环境因素对金钟藤种子萌发的影响
2021-07-16张守信周勇芝王华锋薄文浩龙文兴
张守信, 周勇芝, 黄 韦, 韩 婷, 王华锋, 薄文浩, 龙文兴, 周 韬
(1.热带特色林木花卉遗传与种质创新教育部重点试验室,海南大学林学院,海南五指山森林生态系统国家定位观测研究站, 海口 570228;2.海南大学生命科学与药学院, 海口 570228;3.北京林业大学生物科学与技术学院, 北京 100083)
金钟藤(Merremiaboisiana(Gagn.) v. Ooststr)是旋花科鱼黄草属的多年生藤本植物,在中国的分布地主要为海南、云南和广西。金钟藤在疏林地和次生常绿阔叶林地广泛生长,向上缠绕并覆盖,致使乔、灌木大面积死亡,造成严重经济损失。藤茎可以落地生根,迅速蔓延成片[1-2]。2004年底,广州报道了金钟藤对森林植被的巨大破坏[3-4],引起了人们的重视。2005年之后,海南省多个地区都报道存在不同程度的金钟藤危害现象[5-9]。2013年,金钟藤被国家林业局列入《全国林业危险性有害生物名单》。
金钟藤在适宜条件下枝条生长蔓延迅速,但种子繁殖能力弱,野外极少见幼苗[10-11]。何泽等[12]调查发现,野外金钟藤结实率仅为27%。刘海洋[13]发现,金钟藤种子自然萌发率低,进行种皮处理后使得萌发率提高。倪广艳等[14]发现,金钟藤种子的萌发率在30 ℃条件下明显高于28 ℃和25 ℃条件下的萌发率。曾宋君等[15]发现,金钟藤各个组织的提取液不同程度地降低了菜薹种子的萌发率。程汉亭等[16]发现,金钟藤种子提取液抑制白菜种子的萌发和生长。本试验研究光照、高温、干旱和自身化感作用等环境因素对金钟藤种子萌发的影响,期望为金钟藤灾害的预测和防控工作提供理论参考。
