干旱胁迫对毛豹皮樟扦插苗光合作用与叶片组织结构的影响
2021-07-27曾维军屈坤杰席培宇李蕾佳王济红
曾维军,屈坤杰,万 诚,李 鹏,席培宇,李蕾佳,王济红
(1.贵州省生物研究所,贵州 贵阳 550009;2.上海市岩土地质研究院有限公司,上海 200072)
【研究意义】毛豹皮樟(Litsea coreanaLevl.var.lanuginosa)是樟科木姜子属植物朝鲜木姜子(L.coreana)的变种,零星分布于贵州、四川、重庆、安徽、广东和云南等亚热带地区。嫩叶是西南地区传统茶饮老鹰茶的主要原料,具有消暑解渴、消食祛胀、降血糖血脂等功效[1-2]。在实际应用中,作为老鹰茶原料的毛豹皮樟种植于坡地,洪涝较少,但地理、气候等环境因素造成的干旱缺水情况较多,制约毛豹皮樟的生长发育,间接影响老鹰茶的品质,所以在干旱胁迫下对毛豹皮樟进行植物逆境生理研究尤为重要。【前人研究进展】近5 年,关于毛豹皮樟的研究主要集中在形态特征及地理分布[3]、遗传选育[4]、开发及利用[5]、化学成分及药理作用、生理特征、分子生物学等方面。例如,唐茜等[6-7]研究表明,矿物质营养主要是钾、镁、磷、钙、钠、铁和锰,均为人体必需营养,其中钾参与人体蛋白质、氨基酸和碳水化合物的代谢,对心脏和心血管具有保护作用,能降低血压[8]。范川等研究了毛豹皮樟苗木对低温胁迫的生理响应,发现毛豹皮樟的过氧化氢酶(CAT)、过氧化物酶(POD)活性及丙二醛(MDA)含量在处理温度及品种间具有显著差异,均可作为毛豹皮樟抗寒性评价指标,同时还发现超氧化物歧化酶(SOD)与毛豹皮樟抗涝性有极强的相关性[5];经方差分析发现,毛豹皮樟的CAT、POD 活性及SS、MDA 含量在处理温度及品种间都有显著差异,均可作为毛豹皮樟抗寒性评价指标[9-10]。巩艳红等[11]采用改良SDS 法从毛豹皮樟的老、幼叶片中提取到高质量DNA,可以用于RAPD 分析,从4组随机引物中筛选出16 个显示毛豹皮樟遗传差异的多态性引物,该方法适用于毛豹皮樟的种质资源鉴定分析。【本研究切入点】关于逆境生理下叶片光合能力及组织形态差异的相关研究少见报道,不能进一步阐明毛豹皮樟生理生态的相关规律。【拟解决的关键问题】本研究以用于生产种植的毛豹皮樟扦插苗为研究对象,探究不同程度干旱胁迫对毛豹皮樟扦插苗叶片组织结构及光合作用的影响,探究毛豹皮樟扦插苗逆境生理特性,为其高效栽培管理技术提供理论支撑。
1 材料与方法
1.1 试验材料
供试材料为毛豹皮樟3年生扦插苗,平均株高25(±2)cm、基径粗3(±0.5)mm,由贵州省生物研究所培育。
1.2 试验地点
试验地点位于贵州省湄潭县兴隆镇丁家沟村(107°48′E、27°67′N)。试验场地为覆膜简易大棚,用透光率50%的遮阳网做外遮阳,温度为自然变温,简易塑料大棚上覆盖透光率为50%的遮阳网。试验地属亚热带湿润季风气候,夏季温暖湿润,冬季干燥寒冷,年均气温14.9℃,最冷月(1 月)平均气温3.8℃,最热月(7 月)平均气温25.1℃,无霜期平均284 d,年日照时数 1 163 d。试验于2018 年6 月进行,该月平均气温25.6℃。
1.3 试验方法
用口径15 cm×高20 cm 塑料盆栽植,栽培基质为腐质土、蛭石(体积比1 ∶1)的混合基质;每盆装入基质1.0 kg,定植1 株。确定混合基质田间最大持水量,设置4 种不同程度的干旱胁迫处理:CK(土壤体积持水量为39.6%)、轻度(土壤体积持水量为29.2%)、中度(土壤体积持水量为19.5%)、重度(土壤体积持水量为9.7%),采取称重法定期补水。每个处理30 株苗,3 次重复。2018年1月2日定植,2月1日开始处理(定植30 d),5 月5 日后所有处理统一水管理(5 d 浇一次透水),6 月5~10 日开始取样和测量相关指标。
