柳树耐盐机制及耐盐基因研究进展
2021-07-26江钰娜朱星兆刘昱刘国元张健
江钰娜 朱星兆 刘昱 刘国元 张健


摘要:盐分胁迫是影响植物生长发育的主要非生物胁迫之一,主要引起植物对土壤水分、养料吸收障碍,对植物造成渗透胁迫、离子毒害、光合作用减弱等不利影响。柳树可适应各种不良生境,作为抗逆性强的园林树种,对美化环境和改善土壤均有积极作用。从柳树对盐胁迫的生理生化反应及耐盐响应机制2个方面展开讨论,总结了耐盐品种的选育以及柳树耐盐候选基因的研究进展。在传统育种的基础上,结合现代分子技术培育耐盐柳树品种,为沿海耐盐林木和沿海滩涂开发提供有利资源。
关键词:柳树;耐盐机制;盐胁迫;耐盐品种;耐盐基因
中图分类号: S792.120.1 文献标志码: A
文章编号:1002-1302(2021)11-0028-07
收稿日期:2020-09-25
基因项目:国家自然科学基金(编号:31971681);江苏省林业科技创新与推广项目(编号:LYKJ[2018]36);南通大学引进人才科研启动项目(编号:18R08)。
作者简介:江钰娜(1996—),女,硕士研究生,主要从事园林植物抗逆机理相关研究。E-mail:18252098108@163.com。
植物生长过程会受到众多非生物胁迫,盐胁迫为其中之一,它限制植物生长,给生产和环境带来巨大负担[1]。我国盐碱地占比很大,主要分布在北方以及沿海地区。在沿海地区,滨海盐碱地是土地的主要组成部分,除了少数由海平面自然淤积形成的外,大部分滨海盐碱地是由人工围垦成,目前这类土壤面积还在持续增加。随着人口日益增加,耕地资源越来越紧缺,自然形成及人工围垦形成的滨海盐碱土都是重要的后备土地资源,具有重要的经济和生态价值[2]。造林是解决土壤盐渍化的有效生物方法,在盐碱地上种植耐盐林木,可持续改良环境。研究表明,种植耐盐树木不仅可增加地面绿化覆盖,而且可以缓解土壤的含盐量,促进脱盐率[3]。柳树是一种杨柳科(Salicacea)柳属(Salix)树种,属于落叶乔灌木,全球共有520多种;在我国分布广泛,因其形态优美,常作为园林观赏植物,具有生态多样性、遗传多样性的特点,尤其因它具有广泛的适应性,在许多不良生境中都能生长,被认为是“社会与环境的资源”[4-6]。柳树是盐碱地的重要种植树种,其耐盐性备受关注[7-11]。柳树在盐碱地不仅可以生存,还能改善土壤有机质含量,进而改善环境。研究表明,盐胁迫下柳树通过根系吸收并向上运输离子,可降低土壤中的钠离子含量,可见柳树对改良盐碱土壤有着一定作用[12];但是土壤盐碱化程度加剧极大地限制了柳树的种植。盐胁迫会引起柳树根茎叶受损[13]、苗高降低[14]、基径减小[15]、成活率下降[16],引起柳树水分不足、渗透胁迫、离子毒性等损伤,紧接着会逐渐降低柳树的光合作用和生长代谢强度。研究表明,盐胁迫引起柳树叶片叶绿素含量降低,根系活力减弱,超氧化物歧化酶(SOD)含量逐渐降低,丙二醛含量及相对电导率增加[17]。像大多数植物一样,柳树可以通过维持细胞离子稳态积累渗透调节物质,并通过启动有效的信号转导网络活化活性氧清除剂,以适应盐胁迫。本文综述了柳树对盐胁迫的生理和分子反应,包括盐胁迫对柳树生长的影响、柳树耐盐性的主要机制以及针对柳树耐盐性进行遗传改良的目标基因。
1 盐胁迫对柳树生长的影响
1.1 盐胁迫导致柳树水分缺失
