女贞种质资源ISSR分析与抗寒优株选育
2021-07-09姜楠南刘凤栾杨传祥房义福
姜楠南,孙 音,刘凤栾,王 媛,卢 洁,杨传祥,房义福
(1.山东省林业科学研究院 园林与花卉所,山东 济南 250014;2.上海辰山植物园中国科学院上海辰山植物科学研究中心,上海 201602;3.山东省林木种苗和花卉站,山东 济南 250014;4.泰禾园林绿化有限公司,山东 日照 276800)
女贞Ligustrum lucidum为木犀科Oleaceae 女贞属Ligustrum常绿阔叶小乔木,其树形优美、叶片光亮、果实累累,具有良好的园林应用价值和较高的药用价值[1]。女贞原产于我国长江流域及其以南的华南、西南地区,分布广泛,西北至陕西、甘肃,东到江苏,朝鲜也有分布[2]。女贞适应性强,对低温低湿环境及铅、铬、锌、二氧化硫等污染具有较强的耐受性[3]。由于其叶片大且光亮、经冬不落,近年来被大量引种到我国黄河中下游地区,是山东、河南、河北等黄河中下游地区为数不多的常绿阔叶乔木树种,在隆冬时节其观赏性状突出。但是引入山东及其周边地区后易受冻害,表现为冬末全部或大部分叶片枯黄脱落并保持落叶状态,直至来年新叶萌生,甚至1年生枝条冻死或地上枝干全部冻死等。因而选育适应北方地区气候的抗寒女贞优株,对于华北地区的生态建设和绿化美化具有十分重要的意义。
近年来,关于女贞抗寒性鉴定方法及生理生化指标测定的研究多有报道。林艳等[4]、李忠喜等[5]以相对电导率为指标筛选抗寒女贞;郝明灼等[6]对4 个女贞品种盆栽苗抽梢展叶期的抗寒性进行研究,利用Logistic 方程拟合得出女贞的低温半致死温度为-5.69 ℃。植物在不同物候期的抗寒能力存在较为明显的差异[7]。杨晓玲等[8]对3 种女贞的叶片抗寒、抗旱生理指标进行了研究,认为女贞的抗低温和抗干旱能力较强。林艳等[9]经研究发现,相对较高的温度和湿度可降低女贞主干抽梢率,增强女贞的抗寒性能。上述文献均以电导率为指标研究了不同品种、不同树龄、不同环境条件下女贞的抗寒性。抗寒性是植物在对低温的长期适应中通过本身的变异和自然选择获得的一种遗传特性。女贞的抗寒性由其在低温胁迫下的生理调节能力决定,生理调节能力与植株的遗传特性、解剖结构、环境及当年生长发育情况等有关[10]。前期研究结果表明,部分抗寒女贞优株的抗寒表现不稳定,可塑性较大。目前,对女贞抗寒稳定性进行多年连续观测的研究报道较少,将抗寒表型与生理指标、解剖、遗传相结合的研究鲜见报道。
