低温及光照对彩叶杨树叶片中花色素苷变化的影响
2021-07-15朱秀红蔡曜琦杨金橘程红梅茹广欣
朱秀红,蔡曜琦,杨金橘,程红梅,茹广欣
(河南农业大学林学院,郑州 450000)
杨树作为世界上分布最广、适应性最强的树种,生长迅速,干形通直,成材快,轮伐期短,经济与生态效益大。人们对于生态树种的要求不断提高,不同品种的彩叶杨树应运而生。彩叶杨树不仅维持了杨树的独特优势,且均为雄株免于形成飞絮,降低了对于环境的污染。彩叶杨树对于传统杨树病虫害的抗性有所提高,生长迅速,冠型饱满,且不同品种的叶色随季节变化表现丰富多彩,提高了林木观赏价值。彩叶杨树作为改良品种杨树,拥有良好的社会生态效益,填补了杨树品种在彩叶林上的空白。
对于彩叶杨树的研究多为探讨叶色与外界环境条件的生理作用关系,且大部分研究材料为单一品种或2个品种对比,因此进一步探讨研究不同品种彩叶杨树花色苷合成机制及其在不同环境胁迫下对叶片保护作用的调控机理,对进一步丰富以花色素苷呈色为基础的彩叶杨树品种具有重要意义,同时明确彩叶杨树叶片中花色素苷的作用,能对杨树在开发新型彩叶品种方面有进一步突破提供理论指导。研究表明,适当的低温能够促进植物组织花色素苷的积累[1,2]。本试验对5种彩叶杨树进行低温处理,研究低温对彩叶杨树花色素苷含量、叶绿素含量、类胡萝卜素含量、可溶性糖和蔗糖含量的影响,以及相关酶活性变化规律,同时进行遮光处理,探究光照是否参与对其影响,为低温环境下彩叶杨树叶色变化的机制提供依据。
1 材料与方法
1.1 材料与处理
在新密试验田中,选取长势良好的全红杨、中红杨、金红杨、红霞杨与翡翠杨的一年生苗木同侧枝剪下带回进行扦插培养(每个品种每株重复扦插3盆),幼苗生长至25 cm(包括顶芽,主枝3~4片叶)放置于温室内,温度为15~25℃,每天遮光10 h继续培养,确保可取新鲜样品进行重复试验。
试验共设置4个处理:常温常光(CK),温度为25℃/15℃,光照度为200μmol/(m2·s);低温常光(LT):温度为14℃/8℃,光照度为200μmol/(m2·s);低温遮光(LT+D),温度为14℃/8℃,光照度为0 μmol/(m2·s);低温重新通入光源(LT+L),温度为14℃/8℃,光照度为200μmol/(m2·s)。10 d后对5个品种进行取样测定。