1 材料与方法
1.1 供试材料
2020年6月从海南省琼中县黎母山国家森林公园(海拔530 m)采集金钟藤种子。
1.2 试验方法
去除空瘪和畸形种子,取饱满种子用75%酒精浸泡10 min,蒸馏水冲洗;对种皮前端进行切口 (图1 c、d) ,除种皮处理试验的对照组外,本研究中所有其他试验所用种子均经过切损种皮处理;在培养皿中平铺两层滤纸并放入高温灭菌锅灭菌,将种子均匀放置于滤纸上;培养皿中加入纯净水或处理溶液,不断补充纯净水保持滤纸湿润。种子放置于人工气候箱中培养,对照组设定培养温度为26 ℃,光照12 h,纯净水培养。
种皮切损试验中的对照组采用未经切口的完整种子。光照试验中黑暗处理组使用黑布遮光。温度试验中设置梯度分别为28 ℃、30 ℃、32 ℃和34 ℃。干旱胁迫试验使用PEG-6000(聚乙二醇)模拟干旱条件,配制质量分数为5%、10%、15%和20%的PEG溶液处理种子。自化感试验中提取液的配置方法为:剪碎金钟藤5 g叶片,加入50 mL蒸馏水静置提取24 h,最终获得0.1 g·mL-1的提取液,以及稀释后得到0.05 g·mL-1的提取液。除相应处理条件外,处理组其他培养条件与对照组相同。每个处理20粒种子,设置3次重复。
1.3 指标测定
每天10:00时统计发芽数,连续3 d没有新种子发芽视为发芽结束。使用游标卡尺测量发芽种子的胚根长、胚芽长。使用以下公式计算发芽率、发芽势、发芽指数。
发芽率 (%) = (发芽种子数/供试种子总数) ×100%;