盐胁迫破坏了植物的水势稳态和离子分布,这种状态的失衡发生在植物的细胞水平和个体水平上[13,18]。盐分过多会阻碍植物根系吸收水分,导致液泡中的水分流出,进而降低胞内渗透压导致渗透机制失调。植物无法从外界吸收水和养料,影响其生长代谢和光合作用,严重时会导致植物死亡[19]。在盐胁迫下,当植物暴露的根数量较小时,可观察到盐诱导下根细胞程序性死亡,但是枝条受到Na+毒害的情况往往要在数周以后才会暴露出来[20]。隋德宗等采用水培方法观察盐处理下5个柳树无性系幼苗根系生长,结果发现一定程度的低盐处理可以刺激根系生长,而持续的高盐胁迫下根外部水势显著下降,导致根系吸水困难,抑制了柳树生长[21]。刘铎等发现,随着盐浓度的增加,柳树的叶片鲜质量、叶片相对含水量都会降低,同时株高净生长量、生物量减少[22]。聂莉莉等通过试验发现,土壤持水量和地下水含盐量共同影响着柳树的生长年限[23]。
1.2 鹽胁迫引起柳树离子毒害
Na+不是植物生长必需的营养元素,Na+积累过多会影响植物生长所需的其他离子的吸收利用,比如K+。Shelden等研究了离子稳态对植物盐分的耐受性,得出植物通过Na+和K+转运机制适应土壤盐分;在根尖元素含量观察中,Na+和K+含量存在显著正相关关系;减轻Na+毒性的一种机制是将Na+转移到植物液泡中,置换出K+来维持细胞质中较高的K+含量/Na+含量,所以K+含量/Na+含量是植物盐胁迫程度的衡量指标[24]。通过减少根的分生组织和细胞分裂区域中Na+的吸收量或有效排出Na+可保护根尖细胞分裂[25-26]。王宁等研究了一种有较强区隔盐离子能力的柳树无性系SH31,发现短期内SH31中Na+含量显著上升,在根、茎中的K+含量下降,叶中的K+含量升高,SH31可维持较高的K+/Na+,运输器官茎中大量盐离子的存在有利于根尖细胞分裂,从而实现特定组织内的离子平衡[27]。Mirck等在温室观察SX64、9882-34、9870-40 这3种柳树的生长情况,发现所有品种的根吸收了大部分的Na+,而叶子接收了大部分的Cl-,芽中的Na+/K+、Na+/Ca+保持平衡,说明这3个柳树品种有较强的离子区隔化能力,以适应盐环境生长[28]。在盐处理下,有些敏盐品种的柳树较早地出现了生长衰败现象,可能是离子区隔能力的差异导致,可见不同柳树品种的离子区隔能力是不同的。
1.3 盐胁迫引发柳树氧化危害
盐胁迫不仅会引起柳树水分缺失和受离子毒害,还会导致其细胞内活性氧物质积累,形成氧化胁迫。活性氧(ROS)是一类由O2转化而来的自由基或具有高反应活性的离子或分子,低浓度的活性氧不会对植物产生危害,而能在植物细胞信号转导中作为第二信使来介导应答反应。一旦ROS积累过量,则会由利转害,造成氧化损伤甚至细胞死亡[29]。盐碱、高温等非生物胁迫会造成活性氧在细胞内大量产生,引起质膜损伤,并最终分解成丙二醛(MDA)等化合物。Janero等研究提出,丙二醛含量可作为脂质过氧化和过氧化组织损伤的诊断指标。ROS对膜脂和细胞内酶活性的伤害程度及MDA含量,常作为评判植物耐盐性的重要指标之一[30]。李小艳等研究了7种不同品种柳树的盐胁迫抗性表现,发现在200 mmol/L盐浓度下,筐柳和北沙柳抗性较弱,它们的MDA含量不断上升,其余品种MDA含量小幅增加[12]。
1.4 盐胁迫引起的柳树光合减弱