本课题组连续多年观察筛选引入的女贞,淘汰冻死冻伤女贞种质,初步筛选出抗寒表现较好的60 份女贞种质。由于女贞抗寒表现具有不稳定性,为了从中筛选出抗寒性强且稳定的女贞种质,课题组对初选的60 份女贞种质,在连续调查其露地越冬表现的同时,采集具有不同抗寒表现的叶片观察叶脉和叶肉的解剖结构,采集1年生枝条测定电导率,计算胁迫条件下的低温半致死温度,综合评价各女贞种质的抗寒能力。采用ISSR(inter-simple sequence repeats)分子标记的方式,对60 份女贞种质进行遗传多样性分析,将女贞种质的聚类结果和抗寒能力进行比较分析。
1 材料与方法
1.1 试验地概况
试验地位于山东省济南市长清区(116°63′E,36°38′N)。2016年,平均气温13.8 ℃,年平均降水量623.1 mm,无霜期186 d,1月平均气温-3.2 ℃,极端最低气温-16 ℃(1月20日)。
1.2 供试材料
从山东、河南、河北、四川等地栽植的女贞中收集抗寒种质资源共185 份,2010年进行嫁接繁殖,2011年定植在长清区试验地。2017年,选择不同来源且具有代表性的60 份材料作为试验材料(表1)。其中,D5、D6、D7 为B3 的实生苗后代。
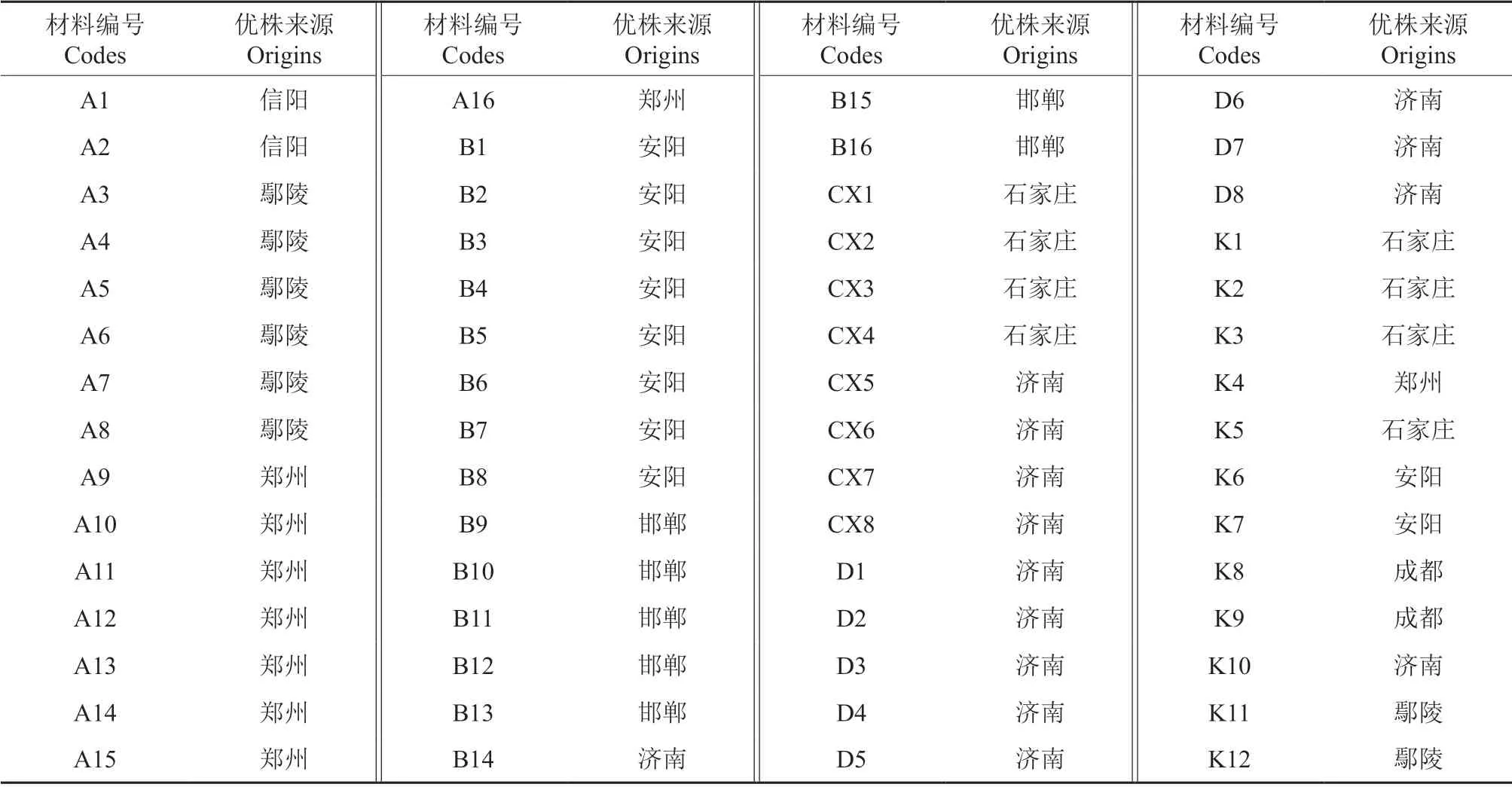
表1 60 份女贞种质材料及其来源Table 1 60 germplasm materials of L.lucidum and their origins
1.3 试验方法
1.3.1 女贞抗寒表型观测
2011—2017年的每年3月(女贞换叶期),调查女贞露地越冬表现,统计主干、枝条受冻情况和叶片枯落情况。
1.3.2 叶片解剖结构观察
2017年1月,采集各女贞种质材料的叶片,采用石蜡切片法[11]制作切片,切片厚度8 μm,番红-固绿染色,使用奥林巴斯B50 显微镜观察并拍照。比较叶片主叶脉、栅栏组织和海绵组织等的差异,分析其与抗寒能力的相关性。