发芽势 (%) = (发芽高峰期发芽的种子数/供试种子总数) ×100%;
发芽指数=∑(Gt/Dt),式中:Dt为发芽天数,Gt为t时间内的发芽数。
1.4 数据处理
使用Excel 2016软件进行数据处理和图表制作,进行单因素方差分析和显著性检验。
2 结果与分析
2.1 种皮对金钟藤种子萌发的影响
本次试验中ck种子 (图1 a、b) 直到第5天仍未出现种子萌发现象。处理组将种皮进行了切损处理 (图1 c、d) ,种子发芽率达72% (图1 e、f) ,显著高于ck。结果表明,种皮完整的种子短时间内不能发芽,切损处理可以消除种皮对种子萌发的阻碍作用。
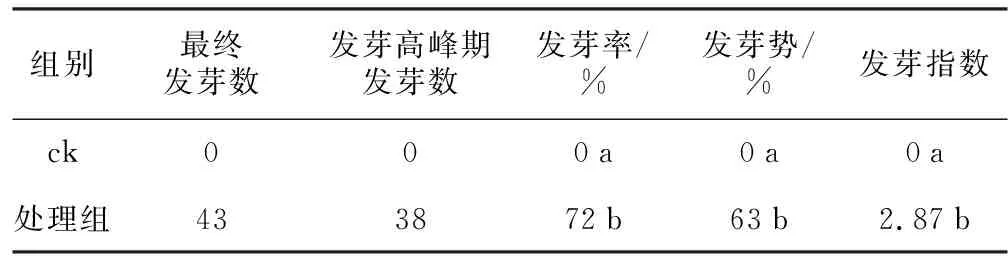
表1 种皮处理对金钟藤种子萌发的影响

注:a、b为对照组正常种子;c、d为处理组切损种皮种子;e、f为已萌发种子。图1 金钟藤种子的种皮处理Fig.1 Cutting treatment of seed coat of M. boisiana seeds
2.2 光照对金钟藤种子萌发的影响
由图2可知,光照条件下与黑暗条件下种子的发芽率、发芽势和发芽指数不存在显著差异,但萌发后种子的胚根和胚芽长度差异显著,光照使得萌发种子对照组的根长比黑暗组增加39.3%,其芽长仅为黑暗组的43.8%。结果说明,光照不会显著影响种子的萌发,但会导致萌发后种子的胚根变长,胚芽变短。
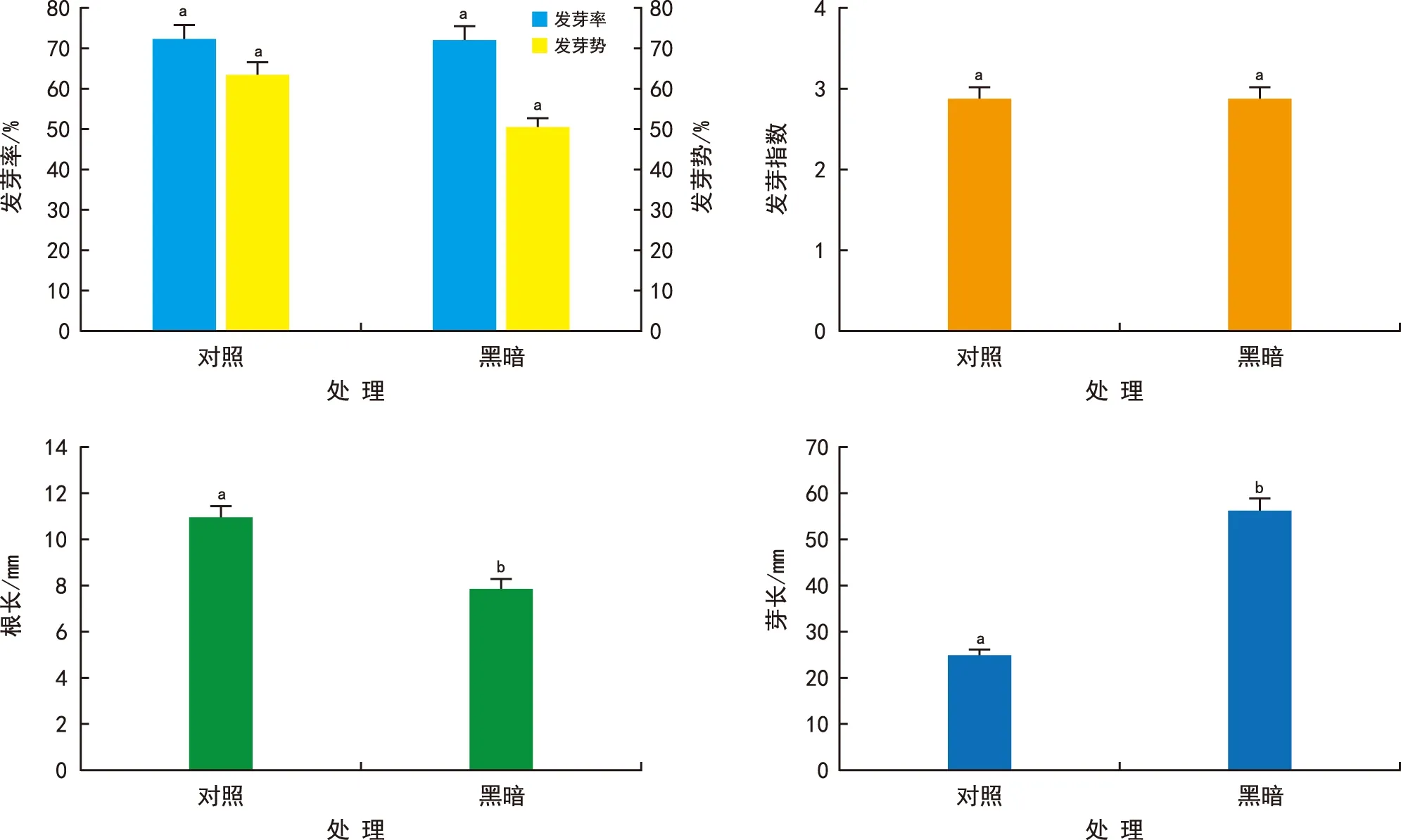
图2 光照对金钟藤种子萌发的影响Fig.2 Effects of light on seed germination of M. boisiana seeds