树木生长发育所需的能量和物质来源于光合作用,光合作用是植物盐胁迫最敏感的生理过程,持续的盐胁迫可能会降低光合作用并导致树木死亡,光合作用降低的原因包括叶绿素浓度降低、营养成分发生变化、气孔关闭等[31-32]。乔志攀对6种耐盐灌木柳进行盐处理时发现,在6‰、8‰、10‰、12‰ NaCl质量分数溶液处理下,柳树无性系的叶绿素总含量呈下降趋势[33]。而李小艳在研究7种柳树的致死盐浓度时发现,在100 mmol/L NaCl处理下杞柳、筐柳和竹柳叶片的光合参数与对照相比没有明显差异,但在200 mmol/L NaCl处理下,叶片光合参数均低于对照,表明在达到柳树盐处理极限浓度前,光合作用的指标也有所变化[12]。周鹏等以2个不同的耐盐性灌木柳无性系为试材,测定盐胁迫下柳树PSⅡ荧光特性参数,发现耐盐型JW2345的参数变化均明显低于盐敏感型JW2367[34]。季琳琳等比较了盐处理下竹柳3号、苏柳172的光合特征变化,试验表明两者的气孔导度随着盐浓度增加而降低[35]。叶绿素、叶片光合参数以及气孔导度等反映光合作用程度的指标显示,盐浓度会降低柳树的光合作用并且耐盐型的柳树光合指标降低得比盐敏感型程度轻。
2 柳树耐盐响应机制
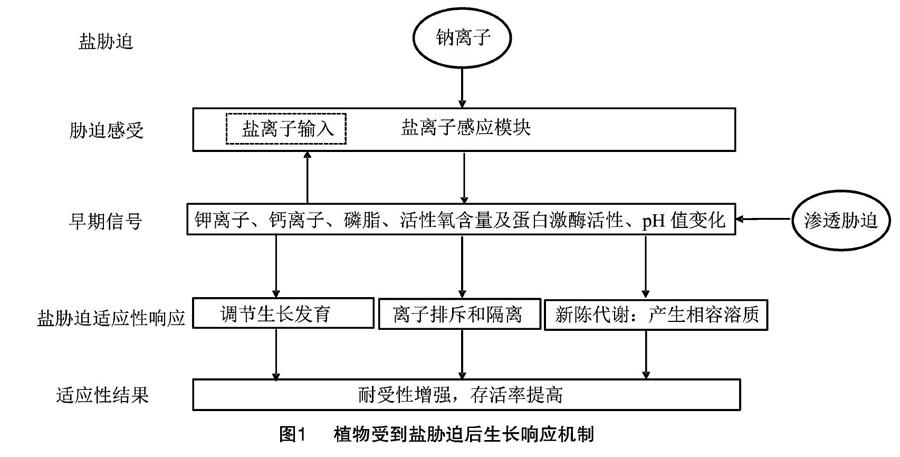
高盐胁迫下,柳树在生长过程中也会产生一系列生理生化响应,目的是维持柳树水利用率、保护关键的生长形态以及光合能力。柳树能在盐环境中良好地生存,得利于其具有克服盐胁迫造成的离子毒害、渗透胁迫和氧化胁迫的调节能力。目前,柳树耐盐机制的研究主要集中在渗透调节、抗氧化酶系统方面。在盐胁迫下,植物的渗透调节发生改变,体内钠离子浓度升高,从而植物感知到盐脅迫。Na+能诱导特异的下游反应,但植物的Na+敏感机制尚不清楚。盐胁迫感知可以发生在Na+输入之前或之后,也可以发生在细胞内或细胞外。在最初感知盐胁迫后,早期的信号反应也被诱导,包括K+转运、Ca2+信号传导、H+转运、磷脂修饰、活性氧诱导和蛋白激酶活性。反过来,这些早期Na+诱导的信号减少了Na+的进入。接着,盐诱导会引发适应性反应,如调节生长发育、离子运输和产生相容的溶质来补偿Na+的渗透压。部分早期信号反应与渗透胁迫诱导的通路重叠。信号链中的每一步都对盐分含量变化产生反应,最终植物在盐碱地上生存[32](图1)。
2.1 渗透调节物质的积累
通常在盐渍土壤中植物根系的水分利用率会明显降低。因此,植物受到盐胁迫后,首先会由渗透胁迫引起植物缺水。植物细胞会积累可溶性渗透物(脯氨酸、可溶性糖)以调节其渗透势。脯氨酸是一种重要的渗透调节物质,在植物细胞中以自由态存在,分子量低,在水中的溶解度高,相对无毒[36]。张健等比较3种柳树的耐盐性,结果显示在盐处理后柳树的脯氨酸和可溶性蛋白含量均高于处理前,而L0906叶片的可溶性蛋白含量高于处理前,但是脯氨酸含量有所下降,耐盐性越强的柳树品种渗透调节物质积累越多[10]。武香研究3种柳树的幼苗在盐处理下渗透调节物质的含量变化,结果表明,盐胁迫后,可溶性蛋白含量增加;根茎叶Na+和Cl-质量分数明显增加,但柳树K+含量均明显下降[37]。在不同柳树品种中,不同渗透调节物质具有明显的差异。在植物耐盐机制中同样起到渗透调节作用的还有甘氨酸甜菜碱(简称甜菜碱),在盐胁迫下,细胞内甜菜碱的积累有助于植株抵抗一定的高盐危害,保护细胞质膜的完整性。已经有研究证实,与甜菜碱合成相关的柳树盐响应BADH基因在盐处理下表达量增加[38]。
2.2 抗氧化酶活性的提高
