葡萄NF-Y核因子家族的鉴定与表达分析
2021-07-16马维峰韩爱民乃国洁李艳梅李彦彪
马维峰,韩爱民,乃国洁,李艳梅,李彦彪,毛 娟
(甘肃农业大学 园艺学院,兰州 730070)
20世纪90年代以来,农业节水生物技术领域的研究工作集中在模式植物拟南芥上,也通过基因转移和重组,获得了小麦(Triticumaestivum)[1]、玉米(Zeamays)[2]和水稻(Oryzasativa)[3]的耐旱性材料,但葡萄的耐旱基因工程还鲜有报道,而核因子Y (nuclear factor Y, NF-Y)就是其研究的热点基因。目前,NF-Y已在多个物种中进行了成员鉴定和功能验证,如拟南芥(Arabidopsisthaliana)[4]、水稻[5]、小麦[6]、胡桃(Juglansregia)[7]、香蕉(Musanana)[8]、桃(Prunuspersica)[9]、高粱(Sorghumbicolor)[10]等大量植物上。核因子又称作反式作用因子(Trans-acting factor)和转录因子(Transcription factor,TF),是一组能够与顺式作用元件(启动子、增强子、调控序列和可诱导元件等)发生特异性结合[11],通过它们之间以及其他相关蛋白之间的相互作用,参与调节靶基因转录,并使之以特定的强度在特定的时间和空间表达的结合蛋白,在植物光合作用,开花调控和逆境胁迫的响应等方面具有重要作用[12]。
NF-Y是由 NF-YA(又称 CBF-B 或 HAP2)、NF-YB(又称 CBF-A 或 HAP3)和 NF-YC(又称 CBF-C 或 HAP5)组成的异源三聚体[13],并与 CCAAT-box结合(25%真核生物基因的启动子区都含的一种顺式作用元件)。NF-YA和NF-YC亚基包含一个富含谷氨酸的结构域和一个疏水残基,对转录激活性起到重要作用[14]。在动物和酵母中,每个 NF-Y 亚基都是由单拷贝基因编码的,而在植物拟南芥整个基因组中有10个 NF-YA 编码基因,13个NF-YB 编码基因和13个NF-YC编码基因[15]。Donald等[2]研究表明在拟南芥和玉米中过表达NF-YB1可以显著地提高植株的抗旱性,在干旱条件下获得显著高于对照的产量。对拟南芥NF-YA5核因子的研究发现,干旱胁迫和脱落酸(ABA)处理下NF-YA5被强烈诱导表达,过表达NF-YA5的转基因拟南芥植株比正常的野生型表现出叶片水分损失量少,对干旱的抗性显著增强等特性。并且发现NF-YA5序列上存在一个microRNA(miR169)的靶位点,而 miR169的表达受干旱抑制[16]。与许多植物基因家族一样,NF-Y基因家族的结构经历了许多基因重复,导致一组紧密相关的同源基因,这些同源基因可能具有重叠的表达模式,或者可能已经经历了亚功能化,呈现特定的组织发育相关表达模式。
本试验利用生物信息学方法和荧光定量PCR,对葡萄NF-Y家族的理化性质、蛋白质二级结构预测、亚细胞定位预测、染色体定位、基因结构进行分析研究。初步探索该基因在葡萄逆境胁迫中的表达特点和响应机制规律,为葡萄NF-Y的功能验证提供参考。
1 材料与方法
1.1 试验材料与处理
