基于野外调查和同质种植园实验的芦苇植物功能性状变异研究
2021-07-12马香艳刘乐乐尹美淇宋慧佳朱鹏程于晓娜王仁卿郭卫华
马香艳,刘乐乐,尹美淇,宋慧佳,朱鹏程,于晓娜,杜 宁,王仁卿,郭卫华,*
1 山东大学生命科学学院生态学与生物多样性研究所,青岛 266237
2 山东省植被生态工程技术研究中心,青岛 266237
3 山东青岛森林生态系统定位研究站,青岛 266237
自然界中的植物个体表型会受到遗传因素与环境因素的共同影响,从发育和遗传机制来看,种内性状可以分为遗传变异主导的性状和表型可塑性主导的性状两类[1- 2],前者产生的变异是由遗传基因决定且可稳定遗传的,后者产生的变异是由环境直接塑造的,即同一基因型的植物个体在不同的生长环境中表现出不同的表型[3],但是短时间内产生的这种表型变异通常不能被后代继承。虽然这两种机制存在较大的差别,但它们协同共存,都是生物适应异质环境的重要手段[4]。在全球气候变化背景下,植物对多重环境变化的响应及适应机制成为生态学研究的热点[5- 6],植物表型变异及其内部机制的研究不仅有助于解析植物对环境的适应策略,还能对物种未来分布和进化的预测模型提供理论依据。
植物功能性状是植物应对环境变化形成的一系列形态、生理、物候等特征属性,这些属性通常是在植物中比较常见的表型,可以直接或者间接地反映生存环境的变化[7- 8]。比如,随着气温升高,植物叶片的氮磷含量下降,氮磷比呈现上升趋势[9];与湿润生境相比,处于干旱生境中的植物叶片单位面积叶干重较大,叶片气孔导度较低,有利于植物提高水分利用效率[10]。同时,植物功能性状也受谱系影响,研究者对中国东部森林中48个植物物种的出叶时间、叶片大小、叶片单位面积质量等叶片性状间的相关性进行研究时,发现了其性状受到了物种系统发育差异的强烈影响[11],遗传因素在植物生长的整个过程中也发挥着重要的作用。此外,当环境发生变化时,植物性状的改变往往不是单一的,而是多个植物功能性状协同变化的,性状间存在复杂的协同和权衡关系,共同维持和调节植物的生命活动[4, 12],植物的叶片氮含量、比叶面积等与资源获取相关的叶性状之间联系紧密,叶片氮含量升高,往往伴随着比叶面积的升高[13]。植物功能性状既相互联系又存在较大的种内与种间差异,普定喀斯特地区的木本植物相比于同纬度其他生境植物来说,有着较小的叶面积和较大的叶组织密度等性状特征,这些有利于减少蒸腾和增加养分储存的性状组合可能是其适应干旱贫瘠的喀斯特环境而采取的生态策略[14]。植物功能性状变异性及性状相关性的研究对于理解植物应对全球变化、环境监测与评价、生态保护和恢复等理论研究和实践活动具有重要意义[15]。
