中山樱花种群呈增长趋势,未来短期内仍是群落中的优势种群。
2.5 山樱花种群点格局分析
统计原始数据得:云台山山樱花种群共有山樱花550株,由山樱花种群不同发育阶段个体在样地中的空间分布(图4)可知,山樱花种群整体上与各年龄段个体并不均匀,呈现一定程度的集群分布,且各年龄段的植株密度存在较大差异,龄级较小的密度最大,密度随着龄级的增大而减小。集群分布的中心多是高龄级的山樱花个体,周围围绕着低龄级的山樱花个体,这应该与山樱花种子的扩散和繁殖有关。

以样地西南角为坐标系原点,x轴代表东西走向,y轴代表南北走向;数值代表样地与原点的距离。Take the southwest corner of sample plot as the origin of the coordinate system, the x-axis represents the east-west direction, the y-axis represents the north-south direction, the value represents the distance from the origin.
山樱花不同龄级的点格局分析结果见图5,从整体上看(图5A),山樱花种群整体的聚集强度随x轴坐标的增大而逐渐减小。山樱花个体在0~50 m坐标范围内的g(r)值大多略高或略低于置信区间,基本呈随机分布。
调查发现,山樱花种群整体的聚集强度随研究尺度的增大而逐渐减小。图5B中,第Ⅰ龄级在0~4 m坐标范围内表现为很突出的聚集分布,在4~50 m坐标范围内,g(r)值在25、32、41 m附近有小幅超出置信区间但绝大多数仍处于置信区间内,且曲线波动较小,属于随机分布。图5C中第Ⅱ龄级在5~50 m坐标范围内表现为随机分布,g(r)值超出置信区间的位点和幅度较图5B更少,曲线波动更平稳,但曲线走势趋于一致。图5D中第Ⅲ龄级在0~10 m坐标范围内偶有零星g(r)值超出置信区间,10~50 m坐标范围内分布较为随机。Ⅳ龄级(图5E)是分布最随机的龄级,除坐标6 m、18 m左右稍稍超出置信区间,其余部分都位于置信区间内,且波动范围较小,和存活曲线极值相呼应。第Ⅴ、Ⅵ龄级(图5F、5G)均在0~50 m的坐标范围内完全处于置信区间内。第Ⅶ、Ⅷ龄级由于龄级内个体数目较少,很多坐标范围内并没有该龄级个体的分布,所以会出现g(r)为零的水平区间,函数可信度下降,未能成图。

图5 山樱花种群不同发育阶段的点格局
对比图5B-5G可以发现,随着个体数量的减少,置信区间范围越来越广,曲线也越来越平稳,但是山樱花不同龄级的分布格局的变化仍然呈现一定的差异。全部个体的分布格局(图5A)与第Ⅰ、Ⅱ龄级的个体空间分布格局(图5B、5C)曲线相似;同时山樱花种群所有个体在坐标跨度较大时,呈随机分布。但随着坐标跨度变小,该种群逐渐体现出聚集分布的态势,这种分布格局很可能是由该区域的地形地貌、海拔、生存资源引起的,同时也可以认为该种群已经趋于稳定[41]。
3 讨 论
山樱花地理分布与水热环境因子相关研究,认为温度对山樱花分布的影响占绝对重要的作用,其中温度(年均温、极端低温、1月均温、极端高温)的重要性大于海拔、降水与经度[42]。通过对云台山山樱花群落结构、种群结构和点格局的研究分析,发现山樱花种群结构受其生物学特征和光照、水分等环境因素共同影响。本研究选取的样地是江苏云台山保护区内具有代表性的暖温带落叶阔叶林,样方内地形较复杂,群落原生性植被较多,有鲜明的由北亚热带向暖温带的过渡性质。“扩张型”种群年龄结构和Deevey-Ⅱ型存活曲线说明该山樱花种群属增长型,自然状况下进行自然更新没有任何压力。死亡率、消失率曲线和静态生命表的分析表明,山樱花种群具有幼龄级生命力旺盛,中龄级和老龄级逐渐衰退的特点。这符合山樱花种子萌发需冷凉湿润环境的特性,云台山紧邻东海,气候温暖湿润,故幼龄级个体存活率较高。随龄级的增长,山樱花死亡率逐渐升高,这是因为高大的阔叶树(短柄枹栎、栓皮栎)使得乔木层林相分为多层,且脱离幼龄级的个体对水肥、光照、空间等的要求日益增长,种间竞争大大限制了中龄级山樱花的生长更新。老龄级山樱花拥有更大的冠幅和更深的根系,对生存资源也有更强的竞争能力。时间序列预测结果说明山樱花种群经过2、4、6个龄级时间后仍可保持强劲的增长状态,说明山樱花目前没有进入衰老阶段且在未来一段时间内山樱花仍然会是该地区优势的种群,与年龄结构分析结果一致,可能和山樱花低龄级个体的大量贮备及其良好的更新能力有关。另外,该地区短柄枹栎成为绝对优势种的情况较为少见,可以尝试探究短柄枹栎与山樱花或该地之间的关联性。