以甘肃农业大学园艺学院保存的‘黑比诺’(VitisviniferaL. cv.‘Pinot Noir’)试管苗为试验材料,配制MS培养基,pH 5.8~6.0,将试验材料的单芽茎段进行继代培养,采用纸桥法进行固定,光照16 h,温度25 ℃,黑暗8 h,温度为 20 ℃,放置在LED白光下继代培养30 d后选择生长一致且无污染的葡萄试管苗,用无菌注射器吸出原培养基后分别加入50 mL含有10%PEG、500 μmol·L-1ABA、200 mmol·L-1NaCl的MS液体培养基进行处理,每组处理设置3个生物学重复,对照为等体积蒸馏水处理,对照与PEG、ABA、NaCl处理时长为24 h。试验于2019年3月-7月于甘肃农业大学园艺学院果树生理与生物技术开放实验室进行。
1.2 葡萄NF-Y转录家族序列数据获得
从拟南芥基因组数据TAIR(https://www.arabidopsis.org)下载拟南芥NF-Y核因子的氨基酸序列,在葡萄基因组数据库中经同源基因比对,共搜索到40个葡萄NF-Y家族成员,分别下载基因的全长,CDS序列,氨基酸序列以及cDNA序列,并对基因全长与CDS的数量进行统计。根据NF-Y基因家族特定的结构区域,利用在线网站(http://smart.embl-heidelberg.de/)和Pfam进行保守域预测,删除其结构域缺失或不完整的序列片段。
1.3 葡萄NF-Y转录因子家族理化性质、二级结构、亚细胞定位和系统进化分析
利用ExPASy数据库(https://web.expasy.org/protparam/)中的ProParam工具进行蛋白质的理论等电点、氨基酸个数、分子量的统计。利用Predict Protein(https://www.predictprotein.org)在线分析蛋白质二级结构;利用WOLF PSORT(https://wolfpsort.hgc.jp)在线分析葡萄NF-Y基因家族的亚细胞定位预测。使用MEGE 7.0软件构建NF-Y转录因子家族系统发育树。
1.4 葡萄NF-Y转录因子家族motif、基因结构、染色体定位和多序列比对
利用MEME(http://meme-suite.org/)在线程序分析葡萄NF-Y转录因子家族保守基序,预测基序的数量设置为10个;在线软件GSDS 2.0(http://gsds.cbi.pku.edu.cn/)进行基因结构分析;使用DNAMAN进行结构域序列多重比对,利用TBtools进行染色体定位及共线性分析。
1.5 荧光定量 PCR 分析
将葡萄 NF-Y家族基因的 CDS 序列进行在线引物设计(表1)[生工生物工程(上海)股份有限公司网站],将设计好的引物交由该公司完成。cDNA用 Prime Script RT reagent Kit(Perfect Real Time)试剂盒(TaKaRa,日本)并按其说明进行。反转录产物在-20 ℃下保存备用。实时荧光定量 PCR(qRT-PCR)用 Light Cycler○R96 Real-Time PCR System(Roche,瑞士)实时定量 PCR 仪,用上述设计的引物进行PCR 扩增,以葡萄GAPDH基因为内参,对葡萄NF-Y基因家族进行特异性表达分析。扩增体系含为20 μL,分别含cDNA 3 μL,上、下游引物各0.8 μL,SYBR MIX 10 μL,ddH2O 5.4 μL,反应程序为:95 ℃ 30 s,95 ℃10 s,58 ℃ 30 s,72 ℃ 20 s共40个循环,3 次重复。