1.4 测定方法
1.4.1 光合日变化测定 6 月5 日,晴天,8:00~ 18:00,每隔1 h 分别选定相同干旱处理下3 株毛豹皮樟扦插苗顶枝的一片成熟叶,使用Li6400光合测定仪(LI-COR,美国)测量其净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr),同时记录光合有效辐射(PAR)、空气中CO2浓度(Ca)、空气温度(T)及相对湿度(RH)。
1.4.2 光合气体交换参数测定 6 月10 日,晴天,9:30~11:30,使用带红蓝光源的LI6400 光合测定仪(LI-COR,美国),以对照顶枝叶片光响应曲线光饱和点(1 192 μmol/m2·s)为参考,设定光强1 100 μmol/m2·s,选定相同干旱处理下3 株毛豹皮樟扦插苗顶枝的一片成熟叶,测量其Pn、Gs、Ci、Tr。
1.4.3 叶片气孔限制值水分利用效率、计算 气孔限制值Ls=1-Ci/Ca。其中,Ca 为空气中CO2浓度。水分利用效率WUE=Pn/Tr。
1.4.4 叶绿素总量测定 不同干旱胁迫程度下各选取3 株毛豹皮樟扦插苗顶枝一片成熟叶,使用UV-1800SPC 双光束紫外分光光度计(Macy,中国美析仪器有限公司),采用丙酮乙醇混合液法测定叶绿素总量(Tc)[12]。
1.4.5 叶片组织结构测量 不同干旱胁迫程度下各选取1 株毛豹皮樟扦插苗顶枝一片成熟叶进行制片,使用Primovert 显微镜(ZEISS,德国卡尔·蔡司股份公司)在每张切片上随机挑选5 个200 倍视野进行观察和拍照。应用Image-Pro Plus 6.0 软件以200 倍标尺为标准,选取10 处测量每张片子上表皮厚度、下表皮厚度,测量栅栏组织和海绵组织的厚度、密度(每个视野中栅栏组织和海绵组织的个数)。
1.5 数据处理
使用Graph Pad Prism9.0,对不同干旱胁迫程度下毛豹皮樟扦插苗叶片气体交换参数、水分利用效率、气孔限制值、叶绿素总量进行方差分析,采用S-N-K 法进行多重比较,实现数据实时可视化。使用SPSS24.0 软件,采用皮尔逊相关性分析法对光合和环境日变化参数进行相关分析。
2 结果与分析
2.1 干旱胁迫对毛豹皮樟扦插苗叶片净光合速率日变化的影响
由表1可知,轻度干旱胁迫下毛豹皮樟扦插苗叶片没有明显的光合午休现象,曲线呈单峰型,日均净光合速率为3.53 μmol/m2·s,15:00 时净光合速率达峰值6.22 μmol/m2·s;CK、中度、重度干旱胁迫的日均净光合速率分别为3.69、3.33、1.97 μmol/m2·s,均存在明显光合午休现象,其中CK曲线呈单峰型,光合午休时间最短、为1 h;中度、重度干旱胁迫的光合日变化为双峰曲线;重度干旱下净光合速率日变异系数最大、为68.23%,CK最小、为28.65%。
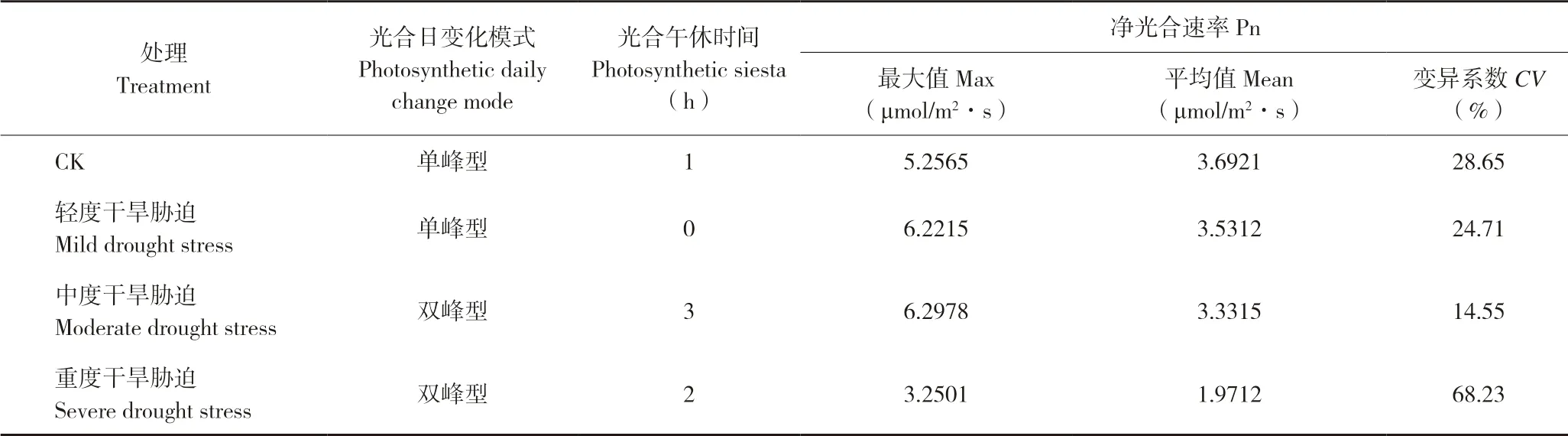
表1 不同干旱胁迫程度下豹皮樟扦插苗叶片净光合速率日变化Table 1 Diurnal changes of net photosynthetic rate of Litsea coreana Levl.var.lanuginosa cutting seedlings under different drought stresses
2.2 不同干旱胁迫程度下毛豹皮樟扦插苗叶片光合和环境参数日变化相关性分析