植物体内的活性氧(ROS)和自由基含量会因植物受到盐胁迫等非生物胁迫而急剧增加,过多的活性氧会破坏生物膜结构,加剧膜的过氧化作用,过量积累MDA,最终会造成细胞死亡。植物会通过自身的抗氧化系统清除ROS和自由基来防止细胞的氧化损伤。同样地,柳树通过超氧化物歧化酶(SOD)、过氧化物酶(POD) 、过氧化氢酶(CAT)等酶和抗氧化物协同作用来减轻盐胁迫造成的氧化伤害。张健等在研究盐处理下3个柳树品系生长指标时发现,处理后耐盐性较好的L0911 MDA含量的增幅要小于其他品系[10]。有效清除植物体内的ROS、SOD可催化超氧自由基产生过氧化氢,减少ROS积累,从而减轻活性氧的伤害[39]。在盐胁迫试验中,韩彪等发现,低浓度的盐处理诱发柳树产生自我保护机制,此时SOD活性增强,随着盐浓度增加,SOD活性开始降低[40]。与SOD一样,POD也是一种抗氧化酶,在植物受到盐害时能减轻细胞由于膜脂过氧化引起的损伤[41]。柳树的抗氧化物质的变化对维持植物渗透稳定有调节作用。
3 柳树的耐盐品种
柳树是耐盐优势树种,其种类多、分布广,不同柳树的耐盐指数有显著差异。例如,来自山东的旱柳9901在盐胁迫后萌枝长度、总根长、叶鲜质量、叶干质量等指标都高于江苏地区的敏盐柳沿江柳[42];盐柳1号来源于沙漠地带的灌木北沙柳,而渤海柳2号来源于旱柳实生选种,二者具有不同的抗盐碱能力,盐柳1号抗盐碱能力强于渤海柳2号[43]。
柳树的内在变异性高,天然杂交现象在柳树生长中很常见,因此柳树杂种是柳树物种多样性的主要原因。目前许多研究调查柳树耐盐性并进行品种挖掘和比较(表1)。在1‰ NaCl质量分数处理下,J172、J795、J799、J842、JW96柳树无性系的根系生长受到一定的刺激[21];在低盐处理时,柳树品种都能承受一定的耐盐性,随着盐浓度的增加,品种间耐盐差异开始显现[23]。陈建建收集了356份柳树种质资源,对其中的67份柳树进行了水培及盆栽试验,将其耐盐能力划分为较强、中等、较差3个等级水平[44]。
4 柳树耐盐相关基因的研究
采用现代分子生物学新技术,将植物的耐盐基因克隆并转化到重要的经济植物中,培养耐盐的转基因植物。目前,国内外学者将已知的盐响应基因在柳树中找到同源基因,通过定量聚合酶链式反应(PCR)找到基因表达量与盐胁迫的某种联系,通过异源表达,在高盐度条件下测试转基因植物的性能(表2)。
随着科学技术的发展,单从某个方面无法解释全部的生物学问题,须要多角度去研究柳树基因、蛋白及分子间的相互作用。有研究者采用比较蛋白质组学方法提取了盐处理下旱柳根、茎、叶的蛋白质,分别得到42、17、19个差异蛋白点,这些差异蛋白参与了12种代谢途径和过程,通过实时定量PCR发现病程相关蛋白(PR)非常丰富并且受盐胁迫上调,表明PR蛋白与旱柳的耐盐能力有关。后来,通过转基因拟南芥进一步验证了SmPR10基因在转基因植株的耐盐性中起着重要作用[58]。盐胁迫下盐敏感型品种JW9-6和耐盐品种JW2372幼苗根部的总蛋白试验,成功鉴定了109个差异表达蛋白,得出结论:代谢过程和蛋白质折叠(BP)、胞质和细胞(CC)以及辅因子、辅酶结合和异构酶活性(MF)这几个功能之间的差异受根蛋白的调节,磷酸葡萄糖异构酶、ATP合成酶ε亚基、磷酸甘油酸激酶、S-腺苷甲硫氨酸合成酶、腺苷激酶5种蛋白质可能是与盐胁迫相关的植物育种候选蛋白质[59]。