1.3.3 相对电导率测定及低温半致死温度计算
2017年11月初,采集各女贞种质材料相对饱满健壮的1年生休眠枝条,剪成10 cm 长度,随机分成5 份,装入自封袋中,于4 ℃条件下平衡24 h,然后分别置于0、-5、-10、-15 和-20 ℃条件下处理12 h。每份样品避开两端及芽眼,剪成约0.3 cm 长度枝段,室温下浸提12 h 后,测定初电导值(S1),煮沸30 min,冷却至室温后,测定终电导值(S2)。室温下测定去离子水电导值(S0)[12],计算相对电导率(SR)。
SR= [(S1-S0)/(S2-S0)]×100%。
古人说,心安茅屋稳,性定菜根香,心安即是福。是的,抬腿之劳,一个小小过失的弥补,便能让人心安,便能让人感念生活,感悟人生,便能让世界充满爱,于人于己都是满满的正能量,何乐而不为呢?!
使用SPSS 软件拟合Logistic 回归方程
y=k/(1+ae-bt)。
式中:y为相对电导率,x为处理温度,k为常数。计算相对电导率的拐点温度lna/b,即低温半致死温度[13]。
1.3.4 ISSR 分子标记及聚类
采用改良的CTAB 法[14]抽提各女贞种质材料的总基因组DNA,采用ISSR 分子标记的方法,使用优化的ISSR-PCR 反应体系[15],对96 条哥伦比亚大学开发的通用引物进行筛选用于PCR 扩增。使用POPGENE 软件计算多态位点百分率。
将电泳图谱同一位置清晰出现的条带记为“1”,无条带记为“0”,将条带信息转换成“0”和“1”组成的原始矩阵,使用NTSYSpc-2.11F 软件进行数据分析。对原始矩阵用SimQual 程序求DICE 相似系数矩阵,并获得相似系数矩阵。依据Nei’s 遗传距离,采用SHAN 程序中的UPGMA 方法进行聚类分析,通过Tree plot 模块生成聚类图,构建各种质材料间的亲缘关系。
2 结果与分析
2.1 各女贞种质材料的抗寒表型分级
2014—2017年,每年11月15日—次年3月15日长清区日最低温度变化如图1所示。
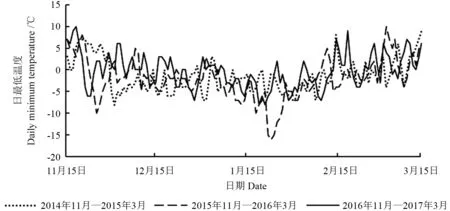