本研究基于1 hm2的样地,利用点格局分析,客观地反映了江苏云台山山樱花种群的空间分布格局。不同龄级的山樱花分布格局不尽相同,幼、中龄级个体以聚集分布为主,多出现在老树周围,而老树个体呈随机分布;种群在20~30 m的空间尺度范围内呈一定程度的随机分布;随空间尺度的增加,分布趋于均匀分布这也符合自然群落种群分布的一般规律[43]。这与山樱花种子扩散机制息息相关:山樱花种子主要通过重力随地形扩散;山樱花果实为核果,可以食用,种子也可以通过鸟类取食传播。因此幼苗常聚集在母树周围产生“庇护效果”,有利于提高幼树成活率。随着年龄的增长,幼苗间的种内竞争愈发强烈,导致发育不良的个体逐渐凋亡,因此表现为随机分布或均匀分布。
综上所述,江苏省连云港市云台山位于亚热带—暖温带过渡区,山樱花种群分布区域生境良好,山樱花群落结构符合地带性植被特征,物种多样性上限高。云台山山樱花种群年龄结构呈增长型,在未来的群落演替进程中占据重要地位,随着云台山次生天然植被的持续保护与恢复,山樱花可以形成云台山地带性植被中重要的春花林相景观。结合野生山樱花的生长习性及其独特的观赏价值,建议完善保护区的管理制度,增强森林巡护,防止滥砍滥伐等行为;适度抚育,增加群落的物种多样性以保证群落的稳定性;合理引种驯化,丰富园艺种质资源。
参考文献(reference):
[1]王贤荣.中国樱花品种图志[M].北京:科学出版社,2014.WANG X R. An illustrated monograph of cherry cultivars in China Illustrated monograph of cherry cultivars in China[M]. Beijing:Science Press,2014.
[2]李蒙.山樱花高海拔居群生态学特征及组织培养[D].南京:南京林业大学,2013. LI M.Ecological characteristics and tissue culture of the high altitude population ofCerasusserrulata[D]. Nanjing: Nanjing Forestry University,2013.
[3]伊贤贵,陈洁,尤禄祥,等.山樱花群体遗传多样性的SSR分析[J].南京林业大学学报(自然科学版),2018,42(5):25-31.YI X G,CHEN J,YOU L X,et al.Genetic divetsity ofCerasusserrulatapopulations assessed by SSR markers[J]. J Nanjing For Univ (Nat Sci Ed),2018,42(5):25-31. DOI: 10.3969/j.issn.1000-2006.201702036.
[4]ZANG D K.Cerasuslaoshanensis(Rosaceae), a new species from Shandong, China[J]. Ann Bot Fenn, 2017, 54(1/2/3):135-137. DOI:10.5735/085.054.0321.
[5]王琳,朱淑霞,李蒙,等.樱花新品种‘惜春’[J].南京林业大学学报(自然科学版),2020,44(1):223-224.WANG L, ZHU S X, LI M, et al. A newCerasuscampanulatacultivar ‘Xichun’[J]. J Nanjing For Univ (Nat Sci Ed),2020,44(1):223-224. DOI: 10.3969/j.issn.1000-2006.201911009.
[6]朱淑霞,伊贤贵,王华辰,等.樱花新品种‘粉彩’[J].南京林业大学学报(自然科学版),2019,43(2):216-217.ZHU S X, YI X G, WANG H C, et al. A newCerasusjamasakuracultivar ‘Fencai’[J]. J Nanjing For Univ (Nat Sci Ed),2019,43(2):216-217. DOI: 10.3969/j.issn.1000-2006.201803055.
[7]KENT M. Vegetation description and data analysis: a practical approach[M]. 2nd ed. Weinheim: Wiley-Blackwell,2012.