1.2 测定方法
采用苏正淑等[3]提出的丙酮乙醇萃取法进行叶绿素与类胡萝卜素含量测定。采用何奕昆等[4]、于晓南[5]的盐酸甲醇分光光度计法进行花色素苷的测定。采用蒽酮比色法[6]测定叶片的可溶性糖及蔗糖含量。花色素苷相关合成酶活性测定采用王惠聪等[7]的方法,略加改动。PAL活性、CHI活性、UFGT活性参照文献[8,9]。DFR活性参照Stafford等[10]在花青苷还原酶转化中采用的方法。
1.3 数据分析
采用Origin 9.0与Excel 2016软件对试验数据进行统计与分析,并用单因素方差分析来检验样本间的差异显著性(置信区间为95%)。
2 结果与分析
2.1 彩叶杨树叶片中叶绿素和类胡萝卜素含量的变化
低温处理后,不同品种彩叶杨树色素含量变化见表1。低温处理下,与对照处理相比,5个品种彩叶杨树的叶绿素a含量均呈下降趋势,其中中红杨、金红杨和红霞杨的叶绿素a含量在1.540~1.653 mg/g。与对照处理相比,全红杨、中红杨、金红杨和红霞杨叶绿素b含量均呈下降趋势,而翡翠杨在低温处理下叶绿素b的含量呈上升趋势,为对照组的2.60倍。低温处理下,5个品种的类胡萝卜素含量下降,其中全红杨类胡萝卜素含量最高,翡翠杨类胡萝卜素含量最低。

表1 低温处理5个品种彩叶杨树的叶绿素及类胡萝卜素含量的变化 (单位:mg/g)
2.2 彩叶杨树叶片中花色素苷含量的变化
如图1所示,单独进行低温胁迫而正常通入光源时,相对低温处理(LT)对5个品种彩叶杨树的花色素苷有明显影响。其中,全红杨的花色素苷含量为常温状态的7.31倍;中红杨在相对低温状态下花色素苷含量为常温状态的7.09倍,变化幅度略低于全红杨;金红杨与红霞杨的花色素苷含量变化幅度基本相同,为常温状态的6.98倍;翡翠杨的花色素苷含量在试验过程中虽然一直保持较低值,但受低温状态的影响其含量上升至常温对照组的5.41倍。由此可知,光照正常时,全红杨叶片内的花色素苷含量易受温度的影响,翡翠杨叶片内的花色素苷受温度影响相对较小。
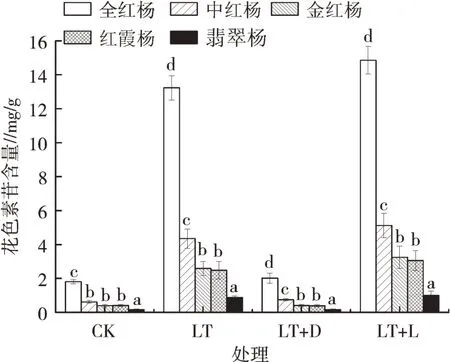
图1 5种彩叶杨树叶片中花色素苷含量的变化