2.3 温度对金钟藤种子萌发的影响
由图3可知,在28 ℃条件下种子的发芽率为47%,随着温度升高发芽率逐渐降低至24%。32 ℃和34 ℃条件下种子的发芽率显著低于28 ℃ 和30 ℃处理。不同温度处理下种子的发芽势未发现明显差异,但32 ℃和34 ℃处理下的种子发芽指数明显降低。随着温度升高至34 ℃,种子的胚根和胚芽长度显著增加,与28 ℃条件下相比,根长增加50%,芽长增加27%。结果表明,在28~34 ℃范围内,温度升高会降低金钟藤种子的发芽率、发芽势和发芽指数。环境温度达到34 ℃时,明显促进胚根和胚芽的生长。
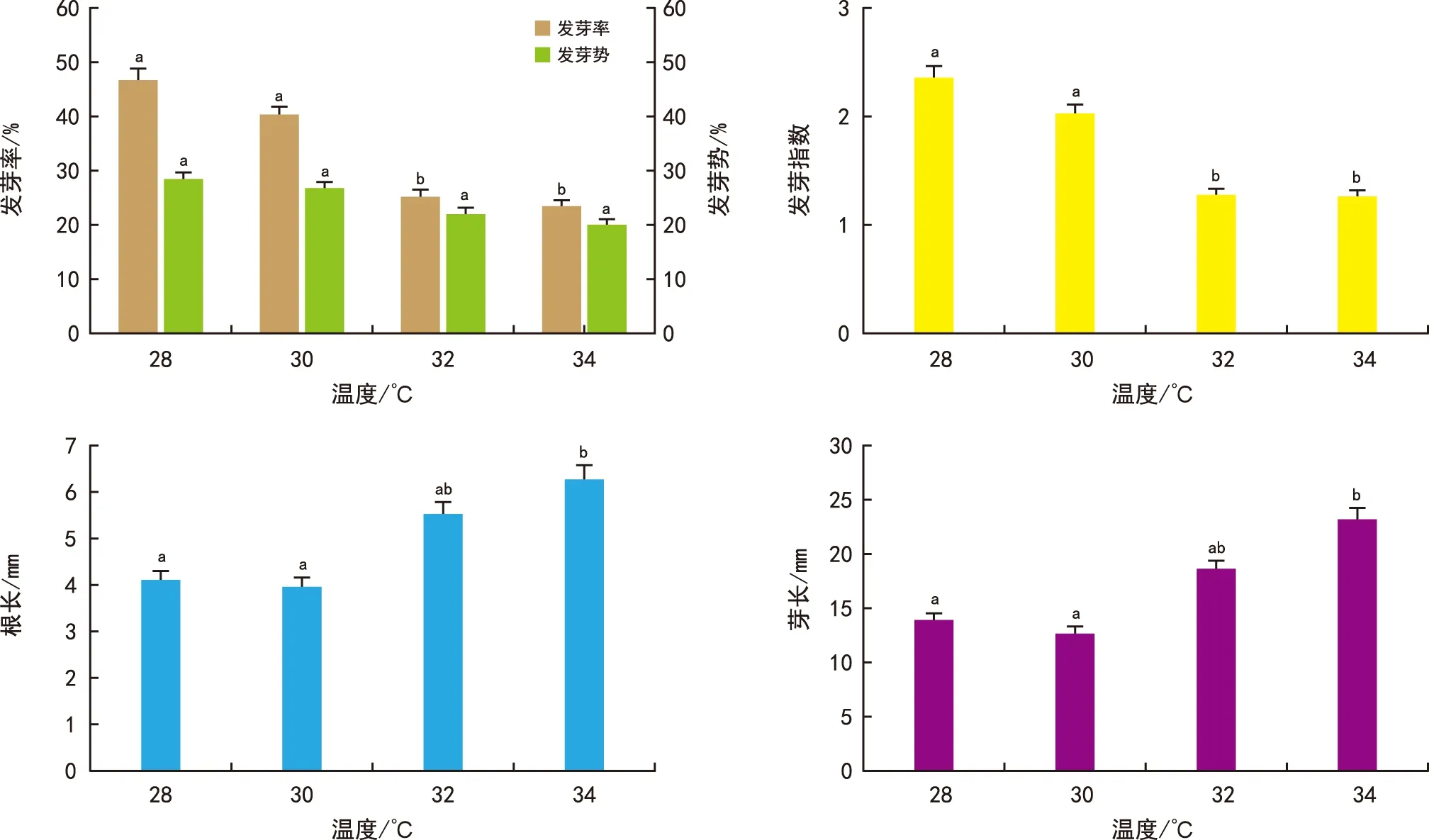
图3 温度对金钟藤种子萌发的影响Fig.3 Effects of temperature on seed germination of M. boisiana seeds
2.4 PEG溶液模拟干旱胁迫对金钟藤种子萌发的影响
由图4可知,PEG溶液使得金钟藤种子脱水,可以模拟干旱环境对种子萌发的影响。当PEG浓度超过10%时,种子萌发率显著降低。15%和20%溶液处理条件下,种子发芽率仅为ck的38.9%。种子发芽势在4种浓度处理下与ck均存在极显著差异,不同浓度之间的发芽势差异不大。15%和20%的PEG溶液显著降低了种子的发芽指数。
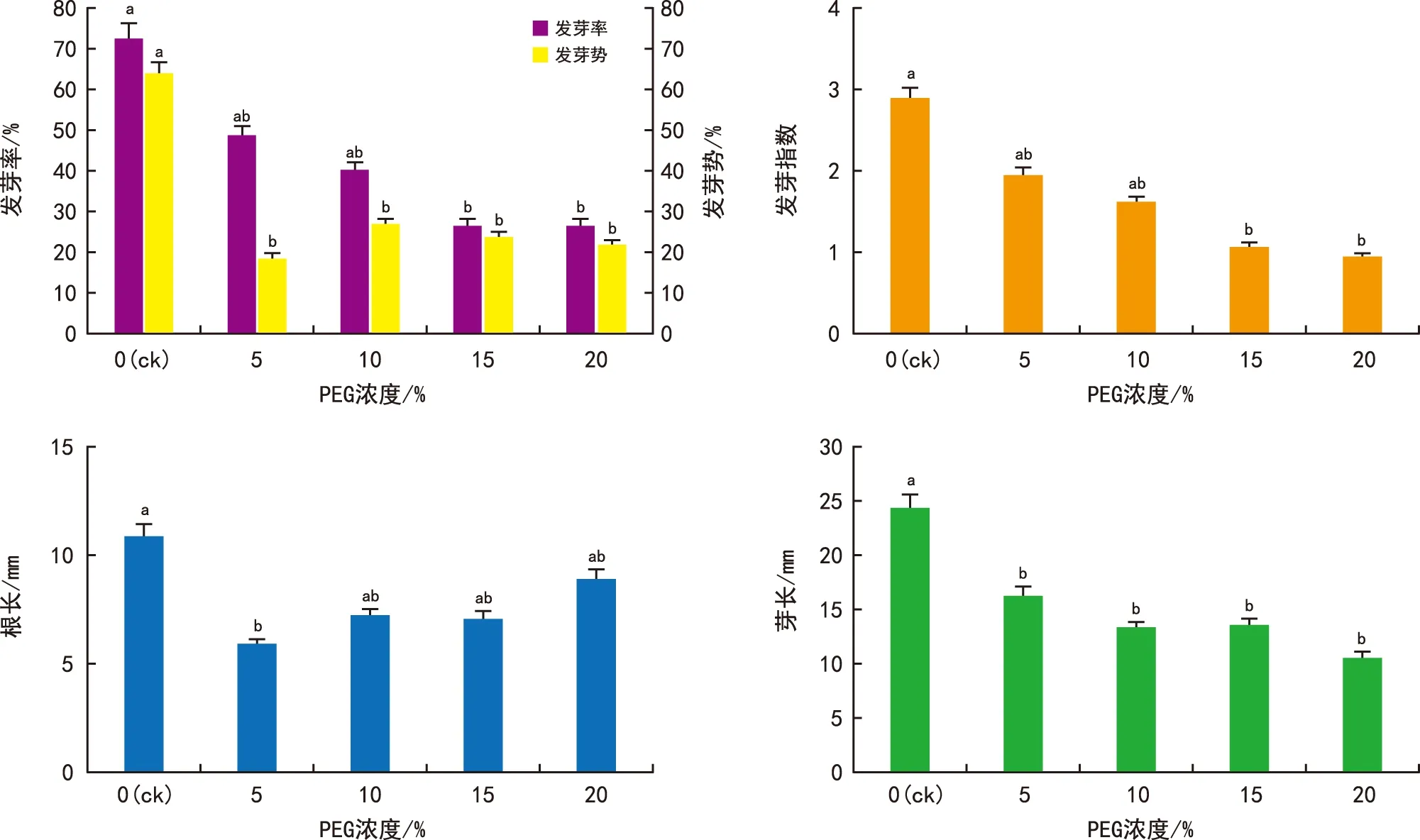
图4 干旱胁迫对金钟藤种子萌发的影响Fig.4 Effects of drought stress on seed germination of M. boisiana seeds