广泛分布的物种所处的环境多样,往往具有大量的表型变异,由于局部适应还会形成稳定的种内分化[16]。芦苇(Phragmitesaustralis)作为一种世界广布的湿地植物,主要通过风媒异交的种子进行有性生殖,也可以通过根状茎进行营养繁殖,具有非常高的种内遗传多样性和表型可塑性,而且其生态适应幅度很宽,可以生长在内陆湖泊、河口和滨海滩涂等浅水湿地,也可以生长在盐碱、荒漠等恶劣的环境中[17- 19]。芦苇生境的多样性导致了其丰富的生态型,在中国河西走廊地区根据生境不同将芦苇划分为沙丘芦苇、沼泽芦苇、盐化草甸芦苇等[20],在黄河下游根据芦苇种群形态的变异规律,将该地区芦苇划分为盐生芦苇、淡水芦苇、巨型芦苇3个形态类型[21]。在全球尺度上,芦苇的株高和单株最大生物量随着纬度的增加而减少,而这两个性状的表型可塑性随着纬度的增加而增加,这种性状和表型可塑性变异都是对环境适应的结果[22]。然而,现在对中国芦苇的研究只局限于小区域,缺少对中国芦苇在较大尺度上的对比研究,这限制了对中国芦苇表型分化与生态适应的深入认识。黄河上游的宁夏平原和黄河下游的黄河三角洲都处于黄河流域,具有连通性,同时跨越中国西部和东部不同的气候区域,又具有气候差异性,这为我们在更大尺度上的研究提供了良好的地理条件。同时,芦苇作为一种世界广布的植物,其高度的表型变异和丰富的生态型为我们研究植物种内表型差异提供了良好的实验材料。目前已有较多研究探讨了芦苇适应异质生境的策略[23],但是从表型可塑性和遗传变异两方面阐明其适应机制的报道却相对较少。
本实验以芦苇为研究对象,将野外调查测量实验和同质园移栽实验结合起来,比较分析黄河上游宁夏平原和黄河下游黄河三角洲两个区域芦苇的植物功能性状差异,揭示芦苇在不同气候区域的表型分化和不同性状之间的潜在联系,加深对芦苇的适应性进化机制的认识。
1 研究方法
1.1 野外调查和采样
宁夏平原位于黄河上游(35.83°N—39.38°N,104.28°E—107.20°E),处于中国西部内陆地区,为典型的温带大陆性气候,昼夜温差和气温年较差大,7月气温最高,平均为20.0—23.1℃,1月气温最低,平均为-8.2—-6.6℃,平均年降水量为262.3—363.2mm,干旱少雨,生态环境脆弱[24- 25]。黄河三角洲位于黄河下游(36.55°N—38.12°N,118.07°E—119.18°E),处于中国东部临海区域,具有中国暖温带最广阔、最完整的新生河口湿地,为温带大陆季风性气候,年平均气温约为12.1℃,年平均降水量约为551.6mm,四季分明,光照充足,雨热同期[26],而且水盐生境异质性强,具有许多的芦苇单优群落[21]。
2018年7月下旬和8月上旬,分别在黄河三角洲和宁夏平原两个区域各调查30个芦苇种群,种群之间距离大于10km,每个种群内选取5棵健康植株,个体间相距约20m,即在宁夏平原和黄河三角洲各选取150株植株个体进行植物功能性状的测定。所测的功能性状和测量方法如下,植株地面位置到植株最大高度的距离为株高;靠近地面或水面的芦苇节间的直径为基径;每个区域中抽穗的芦苇种群数量除以总的种群数量为抽穗占比。在所选植株上随机选取3片完全展开的、生长健康的叶片,叶片最后端到叶尖间的距离记为叶长;叶片最宽处的距离为叶宽;测完叶长、叶宽后将3片叶子叠在一起,对折两次后测量叶片平均厚度为叶厚。