当试验条件变化为相同低温状态进行遮光处理后,与对照组相比,5个品种的花色素苷含量未表现出明显变化,遮光后再次通入光源,5个品种的花色素苷含量超过首次进行低温胁迫时的状态,这表明光照是低温促进彩叶杨树中花色素苷含量的关键条件,当删除这一条件,低温对花色素苷合成与积累的影响较小。
2.3 彩叶杨树叶片中可溶性糖和蔗糖含量的变化
由图2可以看出,与对照相比,相对低温处理(LT)5个品种彩叶杨树的可溶性糖含量及蔗糖含量都呈上升趋势,全红杨的可溶性糖含量为常温状态的4.62倍,蔗糖含量为常温状态的5.01倍;中红杨在相对低温状态下可溶性糖含量上升至常温状态的3.97倍,蔗糖含量上升至常温时的4.09倍;金红杨在相对低温状态下可溶性糖含量上升至常温状态的3.22倍,蔗糖含量上升至常温时的3.29倍;红霞杨在相对低温状态下可溶性糖含量上升至常温状态的2.89倍,蔗糖含量上升至常温时的3.14倍;翡翠杨的可溶性糖含量与蔗糖含量高于金红杨与红霞杨,但变化幅度较低,分别为对照组的1.65倍、1.71倍。
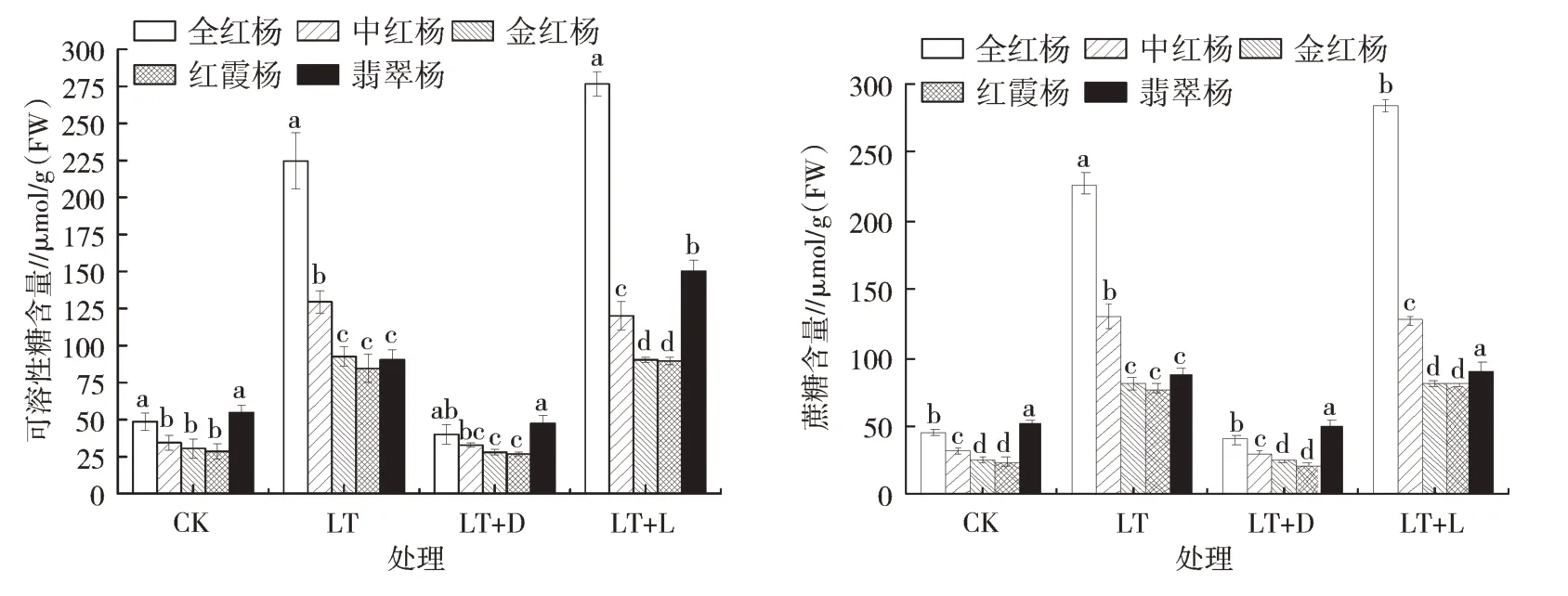
图2 5种彩叶杨树叶片中可溶性糖和蔗糖含量的变化
对彩叶杨树进行遮光处理后,与对照相比,5个品种的可溶性糖及蔗糖含量均有小幅度下降。遮光后再次通入光源,5个品种的可溶性糖及蔗糖含量也大幅度上升,与单独进行低温胁迫持平,其中全红杨与翡翠杨的可溶性糖含量达到4个处理中的最高水平。
2.4 彩叶杨树品种的花色素苷相关酶活性的变化
苯丙氨酸解氨酶(PAL)为花色素苷合成过程中催化苯丙氨酸转化为肉桂酸的关键酶,查耳酮黄烷酮异构酶(CHI)在花色素苷生物合成中将四氢查耳酮环化形成二氢黄酮(即柑橘素)的关键步骤中起催化作用。二氢黄酮醇-4-还原酶(DFR)能够催化3种二氢黄酮醇和2种黄烷酮,生成5种不同的花青素前体,加速花色素苷的生物合成[11]。UFGT作为类黄酮中葡萄糖基的转移酶,将葡萄糖基转移至花色素苷C3的羟基上以稳定花色素苷的结构。这4种酶的功能及活性与花色素苷的合成与积累密切相光。由图3可知,温度下降后,与对照相比,5个品种的4种酶活性均明显上升,与花色素苷含量的变化趋势一致。全红杨的相关酶活性变化最显著,与常温状态相比,低温下PAL活性上升了342%,CHI活性上升了381%,DFR活性上升了390%,UFGT活性上升了420%。