表1 葡萄NF-Y表达分析的实时荧光定量引物Table 1 qRT-PCR primers for expression analysis of NF-Y in grape
1.6 数据分析
采用 Excel 2010和SPSS 23软件分析试验数据,用TBtools v1.0692和Origin 9.0作图。
2 结果与分析
2.1 葡萄NF-Y家族成员理化性质分析
利用拟南芥核因子序列在葡萄基因组网中搜索,通过同源比对检索鉴定出的VvNF-Y家族核因子分为VvNF-YA、VvNF-YB和VvNF-YC三个亚族,其中VvNF-YA包含5个核因子,命名为VvNF-YA1~VvNF-YA5,VvNF-YB包含13个核因子,命名为VvNF-YB1~VvNF-YB13,VvNF-YC包含6个核因子,命名为VvNF-YC1~VvNF-YC6。由表2可知,葡萄NF-Y核因子家族氨基酸大小为132~345 aa,VvNF-YA1氨基酸序列最长,VvNF-YB7的氨基酸序列最短。分子质量集中在14.56~37.70 ku。等电点集中在4.62~9.83,其中VvNF-YB10的等电点最低,VvNF-YC6的等电点最高。

表2 葡萄 NF-Y家族核因子蛋白质理化性质Table 2 Physicochemical properties of NF-Y nuclear factor family in grape
2.2 葡萄NF-Y家族成员亚细胞定位和蛋白二级结构分析
亚细胞定位分析显示葡萄24个NF-Y基因均定位于细胞核(表3),且集中表达于细胞核与叶绿体中,而在线粒体、内质网、细胞膜和细胞质中有少量表达,此外过氧化物酶体中只有VvNF-YB6表达。另外5个蛋白 VvNF-YB2、VvNF-YA1、VvNF-YB4、VvNF-YA2、VvNF-YA4的最大积分值均为13;7个蛋白VvNF-YB3、 VvNF-YA3、VvNF-YB5、VvNF-YB6、VvNF-YB9、 VvNF-YB11、VvNF-YA5的最大积分值均为14;VvNF-YC4的最大积分值为7;VvNF-YC3的最大积分值为3,该14个蛋白均定位于细胞核中,其中VvNF-YC3的最大积分值也出现于叶绿体和细胞质中。6个蛋白VvNF-YC2、VvNF-YB7、VvNF-YB8、 VvNF-YB10、VvNF-YB12和VvNF-YC6的最大积分值分别为5.5、7、7、4、9和7,均定位于线粒体中,其中 VvNF-YB10的最大积分值也出现于叶绿体中。2个蛋白VvNF-YC2和VvNF-YB13最大积分值分别为7和5,定位于细胞质中。VvNF-YB1和VvNF-YC5的最大积分值分别为6和5.5,分别定位于胞外和细胞膜中。
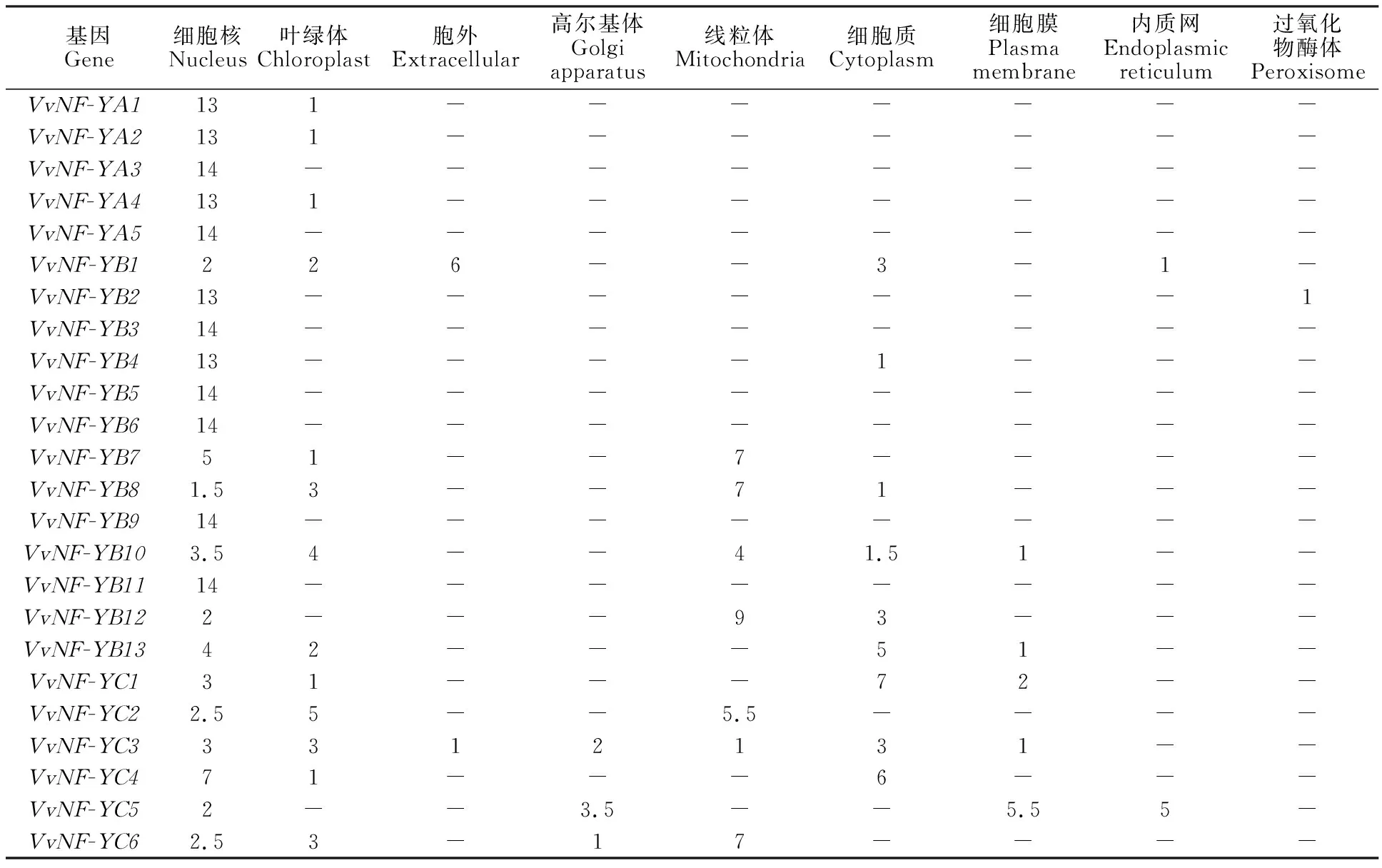
表3 葡萄 NF-Y 核因子亚细胞定位预测Table 3 Subcellular location prediction of NF-Y in grape