通过扫描、图像处理程序ImageJ v1.8.0确定叶面积,然后将叶片放入80℃烘箱中48h烘至恒重得到叶干重,计算得出芦苇叶片的比叶面积(Special leaf area,SLA)。最后沿着地面或水面收获所测植株的地上部分,烘至恒重得到植株干重,再加上之前测得的3片叶子的叶干重即为芦苇整株地上生物量。根据野外测量地点的情况,收集部分种群中所测芦苇的根状茎,将其带回实验室用于后续的同质种植园试验。
1.2 同质种植园实验
芦苇同质园位于山东省青岛市山东大学青岛校区(36.36°N,120.69°E),属于典型的温带季风性气候,年平均气温为13.2℃,年平均降水量为666.7mm,夏季高温多雨,冬季寒冷干燥[27]。我们共采集了宁夏平原18个种群的根状茎(每个种群选取1个个体),黄河三角洲16个种群的根状茎(每个种群选取1个个体),于2018年8月,将这些芦苇根状茎分别种植在内径约25cm、高约33cm、容量约16L的塑料桶中并放置于温室。于2019年5月,将所有发芽芦苇转移到露天同质种植园中,桶间距为1m,确保不同种群芦苇间彼此不受影响。在芦苇生长期间,定期对芦苇进行除虫、施肥、人工除草,定期浇水,保持水分等资源充足。
2019年9月,将每桶芦苇作为统计单位,从每桶芦苇中选取5棵健康植株,用与野外相同的方法测得同质园芦苇的株高、基径、叶长、叶宽、叶厚、比叶面积等性状,并观察数出每桶中芦苇的株数。同时,分别记录桶中芦苇是否抽穗,用每个区域中抽穗的芦苇种群数量除以每个区域的总种群数量得到抽穗占比。除了以上形态性状外,在同质园中还测量了叶片气体交换、叶绿素荧光、叶片含氮量、叶片含磷量等生理指标。在晴朗无云的天气条件下,在早上9点到11点,使用Li- 6800便携式光合仪(LI-COR,美国)进行测量,设置其光强为1800 μmol m-2s-1,二氧化碳浓度为400 μmol/mol,温度为25℃,湿度为50%,空气流速为400 μmol/s,测得芦苇的净光合速率、蒸腾速率、气孔导度和胞间二氧化碳浓度。同时,使用PAM- 2500便携式调制叶绿素荧光仪(WALZ,德国)对植物叶片测定瞬时荧光值(Ft)和最大荧光值(Fm’),然后通过计算得到PSII的实际量子效率、非光化学淬灭、电子传递效率、PSII的最大量子效率等荧光参数。光合和荧光指标测量完毕后,收获同质园中芦苇的地上部分,烘干测定每桶芦苇地上生物量。新鲜叶片的全氮含量使用凯氏定氮法获得。植物样品经过浓硫酸消解后,用钼锑抗比色法测定总磷含量。
1.3 统计分析
测量的数据使用Microsoft Excel 2016进行整理,使用IBM SPSS Statistics V21.0数据分析软件对所测的性状数据进行t检验和Duncan检验,以比较在野外和同质园环境下的宁夏平原和黄河三角洲芦苇性状的差异。为检验性状的可遗传性与可塑性,对同质园中宁夏平原芦苇的株高、基径、叶长、叶宽、叶厚、比叶面积等性状分别与相对应的野外种群性状进行Pearson相关性和线性回归分析。为检验性状之间的协同与权衡关系,对所测的主要性状进行Spearman相关性分析。绘图工具采用GraphPad Prism 8.4.2和R软件。