中红杨的相关酶活性上升幅度次于全红杨,分别提高了251%、324%、369%和389%。金红杨与红霞杨的相关酶活性上升幅度相似但低于中红杨。翡翠杨的相关酶活性变化幅度相对较低,PAL活性上升了94%,CHI活性上升了147%,DFR活性上升了170%,UFGT活性上升了150%。这表明光照正常时,温度的降低使彩叶杨树中与花色素相关的4种关键酶活性显著上升,花色素苷的合成速率上升,远高于分解速度,进一步造成花色素苷的积累及其他色素含量比例的变化,可能是造成彩叶杨树能够在秋季温度下降后维持叶片色彩,且进行再一次变色的原因。
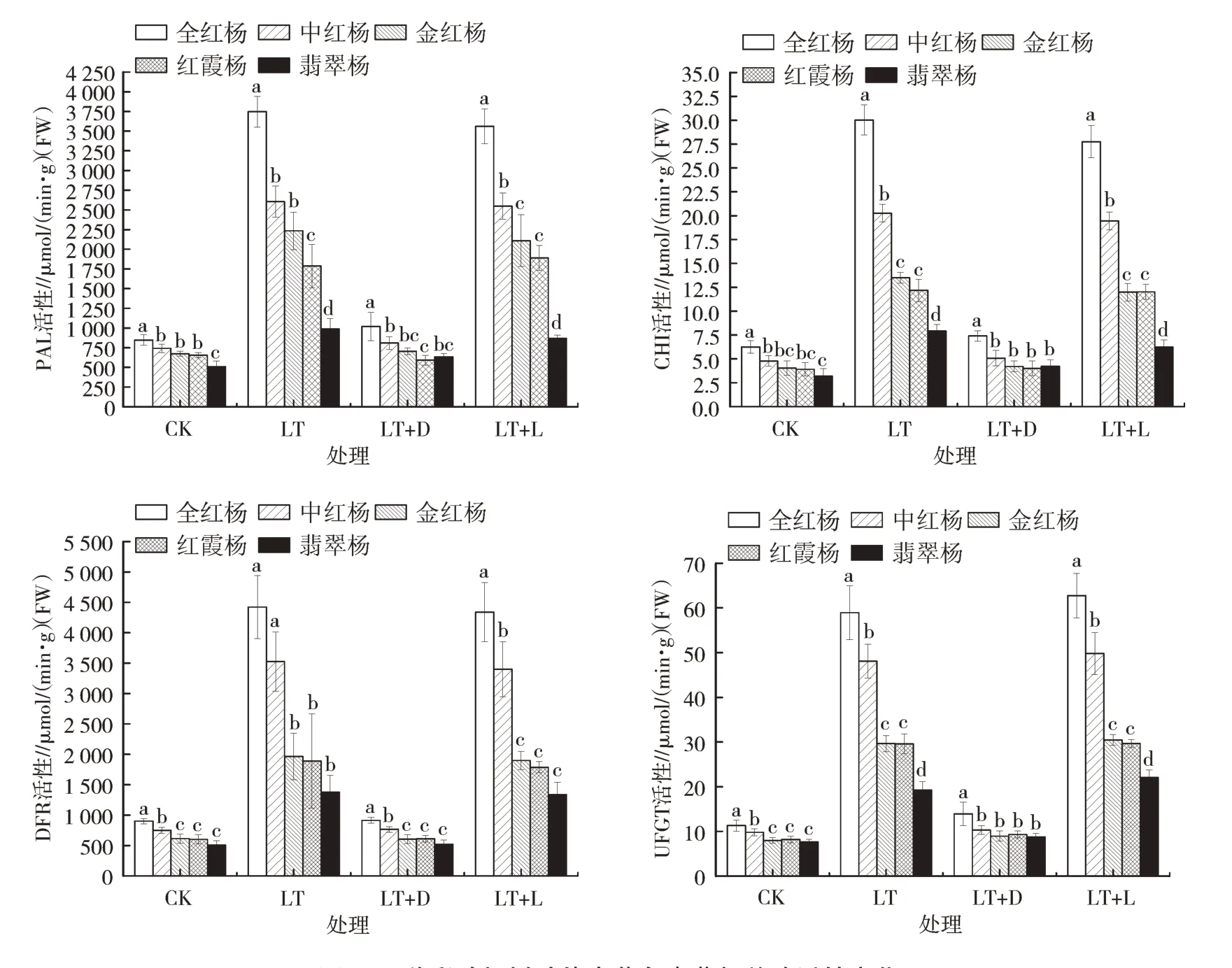
图3 5种彩叶杨树叶片内花色素苷相关酶活性变化
同样地,当没有光照但维持低温环境,5个品种的4种相关酶活性均不再出现大幅度变动,活性维持在常温状态的1倍左右,此时的酶活性与花色素苷含量均较低。低温下再次通入光源后,4种酶活性再次出现大幅度上升。表明温度下降对于彩叶杨树中花色素苷相关酶活性的刺激作用必须有光照作为基本条件,若避免光照而仅降低温度不会引起花色素苷的合成与积累,进而加强花色素苷对于叶片处于胁迫状态时的保护能力。
3 小结与讨论
与极度低温致死环境不同,相对低温环境用来模拟自然生态环境中的秋季低温,以探究彩叶杨树在此时进行变色或维持自身色彩的原因。自然界中的很多彩叶树种,在秋季出现与绿叶树种叶片变黄截然不同的艳丽色彩,有些还能维持进一步的颜色转化,这种鲜明的特点使其具有较高的研究价值。当进行低温处理时,彩叶杨树叶片内叶绿素含量、类胡萝卜素含量呈下降趋势,而花色素苷含量、可溶性糖和蔗糖含量均呈上升趋势,相关酶的活性也不同程度地上升,使得花色素苷不断合成与积累。进行完全暗处理时,彩叶杨树叶片中的花色素苷含量、可溶性糖及蔗糖含量均无明显变化,而重新通入光源后,则呈大幅度的上升,且红叶品种(全红杨、中红杨、红霞杨及金红杨)的变化幅度远大于绿叶品种(翡翠杨)。