图1 2014—2017年女贞越冬期间长清区日最低温度的变化Fig.1 Daily mininmum temperature changes of Changqing district sites during the L.lucidum through winter from 2014 to 2017
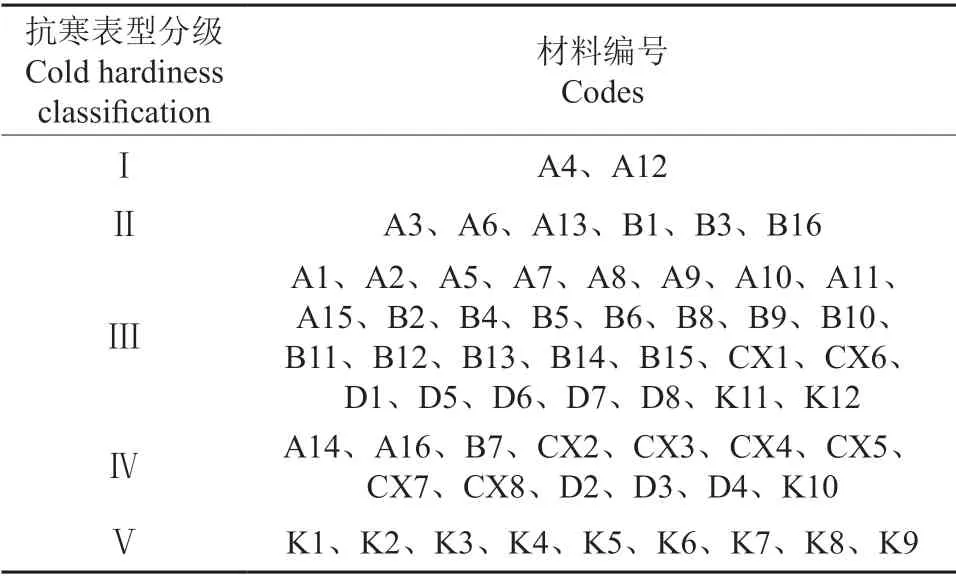
表2 60 份女贞种质材料抗寒表型分级Table 2 Classification of cold resistant phenotype of 60 L.lucidum germplasm materials
2.2 各女贞种质材料的叶片解剖结构
选择越冬叶片保有率较高的女贞种质A4、A5和越冬叶片保有率较低的女贞种质K1、K3,观察其叶片主脉解剖结构,如图2所示。由图2可见,具有较高与较低越冬叶片保有率种质的主叶脉结构无显著差异,其木质部、韧皮部细胞层数接近,细胞大小及排列方式较为相似。

图2 具有不同越冬叶片保有率女贞种质的叶片主脉解剖结构Fig.2 Anatomical structure of leaf main veins of L.lucidum germplasm with different leaf retention rates over winter
选择越冬叶片保有率较高的女贞种质A4、A5和越冬叶片保有率较低的女贞种质K1、K3,观察其叶肉组织解剖结构,如图3所示。由图3可见,越冬叶片保有率较高的女贞种质A4、A5,其叶片的栅栏组织和海绵组织在细胞层数、细胞排列紧密度、细胞完整度等方面显著优于越冬叶片保有率较低的女贞种质K1、K3。

图3 具有不同越冬叶片保有率女贞种质的叶肉组织解剖结构Fig.3 Anatomical structure of mesophyll tissue of L.lucidum germplasm with different leaf retention rates over winter