田雪瑶等以良种苏柳2345的转录组数据为基础,克隆得到2个柳树NAC转录因子基因,结果发现,2个基因在盐胁迫、赤霉素(GA)和脱落酸(ABA)等逆境模拟处理下,在叶片中特异性表达[54]。抗逆转录因子基因家族的研究是了解抗逆分子机制的重要基础。Jia等将沙柳中的SpRLCK1基因转入拟南芥中,结果证明在同等盐胁迫下具有过表达SpRLCK1的转基因株系幼苗的根长、鲜质量、存活率和相对含水量均高于未转化的野生型植株,POD、SOD、CAT活性均明显高于野生型株系,SpRLCK1的过表达通过增加抗氧化酶的活性来改善转基因植物的耐盐性[57]。为了对旱柳的耐盐分子机制有更多的理解,有学者利用微阵列技术分析盐胁迫下旱柳的差異表达基因,共鉴定出403个基因,功能分类分析表明,这些基因属于编码蛋白家族,涉及代谢、转录调控、信号转导、激素反应、非生物胁迫反应等与生长发育相关的过程[60]。
5 展望
土壤盐碱化程度的加剧、土地资源的紧缺威胁着生产、生态、生存。植物修复是改良土壤盐碱化的有效措施。柳树具有耐盐、速生、耐旱、形态优美诸多优点,使其成为改善环境的优势树种。对于柳树耐盐性的研究已经从表型和生理研究逐渐递进到分子层面的探索,运用先进的科学技术对优良柳树耐盐基因资源进行收集、保存、开发、利用,对于丰富柳树耐盐基因库、保持柳树物种的多样性,培育优良品种、满足林业生产需求都有深远意义。柳树的耐盐性不是单一基因调控的,是由多基因共同发挥功能,所以最好能建立柳树完善的遗传表达系统,实现耐盐性的同源表达,为沿海耐盐林木和沿海滩涂开发提供有利资源。
参考文献:
[1] Munns R,Tester M. Mechanisms of salinity tolerance[J]. Annual Review of Plant Biology,2008,59:651-681.
[2]楼锦花,胡 建,葛 云,等. 苏北滨海盐碱土壤性质演化及其对脱盐规律的启示[J]. 海岸工程,2018,37(3):34-42.
[3]张 静. 黄河三角洲重盐碱区植被耐盐性与绿化技术研究[D]. 青岛:中国海洋大学,2010.
[4]方升佐,黄宝龙. 瑞典柳树能源林的研究及发展概况[J]. 世界林业研究,1997(3):67-72.
[5]潘明建. 柳树的遗传改良及栽培技术[J]. 林业科技开发,2004,18(3):3-7.
[6]孙兆祜,张玉霞,梁群宝. 几种柳树抗逆性调查初报[J]. 甘肃林业科技,1999,24(1):3-5.
[7]王 英. 盐碱土地区绿化植树的措施与建议[J]. 中国农学通报,2004,20(6):96-98.
[8]袁玉欣,王印肖,刘柄响,等. 木本植物耐盐选育研究进展[J]. 河北林业科技,2006(增刊1):31-35.
[9]李 晶,王福森,李树森,等. 3个柳树新品种耐盐性试验研究[J]. 防护林科技,2012(4):73-74.
[10]张 健,李 敏,李玉娟,等. 沿海滩涂耐盐柳树能源林造林试验研究[J]. 林业实用技术,2012(11):32-33.
[11]马赞留,张 健,戴云新,等. 江苏沿海滩涂耐盐柳树育苗技术[J]. 现代园艺,2013(5):42-43.
[12]李小艳. 七种柳树对NaCl盐胁迫的生长生理响应[D]. 呼和浩特:内蒙古农业大学,2018.
[13]Dimitriou I,Aronsson P,Weih M. Stress tolerance of five willow clones after irrigation with different amounts of landfill leachate[J]. Bioresource Technology,2006,97(1):150-157.