2 结果
2.1 不同种源间芦苇植物功能性状比较
在野外环境中,来自宁夏平原和黄河三角洲的芦苇在株高、基径、叶长、叶宽、叶厚性状上存在显著差异(图1;表1);在同质园环境下,来自两个地区的芦苇在基径、叶长、叶宽、叶厚性状上也存在显著差异(图1;表2)。来自宁夏平原的芦苇无论是在野外还是同质园环境下,其基径、叶长、叶宽都显著低于黄河三角洲芦苇(图1),但宁夏平原芦苇的抽穗占比要高于黄河三角洲芦苇(表1;表2)。在野外环境中,宁夏平原芦苇的株高、叶厚显著高于黄河三角洲芦苇(图1;表1)。在同质园环境中,两个种源间的芦苇在光合、呼吸等生理性状上均无差异(表3),但在叶片化学元素氮、磷含量和植株密度方面存在显著差异,来自宁夏平原的芦苇叶片含氮量和含磷量较低(表2),在植株数量(密度)上显著多于黄河三角洲芦苇(表2)。

表1 野外宁夏平原和黄河三角洲芦苇的植物功能性状比较

表2 同质园中宁夏平原和黄河三角洲芦苇的形态及化学元素含量性状比较
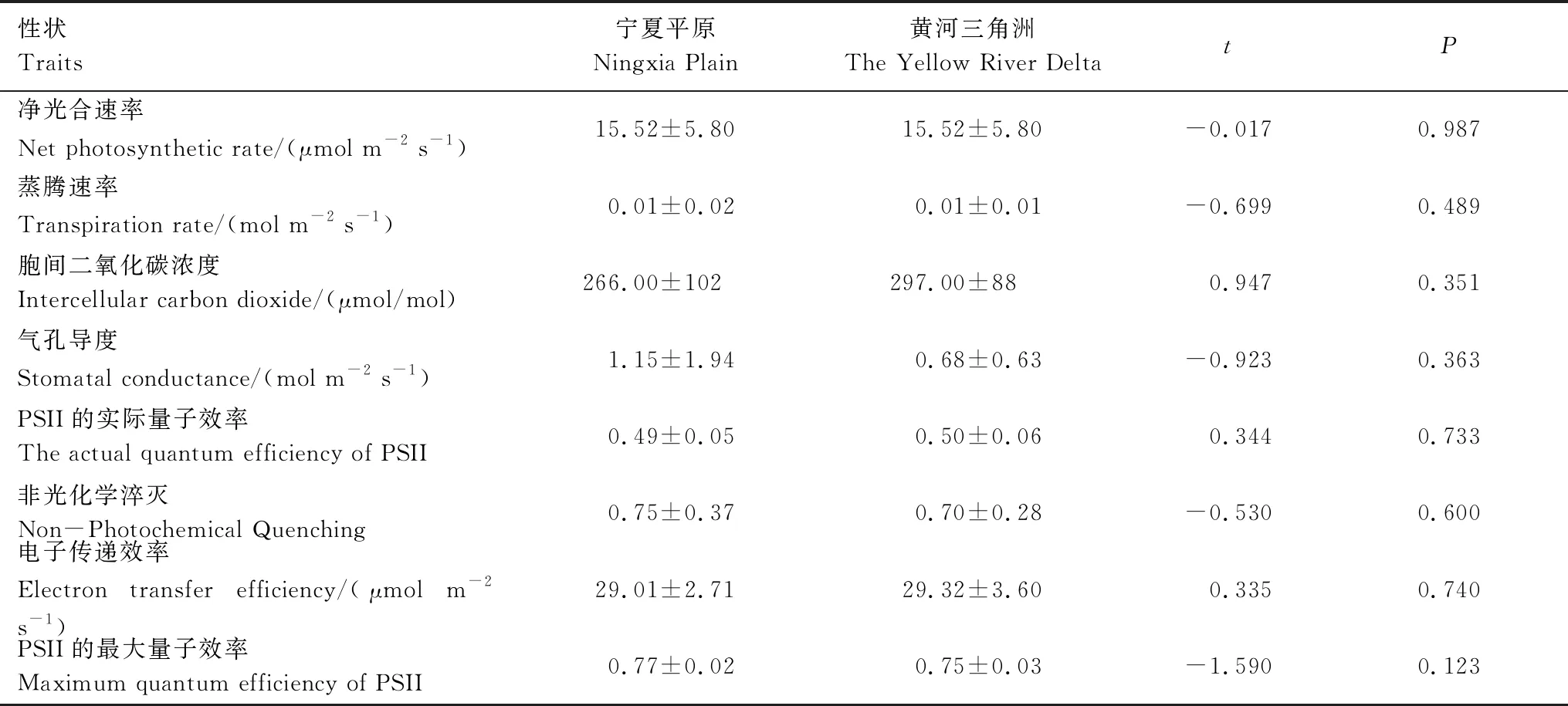
表3 同质园中宁夏平原和黄河三角洲芦苇的生理性状比较
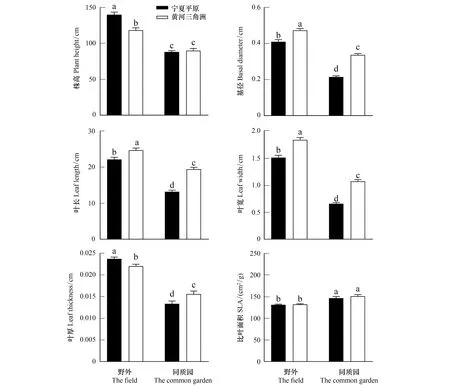
图1 宁夏平原和黄河三角洲芦苇在野外和同质园环境中植物功能性状的比较(数值为平均值±标准误差)
2.2 芦苇植物功能性状间的关联分析
芦苇的各个植物功能性状之间具有不同的关联,研究结果表明,无论是野外还是同质园环境中,比叶面积和基径、叶厚性状间都存在显著负相关关系(图2)。大部分芦苇都表现出叶长和叶宽两个性状的显著正相关,然而大多数芦苇的株高与基径、叶长、叶宽等性状间无显著相关性(图2)。

图2 芦苇植物功能性状间Spearman相关性分析
2.3 野外和同质园中芦苇植物功能性状相关性分析
宁夏平原芦苇在野外和同质园两种环境下表现出了不同的表型,对于株高、叶长两个性状来说,两个环境下生长的芦苇之间存在显著正相关关系(表4),而在基径、叶宽、叶厚、比叶面积等性状上不存在显著的相关关系(表4)。
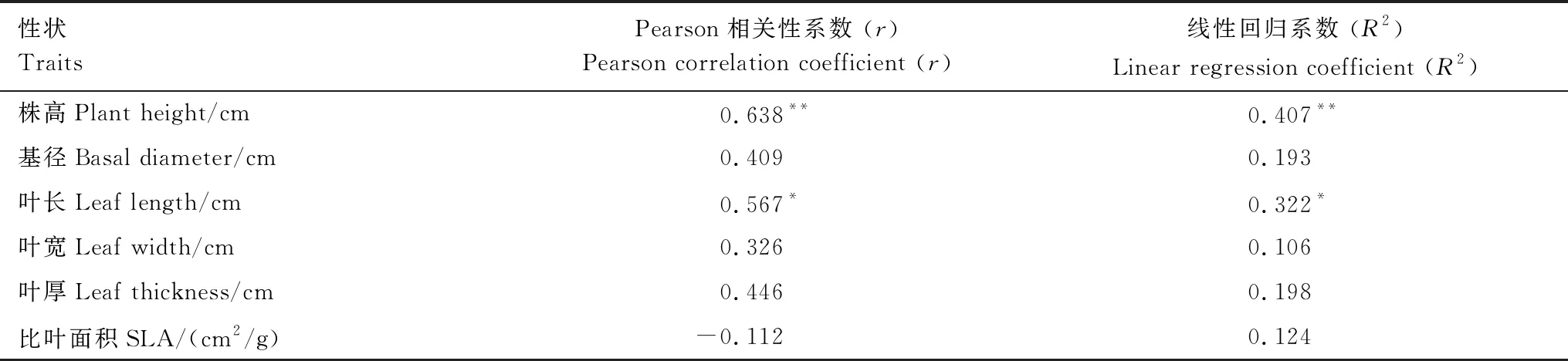
表4 宁夏平原芦苇在同质园和野外的植物功能性状Pearson相关性和线性回归分析
3 讨论与分析
来源于不同环境的植物在株高、基径、叶片性状等植物功能性状上会产生差异,这是植物为适应不同环境
表格第2、3列为性状平均值±标准差,表格第4、5列为t检验结果而采取的生存策略[4]。来自宁夏平原和黄河三角洲的芦苇在野外和同质园中都在株高、基径、叶长、叶宽、叶厚等形态性状方面存在显著性差异,在叶片含氮量和含磷量等化学成分含量等性状上也有显著不同,但是在叶片气体交换、叶绿素荧光等生理性状和比叶面积性状方面并无显著差异。芦苇性状间也存在协变现象,通过性状间相互权衡、相互影响来使植物达到资源利用最优化[28],维持其正常的生长。当生存环境发生很大的变化时,宁夏平原芦苇的株高和叶长还是主要受到遗传因素的影响,基径、叶宽、叶厚、比叶面积等性状受到环境的影响较大,遗传和环境共同影响着植物的功能性状。在未来全球变化的大背景下,植物可能会不断通过遗传变异和表型可塑性两种机制在新环境中生存繁殖和定居[3]。