叶片光合色素含量是反映植物光合能力的一个重要指标,环境因子的改变会引起叶绿体色素含量的变化,进而引起光合性能的改变[12]。Erdal[13]研究表明,低温导致叶绿素含量下降。一方面是叶绿体色素合成酶活性降低,叶绿体合成受到抑制;另一方面叶绿体结构受损[14]。此外,低温使植物体代谢缓慢,合成叶绿素的原料不足,造成叶绿素含量减少[15]。曾乃燕等[16]对水稻低温胁迫光合色素的影响研究得出,低温使光合色素含量降低。低温胁迫期间,一些木本植物,如降香黄檀、檀香、铁刀木和樟树叶片的叶绿素含量均下降[17]。本研究显示,低温处理下5个品种彩叶杨的叶绿素a和类胡萝卜素的含量都呈下降趋势,可能是低温胁迫导致叶绿素的合成酶活性降低,从而导致叶绿素含量降低。
叶色的转变主要受遗传因素控制,但同时还与环境因子有重要关系[18]。温度是影响彩叶植物叶色变化的重要因素,通常情况下,低温可以增强花色素苷的相关基因表达,当夜间温度越低但不低于致死温度时,造成的昼夜温差越大,越能促进花色素苷的积累。低温还可以通过调整植物组织的比例进而对糖类有机物代谢产生影响,以缓解胁迫环境造成的组织内部水势变化。王小青等[19]研究证明,高温会影响红叶桃叶片的着色,因为过高温度抑制了花色苷合成相关基因CHS、LDOX和MYBIO的表达。蔡建超等[20]研究表明,紫叶稠李在低温胁迫下通过提高色素积累和增加可溶性糖含量,提高自身的耐寒性。刘国君等[21]研究花色素苷含量与苹果抗寒性的关系,发现抗寒性强的品种枝条中花色素苷含量较高。在本试验中,当进行低温处理时,5个品种彩叶杨树叶片内花色素苷含量呈上升趋势,相关酶的活性也不同程度地上升,使得花色素苷不断合成与积累,与前人研究结果一致。
光照是影响彩叶植物叶色变化重要因素,花色素苷合成途径中许多相关酶是光诱导酶,如CHS、PAL、DFR等,其活性均受光照影响。史宝胜等[22]通过对紫叶李的研究发现,弱光处理后的紫叶李重新放入全光照环境下,花色素苷含量显著上升。屈营等[23]通过四季秋海棠的低温胁迫试验进一步证明了低温及光照是花色素苷增加的重要条件。吴飞洋等[24]研究表明,全光照下花色素苷含量显著上升更利于乌桕的变色。因此,相对低温环境的作用并非单一因素,光照也是其充分必要条件。
彩叶杨树栽植容易,属彩叶高大乔木,同时又是速生树种,可迅速提供工程彩叶用树,因此在行道绿化中有较强的发展潜力,是其他树种无法比拟的。本研究通过低温和光照处理,从生理生化变化机制角度对彩叶杨树叶片中花色素苷的影响进行分析,为今后提高彩叶杨树的引种适用性和观赏性应用提供理论依据。