[8]康佳鹏,马盈盈,马淑琴,等.荒漠绿洲过渡带柽柳种群结构与空间格局动态[J].生态学报,2019,39(1):265-276.KANG J P, MA Y Y, MA S Q, et al. Dynamic changes of spatial pattern and structure of theTamarixramosissimapopulation at the deser-toasis ecotone of the Tarim Basin[J]. Acta Ecol Sin,2019,39(1):265-276. DOI:10.5846/stxb201711262112.
[9]吴丹妮.油松主伐保留母树林下更新苗的格局分析[D].北京:北京林业大学,2019.WU D N. Spatial pattern analysis of regenerated seedlings ofPinustabuliformisunder main felling with reserved mother trees[D]. Beijing: Beijing Forestry University,2019.
[10]杨凤翔,王顺庆,徐海根,等.生存分析理论及其在研究生命表中的应用[J].生态学报,1991,11(2):153-158.YANG F X, WANG S Q, XU H G, et al. The theory of survival analysis and its application to life table[J].Acta Ecol Sin,1991,11(2):153-158.
[11]胡璇,徐瑞晶,苏琪,等.海南岛甘什岭特有植物无翼坡垒种群结构与动态[J].热带作物学报,2020(3):1-9.HU X, XU R J, SU Q, et al. Population structure and dynamics ofHopeareti-culata, a plant endemic to Ganshiling, Hainan Island[J]. Chin J Trop Crops, 2020(3):1-9.
[12]张晓晨,赵洋,熊中人,等.宝华山青冈种群年龄结构及点格局分析[J].南京林业大学学报(自然科学版),2018,42(6):77-83.ZHANG X C, ZHAO Y, XIONG Z R, et al. Population age structure and spatial point pattern ofCyclobalanopsisglaucain Baohua Mountain,Jiangsu Province, China[J]. J Nanjing For Univ (Nat Sci Ed), 2018, 42(6): 77-83. DOI: 10.3969/j.issn.1000-2006.201801001.
[13]徐建,韦新良,王敬,等.龙王山落叶阔叶林群落特征及物种多样性研究[J].西南林业大学学报,2014,34(3):19-26.XU J, WEI X L, WANG J, et al. Species diversity and the community characteristics of deciduous broad-leaved forest in Longwang Mountain[J]. J Southwest For Univ, 2014,34(3): 19-26. DOI: 10.3969/j.issn.2095-1914.2014.03.004.
[14]伊贤贵,丁晖,方炎明,等.基于固定样地的黄山不同海拔森林群落物种多样性分析[J].南京林业大学学报(自然科学版),2018,42(1):149-155.YI X G, DING H, FANG Y M, et al. Species diversity of forest communities at different altitudes based on fixed plot in Huangshan Mountains[J]. J Nanjing For Univ (Nat Sci Ed), 2018, 42(1): 149-155. DOI: 10.3969/j.issn.1000-2006.201610060.
[15]李晓玲,卢绪志,边震,等.山樱花组培快繁技术研究[J].林业科技,2014,39(5):1-3.LI X L, LU X Z, BIAN Z, et al. Study on rapid propagation ofCerasusserrulataby tissue culture[J]. For Sci Technol, 2014, 39(5):1-3. DOI: 10.3969/j.issn.1001-9499.2014.05.001.
[16]白欢欢,王雪峰,徐建国.天然黄山松群落主要树种营养生态位特征研究[J].南京林业大学学报(自然科学版),2018,42(2):81-88. BAI H H, WANG X F, XU J G. Nutrition ecological niche of dominant arbor species in naturalPinustaiwanensiscommunity[J]. J Nanjing For Univ (Nat Sci Ed), 2018, 42(2): 81-88. DOI: 10.3969/j.issn.1000-2006.201706043.
[17]NANTONGO J S, GWALI S. Long-term viability of populations ofPrunusafricana((Hook. f.) Kalm.) in Mabira forest: implications for in situ conservation[J]. African Journal of Ecology, 2018, 56(1): 136-139. DOI: 10.1111/aje.12423.
[18]NAVARRO-CERRILLO R M, CLEMENTE M, PADRN E, et al. Forest structure in harvested sites of Afromontane forest ofPrunusafricana[Hook.f.] Kalkm. in Bioko (Equatorial Guinea)[J]. African J Ecol, 2008, 46(4): 620-630. DOI: 10.1111/j.1365-2028.2008.00944.x.
[19]STEWART K. Effects of bark harvest and other human activity on populations of the African cherry (Prunusafricana) on Mount Oku, Cameroon[J]. Forest Ecology & Management, 2009, 258(7): 1121-1128. DOI: 10.1016/j.foreco.2009.05.039.