在不同的气候等环境条件下,芦苇种群会发生本地适应,产生稳定的表型分化。中国东西部不同来源植物间的稳定表型分化的形成可能和降水有关,对内蒙古草原与荒漠区中4种一年生植物进行模拟研究时就发现,随着有效降水梯度的增加,植物的生长期、植物高度、叶长、生物量等均随之增加[29]。在本研究中,黄河三角洲靠近海岸,年平均降水量为551.6mm,降水丰沛,地质复杂且异质性强[26],而宁夏平原处于内陆地区,年平均降水量为262.3—363.2mm,降水不足,气候干燥[25],在正常年份,黄河三角洲年平均降水量约为宁夏平原的两倍,水分可能成为限制宁夏平原芦苇叶片性状的主要因素之一。对于草本植物来说,在抽穗、开花期,降水量越多,物候发生时间越提前[30]。但是有的研究却发现,土壤水分的减少可能导致植物快速完成营养生长而提前进入生殖生长阶段,长期干旱造成了植物物候的提前[29, 31]。在同质园中,我们发现宁夏平原芦苇抽穗时间早于黄河三角洲芦苇,宁夏平原芦苇抽穗比例也大于黄河三角洲芦苇(表2),这与在野外观察到的宁夏平原芦苇抽穗植株占比远远大于黄河三角洲芦苇相一致(表1),长期对不同水分条件的适应形成了两个地区芦苇由遗传控制的稳定表型分化。在本研究中,野外宁夏平原芦苇的株高显著高于黄河三角洲芦苇(图1),可能是由于黄河三角洲盐分含量过高,抑制了芦苇的生长,这与在加盐处理的控制实验中得到的结果相一致,加盐处理下,河口型和内陆型两种生态型芦苇的株高都呈现出下降趋势,都遭受到了盐胁迫,株高下降也是芦苇针对高盐生境的一种表型适应策略[32]。两个地区芦苇的株高在野外条件下也存在显著差异,但是在同质园环境下,这种株高差异消失(图1),表明了环境对植物有着很大的影响,表型可塑性也是芦苇产生表型分化以适应环境变化的重要机制。因此,芦苇在不同气候环境下的植物功能性状变异受遗传变异和表型可塑性共同影响。
植物的叶片性状与植物利用资源的能力密切相关,而且也能反映出植物应对不同环境的生存策略[33]。其中,比叶面积是最重要的性状之一,能反映植物对不同生境的适应特征,使其成为植物功能性状研究中的重要指标[34]。许多研究认为,在资源丰富的环境中生长的物种往往有较高的比叶面积,而较低的比叶面积和大的叶片厚度有利于植物适应资源匮乏的环境[35]。在本研究中,同质园中的芦苇比叶面积要显著高于野外环境中的芦苇(图1),因为在同质园中,芦苇生长环境相对优越,与野外生境相比,水热光资源较为充裕,较大的比叶面积有利于其充分利用光照,快速获取资源。在对木本植物的研究中也证明了这点,研究表明,来自湿润环境的栎树具有高的比叶面积和低的叶片厚度,同时具有较高的光合速率,但生长速率因时期而异,是一种提高竞争能力的资源获取策略[36]。