[14]张建锋,孙启祥,Makeschin F. 盐胁迫对柳树新无性系苗木生长和土壤酶活性的影响[J]. 水土保持学报,2005,19(3):125-129.
[15]田生昌,张永宏,武文渊. 土壤盐分对两个品种柳树生长发育的影响[J]. 西北林学院学报,2015,30(6):126-129.
[16]季永华. 海滨湿地杨树、柳树新无性系苗期耐盐性研究[J]. 江苏林业科技,2005,32(4):1-4.
[17]隋德宗. 盐胁迫对柳树无性系幼苗生长影响的研究[D]. 南京:南京林业大学,2006.
[18]Isayenkov S V,Maathuis F J M. Plant salinity stress:many unanswered questions remain.[J]. Frontiers in Plant Science,2019,10:1-11.
[19]Cuartero J,Bolarin M C,Asins M J,et al. Increasing salt tolerance in the tomato[J]. Journal of Experimental Botany,2006,57(5):1045-1058.
[20]Shabala S,Wu H H,Bose J. Salt stress sensing and early signalling events in plant roots:current knowledge and hypothesis[J]. Plant Science,2015,241:109-119.
[21]隋德宗,王保松,施士争. 盐胁迫对5个柳树无性系幼苗根系生长发育的影响[J]. 江苏林业科技,2007,34(4):5-8.
[22]刘 铎,丛日春,党宏忠,等. 盐胁迫对柳树新品系的抗氧化酶活性及过氧化物含量的影响[J]. 种子,2014,33(10):20-23.
[23]聂莉莉,张 越,刘仲齐. 盐胁迫对柳树幼苗生长及生理特性的影响[J]. 天津农业科学,2010,16(3):20-23.
[24]Shelden M C,Gilbert S E,Tyerman S D. A laser ablation technique maps differences in elemental composition in roots of two barley cultivars subjected to salinity stress[J]. Plant Journal,2020,101(6):1462-1473.
[25]Silva E N, Silveira J A G, Rodrigues C R F, et al. Physiological adjustment to salt stress in Jatropha curcas is associated with accumulation of salt ions, transport and selectivity of K+, osmotic adjustment and K+/Na+ homeostasis[J]. Plant Biology (Stuttgart, Germany), 2015,17(5):1023-1029.
[26]Jeschke W D,Stelter W. Mesurement of longitudinal ion profiles in single roots of Hordeum and Atriplex by use of flameless atomic absorption spectroscopy[J]. Planta,1976,128:107-112.
[27]王 宁,周晓星,刘俊祥,等. 盐胁迫对柳树无性系SH31离子含量及光合作用的影响[J]. 林业科学研究,2015,28(4):565-569.
[28]Mirck J,Volk T A. Mass balances and allocation of salt ions from Solvay storm water for shrub willow (Salix spp.)[J]. Biomass and Bioenergy,2012,39:427-438.
[29]张梦如,杨玉梅,成蕴秀,等. 植物活性氧的产生及其作用和危害[J]. 西北植物学报,2014,34(9):1916-1926.
[30]Janero D R. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury[J]. Free Radical Biology and Medicine,1990,9(6):515-540.
[31]Santos V A H F D,Ferreira M J,Rodrigues J V F C,et al. Causes of reducedleaf-level photosynthesis during strong El Nio drought in a Central Amazon forest[J]. Global Change Biology,2018,24(9):4266-4279.
[32]van Zelm E,Zhang Y,Testerink C. Salt tolerance mechanisms of plants[J]. Annual Review of Plant Biology,2020,71:403-433.
[33]乔志攀. 灌木柳耐盐种质筛选及其耐盐机理初探[D]. 南京:南京林业大学,2015.
[34]周 鹏,陈庆生,张 敏,等. 灌木柳叶PSⅡ对盐胁迫的响应及耐盐性[J]. 东北林业大学学报,2014,42(9):98-101,106.
[35]季琳琳,吴中能,刘俊龙,等. NaCl胁迫对2种柳树幼苗光合特征的影响[J]. 河北农业大学学报,2013,36(1):50-54.
[36]李业宇,曹帮华,倪智雄,等. 柳树耐盐性及良種选育研究进展[J]. 山东林业科技,2018,48(2):120-126.