PEG处理使得种子的根长与芽长低于ck。所有处理中,5%PEG浓度组的胚根长度最短。随着PEG浓度的提高,胚根长度逐渐增加,接近但略低于ck的长度。胚芽的长度与PEG浓度呈负相关。
2.5 自化感作用对金钟藤种子萌发的影响
金钟藤组织中含有化感物质,其提取液能抑制其他植物种子的萌发。本组试验研究金钟藤叶片提取液是否会抑制自身种子的萌发。由图5可知,种子的发芽率随着提取液浓度的增加而逐渐降低,浓度为 0.1 g·mL-1的提取液显著降低了种子的发芽率。0.05 g·mL-1和0.1 g·mL-1提取液使得种子的发芽势分别比ck降低28%和22%,差异极显著 (p<0.01) 。提取液抑制了种子胚根的生长,但胚芽长度未受到影响。试验结果表明,金钟藤种子的萌发同样会受到自身化感作用的影响,发芽率和发芽势降低,胚根的生长受到抑制,但胚芽的长度未出现变化。
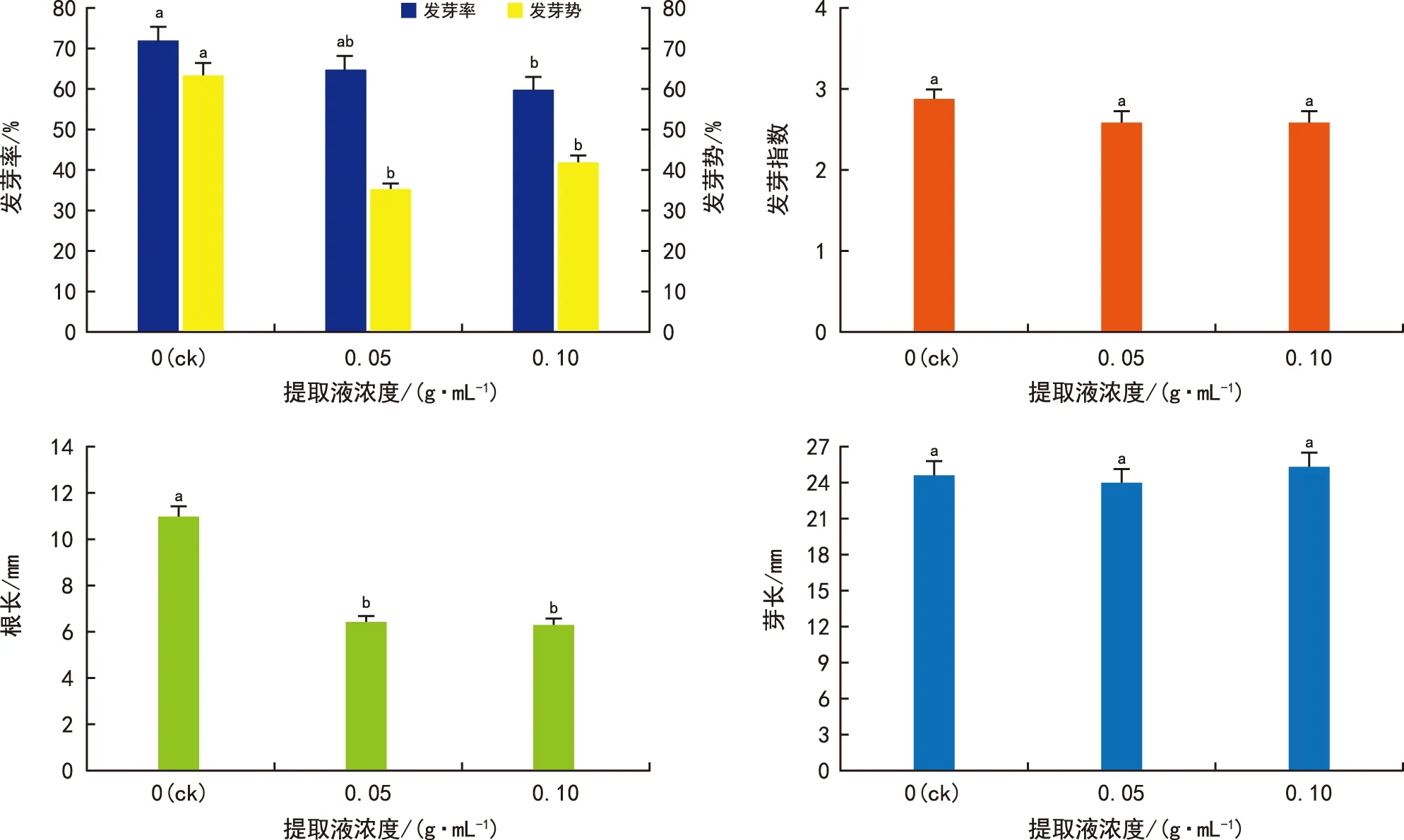
图5 叶片提取液对金钟藤种子萌发的影响Fig.5 Effects of leaf extracts on seed germination of M. boisiana seeds
3 讨 论
3.1 金钟藤种子的硬实性
自然界中部分植物的种子存在硬实现象,种皮坚硬不透水,种子难以吸胀;已经发育成熟的胚无法萌发,出现物理休眠。硬实现象是植物对环境的一种适应机制,可以使种子长期保持活力,直到出现最佳环境条件再萌发生长,对于物种的繁衍具有重要的意义[17]。Li等[18]在调查中发现,野外金钟藤的种子萌发率极低,即使在大量开花的金钟藤分布区域,连续两年都未发现金钟藤幼苗。