黄河三角洲芦苇的叶长、叶宽等叶片性状都显著大于宁夏平原,叶片中的氮磷含量也较高,但是黄河三角洲芦苇的密度却远远小于宁夏平原芦苇(表2),这反映了植物适应环境的不同策略,黄河三角洲芦苇偏向于对资源快速获取的方式来适应环境,而宁夏平原芦苇则通过增加株数、提高繁殖力来维持生长,都是植物长期适应环境形成的结果[37]。
植物对环境的适应并不是通过单一性状的改变而实现的,植物的各个性状之间是相互关联、相互影响的[28]。现在研究的对象也从植物叶片拓展到了根、茎、花、种子等其他器官及植物整体性状[12],植物的不同器官(如叶、茎和花)对资源环境变化的响应往往是相互作用的,以便其进行有效的生长和繁殖,例如在武夷山59种木本植物的树皮、茎干、叶片功能性状特征及关联研究中发现,树皮厚度与茎干含水率、茎干氮含量呈极显著负相关,树皮碳含量与茎干碳含量呈显著正相关[38],功能性状间的关联性有利于植物快速调整生存策略以适应环境。在本研究中,比叶面积和基径两个植物性状是协同变化的,两者间呈显著负相关关系(图2),即植物比叶面积越大,基径越小,芦苇通过比叶面积和基径两个性状间的协变来共同适应环境,体现性状间的相互权衡。这种相关性不仅体现在植物不同器官之间,同一个器官内也具有很强的协变性[39],例如叶片形态大小和叶片表面温度间存在某种关联,在高温干旱区域,小的叶片利于降低叶片表面温度,这是植物对特殊生境的一种适应策略[40]。这种性状协变性的大小还可能会限制表型可塑性,有研究表明不同的环境条件会改变或导致性状协变性的变化,从而使表型可塑性得到改变,但是在研究中这种协变性相对较为稳定,所以对表型可塑性影响较小[41]。
同种植物的植物功能性状变异不仅受到表型可塑性的影响,也受到遗传因素的控制,二者可以同时作为某些植物耐受性强、生态型丰富的重要原因。当植物的生存条件突然改变,植物的表型可塑性导致其产生与原来不同的功能性状来适应新环境,例如本研究中芦苇在同质园环境中比叶面积大大增加。这种机制能使植物快速的改变生存策略,在最短的时间内获得最高的资源利用效率。表型可塑性也可能是物种避免灭绝或迁徙的一种策略,通过增强个体的适应能力来扩大其对多种环境更广泛的耐受性[22]。植物如果长期生存在某一特定环境中,遗传基因可能发生改变,导致植物表型产生稳定的变异,这是植物长期适应其生存环境的结果,就像本研究中宁夏平原芦苇的株高和叶长受到遗传因素的影响,当环境改变时仍然保持相同的生长策略,在野外和同质园环境下都具有显著相关性。当然,在全球气候变化的大背景下,更多不同来源的芦苇对多重环境因子的响应机制还需要得到持续的关注和探究[42]。
本论文探究了黄河上下游芦苇在野外和同质种植园中的植物功能性状变异规律,揭示了遗传分化和表型可塑性在芦苇环境适应中的重要作用,对进一步研究广布植物物种对全球环境变化的响应和适应机制具有重要意义,也为芦苇湿地的保护、恢复和管理提供了科学依据。