如表2 所示,在毛豹皮樟扦插苗叶片光合和环境参数的16 组对应组合中,CK 有10 组对应组合显著相关,轻度干旱胁迫有15 组,中度干旱胁迫15 组,重度干旱胁迫11 组;CK 的Ci 与RH 的正相关性最高、相关系数为0.689,Tr 与RH 的负相关性最高、相关系数为-0.593;轻度干旱胁迫下,Ci 与RH 的正相关性最高、相关系数为0.870,Pn 与Ca 的负相关性最高、相关系数为-0.920;中度干旱胁迫下,Gs 与RH 的正相关性最高、相关系数为0.749,Gs 与T 的负相关性最高、相关系数为-0.723;重度干旱胁迫下,Gs与RH 的正相关性最高、相关系数为0.913,Gs与T 的负相关性最高、相关系数为-0.869。

表2 不同干旱胁迫程度下毛豹皮樟扦插苗叶片光合和环境参数日变化相关性Table 2 Correlation of diurnal changes of leaf photosynthetic and environmental parameters of Litsea coreana Levl.var.lanuginosa cutting seedlings under different drought stresses
2.3 干旱胁迫对毛豹皮樟扦插苗叶片气体交换参数的影响
如图1 所示,随着干旱胁迫程度增加,毛豹皮樟扦插苗叶片Pn 均值显著降低,数据离散程度增加,中度干旱胁迫的Pn 均为接近于0 的负值;CK 的Gs 显著高于其他3 个干旱处理,中度、重度干旱胁迫下的Gs 差异不显著;两者Ci 均值较高,CK 的Ci 均值较低,CK、重度干旱胁迫的Ci值聚集程度较高;处理间Tr 相比较,CK 与轻度干旱胁迫差异不显著,但显著高于中度、重度干旱胁迫。

图1 干旱胁迫对毛豹皮樟扦插苗叶片气体交换参数的影响Fig.1 Effects of drought stresses on gas exchange parameters of Litsea coreana Levl.var.lanuginosa cutting seedlings
2.4 干旱胁迫对毛豹皮樟扦插苗叶片水分利用效率、气孔限制值和叶绿素总量的影响
由图2 所示,中度、重度干旱胁迫下毛豹皮樟扦插苗叶片的Ls、WUE 均为负值,其中重度干旱胁迫下的Ls 值最低,中度干旱胁迫下的WUE 值最低,轻度干旱胁迫下的Ls 显著高于CK,CK 的WUE 显著高于轻度干旱胁迫,是轻度干旱胁迫下的近两倍。CK 的Tc 为1.61 mg/L,极显著高于轻度、重度干旱胁迫,显著高于中度干旱胁迫,其中轻度和重度干旱胁迫下的WUE 差异不显著,但均显著低于中度干旱胁迫。
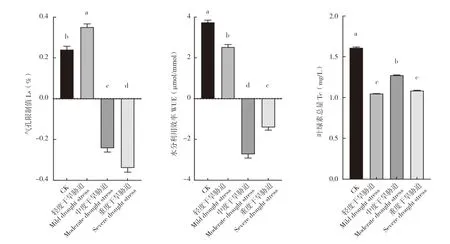
图2 干旱胁迫对毛豹皮樟扦插苗叶片水分利用效率、气孔限制值和叶绿素总量的影响Fig.2 Effect of drought stresses on WUE,Ls and Tc of Litsea coreana Levl.var.lanuginosa cutting seedlings
2.5 干旱胁迫对毛豹皮樟扦插苗叶片横切组织结构的影响
由图3、图4 可知,3 个干旱处理的毛豹皮樟扦插苗叶片上表皮厚度均显著小于CK;随着干旱胁迫程度增加,叶片下表皮厚度先减后增;CK 的栅栏组织厚度最高、轻度最低;海绵组织厚度均值的变化随干旱胁迫的加强而先增后减,重度干旱胁迫的海绵组织厚度均值最高、为0.097 mm;干旱处理下的叶片栅栏组织密度与CK 相比均不显著;轻度干旱胁迫的叶片海绵组织密度为179 个,显著高于CK。
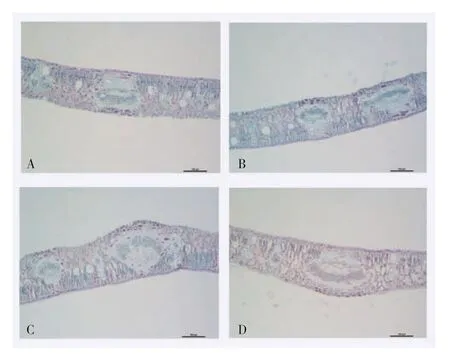