蛋白质的二级结构主要以α-螺旋、β-折叠、β-转角、延伸链和不规则卷曲构成,通过EXPASy提供的在线SOPMA程序对葡萄NF-Y核因子家族蛋白二级结构进行分析得出(表4),24个NF-Y蛋白主要以α-螺旋(Alpha helix)(13.04%~66.91%)、β-转角(Beta turn)(0.95%~8.42%)和不规则卷曲(Random coil)(20.15%~ 77.68%)3种结构构成。其中A亚族α-螺旋占 13.04%~29.12%,β-转角占2.03%~7.08%,不规则卷曲占54.95%~77.68%;B亚族α-螺旋占35.35%~63.96%,β-转角占0.95%~8.37%,不规则卷曲占24.87%~57.82%;C亚族α-螺旋占42.31%~66.91%,β-转角占1.65%~8.42%,不规则卷曲占20.15%~52.20%。可见,VvNF-YB和VvNF-YC亚族α-螺旋、β-转角和不规则卷曲所占比例相近,但远高于VvNF-YA亚族。

表4 葡萄NF-Y蛋白二级结构分析Table 4 Secondary structure of NF-Y protein sequence in grape
2.3 葡萄NF-Y核因子家族进化分析
为进一步了解VvNF-Y核因子家族的进化特性,利用葡萄、拟南芥和水稻共94个NF-Y核因子蛋白序列构建NF-Y核因子家族系统进化树(图1),可见葡萄NF-Y核因子与拟南芥NF-Y核因子同源性较高,与水稻NF-Y核因子同源性较低。94个成员可分为Ⅰ、Ⅱ和Ⅲ 3个亚族,分别包含38、30和26个核因子。进一步分析发现NF-YA亚族核因子全部分布于第Ⅰ亚族中,NF-YC亚族核因子全部分布于第Ⅱ亚族中,NF-YB亚族核因子全部分布于第Ⅲ亚族中。此外Ⅰ亚族中包含1对直系同源核因子和7对旁系同源核因子,Ⅱ亚族中含有2对直系同源核因子和9对旁系同源核因子,Ⅲ亚族中含有7对直系同源核因子和1对旁系同源核因子。进一步分析发现葡萄NF-Y蛋白序列单独聚类形成的进化树与此相一致,也分为Ⅰ、Ⅱ和Ⅲ3个亚族(图2)。
2.4 葡萄NF-Y核因子家族保守结构域特征和基因结构分析
葡萄NF-Y基因的结构分析表明,24个NF-Y核因子的结构差异明显,如图2-A所示,其所含内含子数0~5,外显子数1~6。第Ⅰ亚族中NF-YB6和NF-YB8外显子数最多,均为5个,NF-YB5、NF-YB7、NF-YB9和NF-YB11等4个核因子内含子数目最少,均为1个。且NF-YB1、NF-YB7、NF-YB9和NF-YB11均不含上下游编码区。第Ⅱ亚族中VvNF-YA1和VvNF-YA5内含子和外显子数目相同,分别有4个、5个,VvNF-YA3和VvNF-YA4内含子和外显子数目相同,分别有3个、4个,VvNF-YA2只有1个内含子和2个外显子。第Ⅲ亚族中VvNF-YC3和VvNF-YC6外显子数目相近,二者具有相似性,而VvNF-YC1只有1个外显子,没有下游编码区,与VvNF-YC2和VvNF-YC3差异较大。另外VvNF-YC2外显子最多,有6个,VvNF-YC4外显子最少,仅有1个。
葡萄NF-Y家族蛋白含有10个保守基序(图2-B),每个基序含有14~54个氨基酸(表5),其中Motif1、Motif3、Motif5和Motif6都包含54个氨基酸,Motif7最少,为14个。24个NF-Y中NF-YB5、NF-YB9和NF-YB11所具有最多的Motif,为5个,而NF-YA3、NF-YA5和NF-YB13只有1个Motif,此外,分析发现保守基序Motif1具有较高的保守性。

表5 葡萄NF-Y蛋白氨基酸保守序列Table 5 Conserved motifs of NF-Y proteins in grape
2.5 葡萄 NF-Y 家族基因的染色体定位及共线性分析