[20]Al-SHEHRI M A, El-SHEIKH M A, Al-FARHAN A H, et al. Ecology of endangeredPrunuskorshinskyiHand.-Mazz. in Jabal ai-lauz, Saudi Arabia: plant associations, size structure, and nutritional screening[J]. Saudi Journal of Biological Sciences, 2020, 27(1): 147-156. DOI: 10.1016/j.sjbs.2019.06.011.
[21]聂超仁,许小过,段庆明,等.两种方法提高山樱花种子发芽率试验[J].安徽农业科学,2014. 42(13):3835-3838.NIE C R, XU X G, DUAN Q M, et al. Two methods of improving germination rate ofCerasusserrualta(Lindl.) G. Don ex London[J]. J Anhui Agric Sci, 2014, 42(13): 3835-3838.
[22]李祯,伊贤贵,顾宇,等.山樱花基因组大小的测定[J].南京林业大学学报(自然科学版),2014(S1):17-19.LI Z, YI X G, GU Y, et al. The genome size determination ofCerasusserrulata[J]. J Nanjing For Univ (Nat Sci Ed), 2014(S1):17-19. DOI: 10.3969/j.issn.1000-2006.2014.S1.004.
[23]陈洁.山樱花居群遗传多样性的SSR分析[D].南京:南京林业大学,2016.CHEN J. Genetic diversity analysis ofCerasusserrulatapopulations based on SSR markers[D]. Nanjing: Nanjing Forestry University, 2016.
[24]伊贤贵.山樱花种群变异及亲缘地理学研究[D].南京:南京林业大学,2018.YI X G. The variation and phylogeography ofCerasusserrulataMill. populations[D]. Nanjing: Nanjing Forestry University, 2018.
[25]曹奇刚.连云港地区生态环境研究:以后云台山国家森林公园为例[J].水土保持研究,2007,14(6):100-102.CAO Q G. Research on the ecological environment of Lianyungang region: take Houyuntai Mountain National Forest Park as an example[J]. Res Soil Water Conserv, 2007, 14(6): 100-102.
[26]张勇,李土生,潘江灵,等.连云港市云台山宜林荒山立地质量分类及评价[J].水土保持通报,2014,34(3):171-177.ZHANG Y, LI T S, PAN J L, et al. Site quality classification and assessment in barren hills for suitable afforestation in Yuntai Mountains of Lianyungang City[J].Bull Soil Water Conserv,2014,34(3):171-177.
[27]赵增玉,陈火根,潘懋,等.基于GOCAD的宁芜盆地云台山地区三维地质建模[J].地质学刊,2014,38(4):652-656.ZHAO Z Y, CHEN H G, PAN M, et al. 3D geological modeling of Yuntaishan area in Ningwu Basin based on GOCAD software[J].J Geol,2014, 38(4):652-656. DOI: 10.3969/j.issn.1674-3636.2014.04.652.
[28]钟华邦.地质素描:江苏云台山地貌[J].地质学刊,2012,36(1):47.ZHONG H B. Geological sketch: the Yuntai Mountain landform in Jiangsu[J]. J Geol, 2012, 36(1):47.
[29]孟艳,赖雨,杨文嘉,等.江苏云台山三桠乌药群落研究[J].生物学杂志,2016,33(6):68-72.MENG Y, LAI Y, YANG W J, et al. Study on community ofLinderaobtusilobain Yuntai Mountain of Jiangsu Province[J].J Biol,2016,33(6):68-72. DOI:10.3969/j.issn.2095-1736.2016.06.068.
[30]张佳平.云台山野生草本植物资源的园林开发利用评价[J].南京林业大学学报(自然科学版),2013,37(1):37-43.ZHANG J P. Comprehensive evaluation of landscape exploitation and application of wild herbaceous plant resources of Yuntai Mountain[J]. J Nanjing For Univ (Nat Sci Ed), 2013, 37(1): 37-43.
[31]李红芳,丁平,巢建国.江苏省连云港市云台山药用植物资源概况[J].北方药学,2012,9(5):114.LI H F, DING P, CHAO J G. Investigation on resources of medicinal plants in Yuntai Mountain of Lianyungang, Jiangsu Province[J]. North Pharm, 2012, 9(5):114.