图3 干旱胁迫对毛豹皮樟扦插苗叶片横切组织显微结构的影响Fig.3 Effects of drought stresses on the microstructure of leaf transverse tissue structures of Litsea coreana Levl.var.lanuginosa

图4 干旱胁迫对毛豹皮樟扦插苗叶片横切组织结构的影响Fig.4 Effect of drought stresses on leaf transverse tissue structure of Litsea coreana Levl.var.lanuginosa cutting seedlings
3 讨论
“光合午睡”过长不利于植物光合作用积累有机物。对照、中度、重度干旱胁迫下毛豹皮樟均有明显“光合午睡”现象,中度时间最长,轻度干旱胁迫无此现象,说明轻度干旱胁迫能打破毛豹皮樟“光合午睡”,但继续干旱胁迫会加重该现象,不利于光合作用。光合能力是植物进行新陈代谢的基础,也是影响作物品质和产量的重要条件[13]。净光合速率反映植物积累有机物快慢,是衡量光合能力的重要参数。净光合速率日变化模式有双峰型、单峰型、平坦型和不规则型等多种类型[14],不同干旱胁迫程度下毛豹皮樟扦插苗叶片净光合速率日变化类型主要是双峰型和单峰型。重度干旱胁迫下的净光合速率日变异系数最大,高于对照2 倍,同时均值和最大值也最低,说明自然环境中遇到严重干旱,毛豹皮樟利用光能合成有机物的水平较低且不稳定。随着干旱程度增加,光合参数与环境因子的相关性先增后降,且相关程度也在发生变化,其中关联性较高的环境因子为空气相对湿度和温度。
干旱胁迫往往显著降低植物的光合作用[15],与本试验中“毛豹皮樟扦插苗叶片净光合速率均值伴随干旱胁迫程度增加而显著降低”的结论相似。对照的气孔导度高,胞间CO2浓度低,干旱胁迫下则相反,说明干旱胁迫显著影响毛豹皮樟扦插苗叶片的气孔开张程度,让气孔变小甚至关闭,导致胞间CO2浓度升高。中度和重度干旱下,气孔限制值和水分利用效率呈负值,说明中度干旱开始毛豹皮樟扦插苗已经缺水、叶片气孔不能正常工作,同时影响叶片叶绿素合成、叶绿素总量降低,但在中度干旱下,叶绿素总量有小幅增加,可能是毛豹皮樟响应干旱胁迫的应激反应。
在自然生长条件下,核酮糖-1,5-二磷酸梭化酶/加氧酶(Rubisco)的活化、核酮糖-1,5-二磷酸(RuBP)的再生是完整叶片以最佳状态进行光合作用的两个重要代谢步骤,主要发生在叶片的栅栏组织和海绵组织细胞内[16-17]。细胞内真正的CO2供应速率,则受叶片表皮部分气孔保卫细胞的控制[18-20]。3 个干旱处理的叶片上表皮厚度均显著小于对照,说明干旱胁迫通过改变表皮气孔保卫细胞的状态来影响CO2供应速率,从而影响毛豹皮樟扦插苗的光合速率;CK 叶片栅栏组织厚度最高,重度干旱胁迫最低,中度干旱胁迫的叶片海绵组织厚度均值最高,轻度干旱胁迫的叶片海绵组织密度最高与中度、重度干旱胁迫差异不显著,说明毛豹皮樟扦插苗叶片在干旱胁迫下响应光合作用而特化出来的细胞减少、气体交换增加,栅栏组织是豹皮樟扦插苗叶片传递环境改变信号的传递层,而海绵组织则是应对这种环境改变的反应层。
4 结论
干旱胁迫影响毛豹皮樟扦插苗叶片“光合午睡”时长、净光合速率日变化及气体交换参数与环境因子的相关性,重度干旱加重“光合午睡”,降低光合能力,维持叶片正常光合生理需增加空气水分、降低温度;干旱胁迫影响毛豹皮樟扦插苗叶片气孔开张状态和叶绿素总量,中度干旱下能增加叶片叶绿素总量来响应干旱胁迫,重度干旱严重影响其水分利用和气体交换;毛豹皮樟扦插苗通过降低叶片上表皮厚度、增加栅栏组织厚度、降低海绵组织厚度来响应干旱胁迫。