从葡萄基因组数据库中下载NF-Y基因家族成员的位置和该基因所在染色体的全长数据,发现VvNF-YA5、VvNF-YB11~VvNF-YB13和VvNF-YC5~VvNF-YC6这6个基因位于未知染色体上。将VvNF-Y的24个基因利用TBtools绘制NF-Y 家族基因在染色体上的分布图。由图3可知,葡萄24条NF-Y基因分别位于12条染色体和未知染色体上,较为分散。其中未知染色体上分布有6个,6号和19号染色体上各分布3个,1号、5号和13号染色体各分布2个基因。7、8、9、12、14和17号染色体各分布1个,且除12号染色体和未知染色体外,NF-Y基因都位于染色体末端,而其余染色体则没有NF-Y分布。
为进一步了解VvNF-Y基因家族的进化作用力,从在线网站(http://tools.bat.infspire.org/circoletto/)分析出identity>75%的8对葡萄NF-Y基因对(图3)进行同义突变频率(Ka)及非同义突变频率(Ks)值的计算,并获得葡萄基因的突变频率比值(Ka/Ks)(表6)。8对基因的Ka均小于1;VvNF-YB5和VvNF-YB11的Ks最大,为50.925 1,VvNF-YA2和VvNF-YA4最小,为0.074 6。另外,Ka/Ks对基因的进化选择有重要的指导作用:Ka/Ks>1,认为受到正向选择作用;Ka/Ks=1,认为受到中性选择作用; Ka/Ks<1,则认为受到纯化选择作用。本研究中所有基因对的Ka/Ks均小于0.5,认为基因对之间存在纯化选择作用。

表6 葡萄NF-Y同源核因子Ka/Ks分析Table 6 Ka/Ks analysis of VvNF-Ys homologous nuclear factor
2.6 葡萄NF-Y家族基因多序列比对分析
为了进一步研究VvNF-Y基因家族成员的保守区,利用DNAMAN 软件对其氨基酸序列进行多序列比对分析,发现VvNF-Y的3个亚族均含有保守区(图4)。VvNF-YA蛋白含有1个DNA结合结构域(DNA Binding)和1个NF-YB/YC亚基互作结构域(NF-YB/YC Interaction);VvNF-YB蛋白包含1个DNA结合结构域、1个NF-YA亚基互作结构域(NF-YA Interaction)和2个NF- YC亚基互作结构域(NF-YC Interaction);VvNF-YC蛋白具有1个DNA结合结构域、2个NF- YA亚基互作结构域(NF- YA Interaction)和1个NF-YB亚基互作结构域(NF-YB Interaction)。
2.7 葡萄NF-Y家族基因顺式作用元件分析
本研究利用在线软件PlantCARE分析了葡萄NF-Y起始密码子上游2 000 bp的碱基序列,并利用TBtools绘制顺式作用元件图(图5)。在葡萄NF-Y基因家族中鉴定出大量光响应元件如G-Box、Box 4和MRE;多种激素反应元件,包括脱落酸响应元件(ABRE)、生长素(AuxRE和TGA-element)、赤霉素(P-box)和茉莉酸甲酯(CGTCA-motif和TGACG-motif);多种响应环境信号元件,如防御与胁迫(TC-rich repeats)、厌氧诱导(ARE)、低温(LTR)和干旱诱导(MBS)以及水杨酸响应元件(TCA-element),由此可知其表达调控机制复杂。
2.8 葡萄NF-Y核因子家族qRT-PCR分析