[32]张佳平,丁彦芬.连云港云台山野生草本植物资源调查、应用及保护研究[J].草业学报,2012,21(4):215-223.ZHANG J P, DING Y F. A study on resource investigation, application and protection of wild herbaceous plants of the Yuntai Mountain in Lianyungang[J]. Acta Prataculturae Sin., 2012, 21(4):215-223.
[33]周戎.江苏省云台山地区楸树生长规律与分布研究[D].南京:南京林业大学,2009.ZHOU R.Study on growth regulartity and distribution ofManchuriancatalpain Yuntai Mountainous areas of Jiangsu Province[D]. Nanjing: Nanjing Forestry University, 2009.
[34]曹秀芹,宋维春.江苏省云台山区野生果树种质资源初报[J].中国果树,2004(1):10-12.CAO X Q, SONG W C. Initial report on germ plasm resources of wild fruit trees at the Mount Yuntai in Jiangsu Province[J]. China Fruits, 2004(1): 10-12. DOI: 10.3969/j.issn.1000-8047.2004.01.006.
[35]谢春平,吴昌魁,付桂,等.五指山地区海南苏铁种群结构特征与动态[J].中南林业科技大学学报,2019,39(1):77-85.XIE C P, WU C K, FU G, et al. Population structure characteristics and dynamics ofCycashainanensisin Wuzhi Mountains, Hainan Province[J]. J Central South For Univ, 2019, 39(1):77-85.
[36]谢春平,邱靖,伊贤贵.南京城市近郊构树种群结构分析[J].云南农业大学学报(自然科学),2018,33(4):696-704.XIE C P, QIU J, YI X G. Analysis of the population structure ofBroussonetiapapyriferain suburban of Nanjing[J]. J Yunnan Agric Univ, 2018, 33(4):696-704. DOI: 10.12101/j.issn.1004-390X(n).201705037.
[37]郭有燕,张文辉,何景峰,等.黄土高原丘陵区文冠果种群特征研究[J].西北农林科技大学学报(自然科学版),2011,39(1):61-68.GUO Y Y, ZHANG W H, HE J F, et al. Population characteristic ofXanthocerassorbifoliain hill area of Loess Plateau[J]. J Northwest Sci-Tech Univ Agr, 2011, 39(1): 61-68.
[38]吴秀萍,许晓岗,王璐,等.江西武夷山南方铁杉种群结构与点格局分析[J].北京林业大学学报,2019,41(4):60-68.WU X P, XU X G, WANG L, et al. Population structure and spatial distribution point patterns ofTsugachinensisvar.tchekiangensisin Wuyishan Mountain,Jiangxi of eastern China[J]. J Beijing For Univ, 2019, 41(4): 60-68. DOI:10.13332/j.1000-1522.20180309.
[39]沈志强,卢杰,华敏,等.西藏色季拉山高山松种群点格局分析[J].西北农林科技大学学报(自然科学版),2016,44(5):73-81.SHEN Z Q, LU J, HUA M, et al. Spatial point pattern ofPinusdensataforestsof Sejila Mountains in Tibet[J]. J Northwest Sci-Tech Univ Agr, 2016, 44(5): 73-81. DOI: 10.13207/j.cnki.jnwafu.2016.05.010.
[40]DEEVEY E S Jr. Life tables for natural populations of animals[J]. Q Rev Biol, 1947, 22(4): 283-314. DOI:10.1086/395888.
[41]张甍.太岳山针阔混交林不同海拔空间格局研究[D].临汾:山西师范大学,2017.ZHANG M. A study on the spatial pattern of coniferous and broad-leaved mixed forest at different altitude in Taiyue Mountain[D]. Linfen: Shanxi Normal University, 2017.
[42]李蒙,伊贤贵,王华辰,等.山樱花地理分布与水热环境因子的关系[J].南京林业大学学报(自然科学版),2014,38(S1):74-80.LI M, YI X G, WANG H C, et al. Studies on the relationship betweenCerasusserrulatadistribution region and the environmental factors[J]. J Nanjing For Univ (Nat Sci Ed), 2014, 38(S1):74-80.
[43]宗国,白雪娇,张淑媛,等.辽东山区次生林乔木幼苗分布格局与种间空间关联性[J].应用生态学报,2018, 29(1):18-24. ZONG G, BAI X J, ZHANG S Y, et al. Distribution pattern and interspecific spatial correlation of tree seedlings in secondary forests in eastern Liaoning[J]. Chin J Appl Ecol, 2018, 29(1):18-24. DOI: 10.13287/j.1001-9332.201801.002.