[37]武 香. 盐胁迫下植物的渗透调节及其适应性研究[D]. 北京:中国林业科学研究院,2012.
[38]李 敏,张 健,冯立国,等. 柳树液泡膜ATP酶B亚基基因克隆及在盐胁迫下的表达分析[J]. 江苏农业学报,2013,29(5):1149-1153.
[39]Wilson J P,White J D,Montaez I P,et al. Carboniferous plant physiology breaks the mold[J]. New Phytologist,2020,227(3):667-679.
[40]韓 彪,解孝满,董 昕,等. 盐胁迫对柳树无性系幼苗生理生态特性的影响[J]. 山东林业科技,2013,43(3):9-12.
[41]陈少裕. 膜脂过氧化对植物细胞的伤害[J]. 植物生理学通讯,1991,27(2):84-90.
[42]李 敏,王 莹,李玉娟,等. 2个柳树亲本耐盐性比较试验[J]. 浙江农业科学,2017,58(7):1220-1222.
[43]田生昌,李月祥. 不同含盐量土壤对盐柳1号苗木生长影响[J]. 西北林学院学报,2018,33(1):120-125.
[44]陈建建. 柳树种质资源收集及抗盐碱性评价[D]. 泰安:山东农业大学,2015.
[45]郭 聪,张 健,马祥建,等. 15种柳树苗期耐盐性评价[J]. 上海农业学报,2020,36(2):66-71.
[46]李佳迪,刘 铎,李子英,等. 5个柳树无性系苗期耐盐性综合评价及鉴定指标的筛选[J]. 西北林学院学报,2020,35(3):114-120.
[47]王 辉. 杨柳树SSR遗传多样性分析及4种柳树耐盐性比较研究[D]. 保定:河北农业大学,2008.
[48]张 健,李 敏,李玉娟,等. 盐胁迫下3个柳树新品系生理指标的变化[J]. 江苏农业科学,2012,40(8):197-199.
[49]王伟伟,乔志攀,何旭东,等. 灌木柳种质资源的耐盐性变异[J]. 江苏林业科技,2016,43(6):15-19.
[50]施士争,隋德宗,王红玲,等. 灌木柳速生无性系的耐盐性选择研究[J]. 西北林学院学报,2010,25(4):72-77.
[51]李 敏,张 健,王奎山,等. 耐盐柳树BADH基因克隆及表达分析[J]. 江苏农业学报,2013,29(3):485-489.
[52]余春梅,李 敏,何新雨,等. 过量表达柳树两个SVP1s基因提高拟南芥抗盐胁迫能力[J]. 中国生物化学与分子生物学报,2015,31(4):414-421.
[53]周 洁,宋雪晴,何旭东,等. 柳树SjMIPS基因的克隆及其表达分析[J]. 江苏林业科技,2016,43(6):1-5.
[54]田雪瑶,周 洁,王保松,等. 柳树NAC基因的克隆与表达模式分析[J]. 南京林业大学学报(自然科学版),2020,44(1):119-124.
[55]Han X J,He X L,Qiu W M,et al. Pathogenesis-related protein PR10 from Salix matsudana Koidz exhibits resistance to salt stress in transgenic Arabidopsis thaliana[J]. Environmental and Experimental Botany,2017,141:74-82.
[56]周 洁,黄 婧. 柳树几丁质酶基因SlChi的克隆和功能验证[J]. 分子植物育种,2018,16(24):8013-8021.
[57]Jia H X,Li J B,Zhang J,et al. The Salix psammophila SpRLCK1 involved in drought and salt tolerance.[J]. Plant Physiology and Biochemistry,2019,144:222-233.
[58]Qiao G R,Zhang X G,Jiang J,et al. Comparative proteomic analysis of responses to salt stress in Chinese willow (Salix matsudana Koidz)[J]. Plant Molecular Biology Reporter,2014,32:814-827.
[59]Dezong S,Baosong W,Lian X,et al. Identification of candidate proteins and networks related to salinity stress in shrub willow roots by comparative proteomic analysis[J]. Pakistan Journal of Botany,2017,49(5):1627-1640.
[60]Liu M Y,Qiao G R,Jiang J,et al. Identification and expression analysis of salt-responsive genes using a comparative microarray approach in Salix matsudana[J]. Molecular Biology Reports,2014,41(10):6555-6568.