qRT-PCR 结果显示,当VvNF-Y基因家族受到500 μmol·L-1ABA、10% PEG和200 mmol·L-1NaCl的诱导时,VvNF-YA、VvNF-YB和VvNF-YC亚族基因的相对表达量差异显著(图6)。当受到500 μmol·L-1ABA和200 mmol·L-1NaCl处理时,VvNF-YA亚族和VvNF-YC亚族基因中各有1个基因上调表达,分别是VvNF-YA2和VvNF-YC3,尤其VvNF-YC3在500 μmol·L-1ABA处理的条件下其表达量显著上调,为CK的3.31倍,而在VvNF-YB亚族中有VvNF-YB3和VvNF-YB4表达量显著上调,其余各VvNF-Y家族基因表达量显著下调;在10% PEG条件下,VvNF-YA亚族和VvNF-YB亚族中各有2个基因呈上调表达,分别是VvNF-YA1、VvNF-YA2和VvNF-YB3、VvNF-YB4,尤其VvNF-YB3基因的表达量与对照相比呈显著上调,为对照的3.50倍,其余VvNF-YA亚族和VvNF-YB亚族基因表达量均显著下调,而在VvNF-YC亚族基因中仅VvNF-YC3基因显著上调,为对照的5.60倍,其余VvNF-YC亚族基因表达量均显著下调。
3 讨论与结论
NF-Y是NF-YA、NF-YB和NF-YC三个独特亚族成员组成的结合于启动子CCAAT上的异源三聚体蛋白复合物,在植物生长发育过程中表现出重要作用[17]。本研究从葡萄全基因组中鉴定出24个葡萄NF-Y基因家族成员,这与33个毛竹(Phyllostachysedulis)[18]和24个胡桃(Juglansregia)[7]NF-Y家族成员的数量相近,说明通过Blast比对进行基因筛选和MEME结构域缺失或不完整基因的剔除条件较为合理,从而得到的葡萄NF-Y家族成员是可信的。
鉴定出的24个VvNF-Y家族成员中有5个VvNF-YA、13个VvNF-YB和6个VvNF-YC亚族成员。基因定位发现24个VvNF-Y主要定位于细胞核中,部分基因也定位于线粒体、内质网、细胞膜和细胞质中,可见VvNF-Y核因子主要在细胞核中行使其功能。部分基因也可能在线粒体、内质网、细胞膜和细胞质中少量表达。此外葡萄NF-Y核因子家族氨基酸大小在132~345 aa之间,相对分子质量集中在14.56~37.70 ku,等电点为4.62~9.83,这与Mai等[19]的试验结果相似,而在鹰嘴豆(Cicerarietinum)[20]和西瓜(Citrulluslanatus)[21]中NF-Y家族氨基酸残基相对较多,相应分子量也较葡萄NF-Y核因子家族大,但等电点范围相近。系统同进化分析发现葡萄NF-Y可归为3个亚族(VvNF-YA1~5、VvNF-YB1~13和VvNF-YC1~6),进一步分析发现成员关系较近的基因具有相似的基因结构:葡萄NF-Y基因内含子数量少,尤其NF-YB5、NF-YB9、NF-YB11和NF-YC1不含内含子,NF-YB1、NF-YB4、NF-YB7、NF-YB10、NF-YB12和NF-YA2仅具有1个内含子,这可能是由于葡萄NF-Y基因在进化过程中大量内含子丢失,基因结构保持稳定,加速CCAAT-box的转录,促进基因的表达有关[22-23]。