从本试验结果来看,种皮对金钟藤种子萌发的影响非常显著,对种皮进行物理切损即可大幅提高种子萌发率,试验结果与刘海洋等[13]的结论一致。程汉亭等[16]的研究表明,金钟藤种皮虽然一定程度上阻碍了水分的吸收,但经过112 h的浸泡后仍然可以达到完全吸水的状态。这表明在特定的情况下,比如降水丰富地区或者河流水库周围的区域,金钟藤种子长时间吸水后仍存在萌发生长的可能性。林业管理部门对这些区域应给予关注,加强巡护工作,及早控制金钟藤的种子萌发和扩散。
3.2 温度对金钟藤繁殖的影响
金钟藤是阳性植物,喜光喜热。结合倪广艳等[14]的研究结果和本试验的结论,金钟藤种子萌发的最适温度为28~30 ℃,温度的升高能够进一步促进胚根和胚芽的生长。目前,全球气候出现变暖的趋势,以海南为例,1948—1980年间海南的平均温度为23~25 ℃[19],1980—2010年间平均温度提高到23~27 ℃[20],预计在未来50年平均气温还有可能继续上升1.5 ℃[21],趋近金钟藤种子的最适发芽温度。王伯荪等[79]认为,金钟藤在广州蔓延和为害的重要因素就是广州地区连续几年出现暖冬。因此,未来全球气温的升高可能会造成金钟藤分布区域向高海拔和高纬度地区扩散,同时加快金钟藤生长速度,造成更大的危害。
3.3 金钟藤种子的耐旱能力
干旱条件会使种子失水,显著降低萌发率。金钟藤种子在干旱环境的影响下,发芽率、发芽势和发芽指数都显著降低,但干旱程度的增加促进了胚根的生长,这显示出金钟藤种子对于干旱环境具有较强的适应调节能力。