Malviya等[10]将高粱NF-Y基因家族的33个基因归为3个亚族,且进化距离较近的基因其结构也越为相似,其含有少量甚至不含内含子,这与本试验结果相似,在桃[9]和茶树(Camelliasinensis)[24]NF-Y基因家族生物信息学分析中也有类似的结果。多序列比对发现VvNF-Y具有NF-Y家族成员的典型特征[25],具体表现为VvNF-YA、VvNF-YB和VvNF-YC蛋白均含有1个DNA结合结构域,不同的是VvNF-YA含有1个NF-YB/YC亚基互作结构域,VvNF-YB蛋白含有1个NF-YA亚基互作结构域(NF-YA Interaction)和2个NF- YC亚基互作结构域(NF- YC Interaction),VvNF-YC蛋白具有1个NF-YB亚基互作结构域(NF-YB Interaction)和2个NF- YA亚基互作结构域(NF- YA Interaction),这与滕露等[20]和Li等[9]的研究结果一致,但Yan等[8]研究发现香蕉NF-YB亚族基因只含有一个NF-YC亚基互作结构域,这与本试验中VvNF-YB蛋白具有2个NF- YC亚基互作结构域的结果有一定的差异。
大量研究表明植物响应非生物胁迫是通过转录因子结合顺式作用元件进而调控靶基因的表达[26-27],因此本试验对VvNF-Y家族基因上游 2 000 bp区域的启动子顺式作用元件进行了深入分析,发现葡萄NF-Y基因家族具有大量G-Box、Box 4和MRE等光响应元件、脱落酸(ABRE)、生长素(AuxRE和TGA-element)和赤霉素(P-box)等激素响应元件和厌氧诱导(ARE)、低温(LTR)和干旱(MBS)等逆境信号响应元件,推测该核因子表达调控机制复杂,如VvNF-YA2、VvNF-YB3、VvNF-YB4和VvNF-YC3均包含ABRE和MBS作用元件,表现出VvNF-Y核因子家族多个基因均参与了抗旱响应,而且在其他非生物胁迫中也具有重要作用,这已在前人的研究中得到证实,如在谷子[28]中具有抗旱作用元件的SiNF-YA1可增强植株的耐旱性和耐盐性;水稻中NF-Y基因参与了叶绿体的形成,其可抑制水稻hap3基因的表达从而降低光合作用,而且OsNF-YA10基因的表达也会受干旱和盐胁迫的影响[29-30],拟南芥中也发现NF-YA5与多个下游抗旱抗逆基因表达相关[2]。
此外,qRT-PCR分析发现VvNF-YA2、VvNF-YB3、VvNF-YB4和VvNF-YC3在500 μmol·L-1ABA、200 mmol·L-1NaCl和10% PEG处理下表现为显著上调表达,VvNF-YA1在10% PEG处理下也会显著上调表达,推测该NF-Y核因子在受到激素、干旱和盐胁迫时具有一定的调节作用,这与前人的报道NF-Y具备多种生物学功能的研究结果相符,如水稻转录因子OsHAP2E不仅能增加水稻分孽数和光合作用,还能提高对盐、干旱和病原菌等逆境抗性[31],拟南芥AtNF-YA2和AtNF-YA10可提高植株对逆境的抗性[32],毛果杨(P.trichocarpa)PtNF-YA9在其苗期的过表达能够提高其耐盐性和抗旱性[33]。近年来,通过促进核因子的表达来提高植物抗旱性也在水稻、烟草(Nicotianatabacum)等植物中都得到证实[34-37]。
综上可知,葡萄NF-Y核因子进化过程较为保守,且部分NF-Y基因对ABA、PEG和盐害等非生物逆境胁迫响应明显,为进一步研究葡萄NF-Y核因子在非生物胁迫下的功能提供了 参考。