有研究显示,三叶草、大花金鸡菊、桂圆菊和赛菊芋等多种植物在20% PEG-6000胁迫下种子萌发率为零[22-23],但本试验中,同样胁迫条件下金钟藤种子仍保持28%的萌发率。金钟藤植株的耐旱能力也强于与其相同生境下的葛藤[24]。金钟藤种子和植株对干旱环境的较强适应能力是其蔓延扩散的一个因素。
3.4 金钟藤的自化感作用
植物通过合成和释放化感物质,来抑制其他植物的生长,获得竞争优势。曾宋君等[15]和黄乔乔等[25]通过试验证明,金钟藤含有化感物质,其提取液可以抑制菜薹和生菜种子的萌发,但金钟藤种子是否会受到自身化感物质的影响,尚未有明确的结论。本试验发现,金钟藤组织提取液同样会显著抑制自身种子萌发,浓度越高抑制作用越明显。这种现象表明,金钟藤可能通过自化感作用控制自身萌发种子的数量,避免物种内部过度竞争,减少资源消耗。Singh等[26]研究认为,自化感现象是植物适应环境,充分利用资源的一种重要的生态机制。金钟藤和大树杜鹃[27]、菊花[28]一样,能够通过自化感作用来充分利用有限资源,适应环境。
4 结 论
本试验表明,种皮处理、光照、温度、干旱胁迫和自化感作用都会不同程度地影响金钟藤种子的萌发以及幼苗的发育。具体来说,种皮阻隔水分是种子发芽率低的主要因素;光照不会改变种子的发芽率,但会影响种子萌发后幼苗的生长;28 ℃是金钟藤种子萌发的最适温度,温度提高会促进胚根胚芽的生长;干旱会降低种子的发芽率,但促进胚根的生长;化感作用会限制金钟藤自身种子萌发。
虽然目前种子繁殖不是金钟藤泛滥成灾的主要因素,但全球气候变化和特定环境条件可能会提高其有性繁殖能力,扩大分布范围,造成新的危害。对此应加以重视,加强研究和防控工作。