抗寒种质A4 的叶片栅栏组织多达3 层,栅栏组织厚度达425.2 μm,细胞完整性高、排列紧密,海绵组织细胞解体少、排列紧密,海绵组织厚度297.5 μm,栅栏组织和海绵组织厚度的比值为1.43。越冬叶片保有率较低的种质K3 的叶片栅栏组织厚度仅为198.6 μm,海绵组织细胞排列疏松,海绵组织厚度达435 μm,栅栏组织和海绵组织厚度的比值为0.46。
2.3 各女贞种质材料的低温半致死温度
以60 份女贞种质1年生枝为材料,测定相对电导率,推导Logistic 回归方程,计算低温半致死温度,结果见表3。由表3可知,60 份女贞材料的低温半致死温度与其抗寒表现较为相符,其中种质A4 的低温半致死温度最低,为-15.1 ℃。

表3 60 份女贞种质材料的低温半致死温度Table 3 Semi-lethal low temperature of 60 L.lucidum materials ℃
2.4 各女贞种质材料的ISSR 分子标记和聚类分析
2.4.1 ISSR 引物筛选及扩增产物的多态性
从96 条ISSR 引物中筛选出9 条扩增条带较完整、清晰、多态性丰畗的引物,分别为UBC807、UBC808、UBC810、UBC811、UBC836、UBC840、UBC880、UBC881 和UBC886。所筛选9 条ISSR 引物对60 份女贞种质材料的扩增结果见表4。由表4可知,这9 条引物的平均扩增多态性位点数为224,多态性位点比率达100%。多态性水平无明显差异,扩增的DNA片段集中在200~1 000 bp,60 份女贞种质材料的Nei’s 基因多样性指数为0.116 3~0.172 1,平均为0.143 8;Shannon 信息指数为0.206 0~0.295 3,平均为0.254 5。
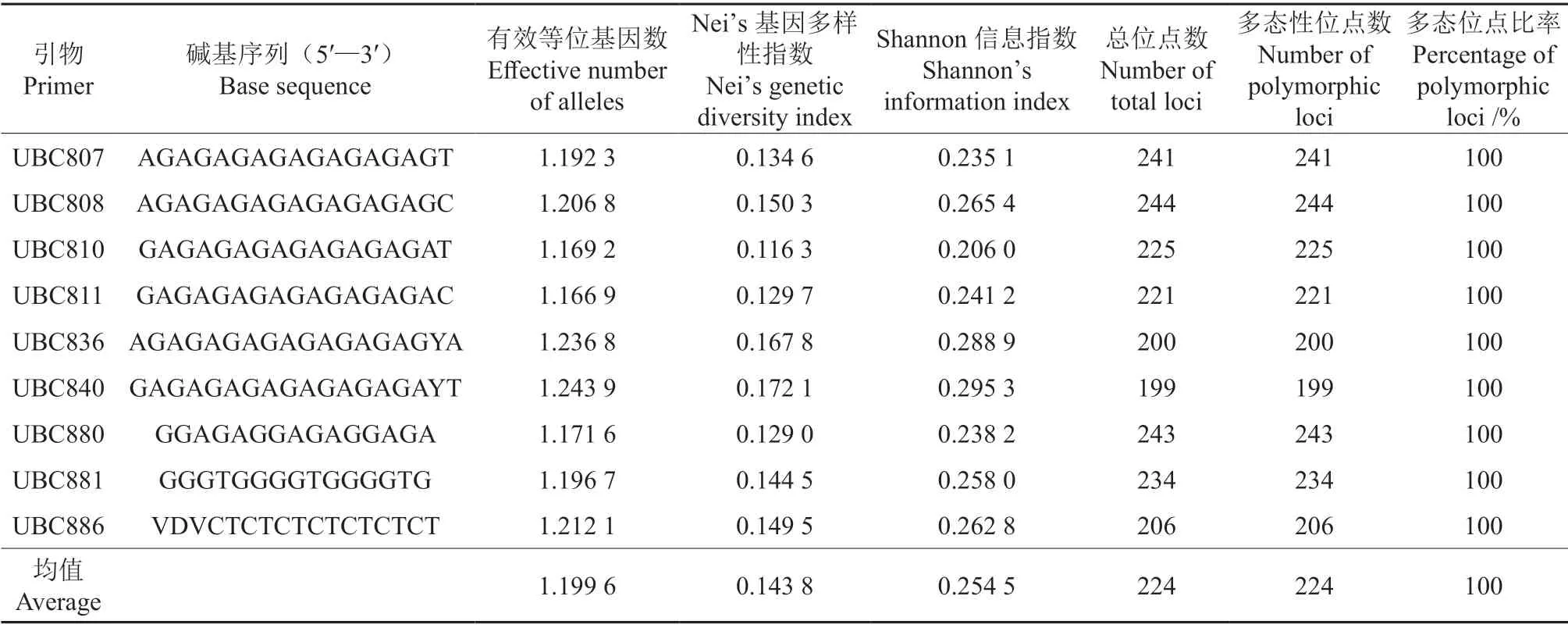
表4 所筛选9 条ISSR 引物对60 份女贞种质材料的扩增结果Table 4 Amplification results of the 9 ISSR primers to 60 L.lucidum materials
使用这9 条引物扩增出的DNA 片段长度基本相同,重复性较高,说明引物的重复性好,可以用于遗传多样性分析。其中,引物UBC808 多态性扩增效果最好(图4),其次是UBC836。
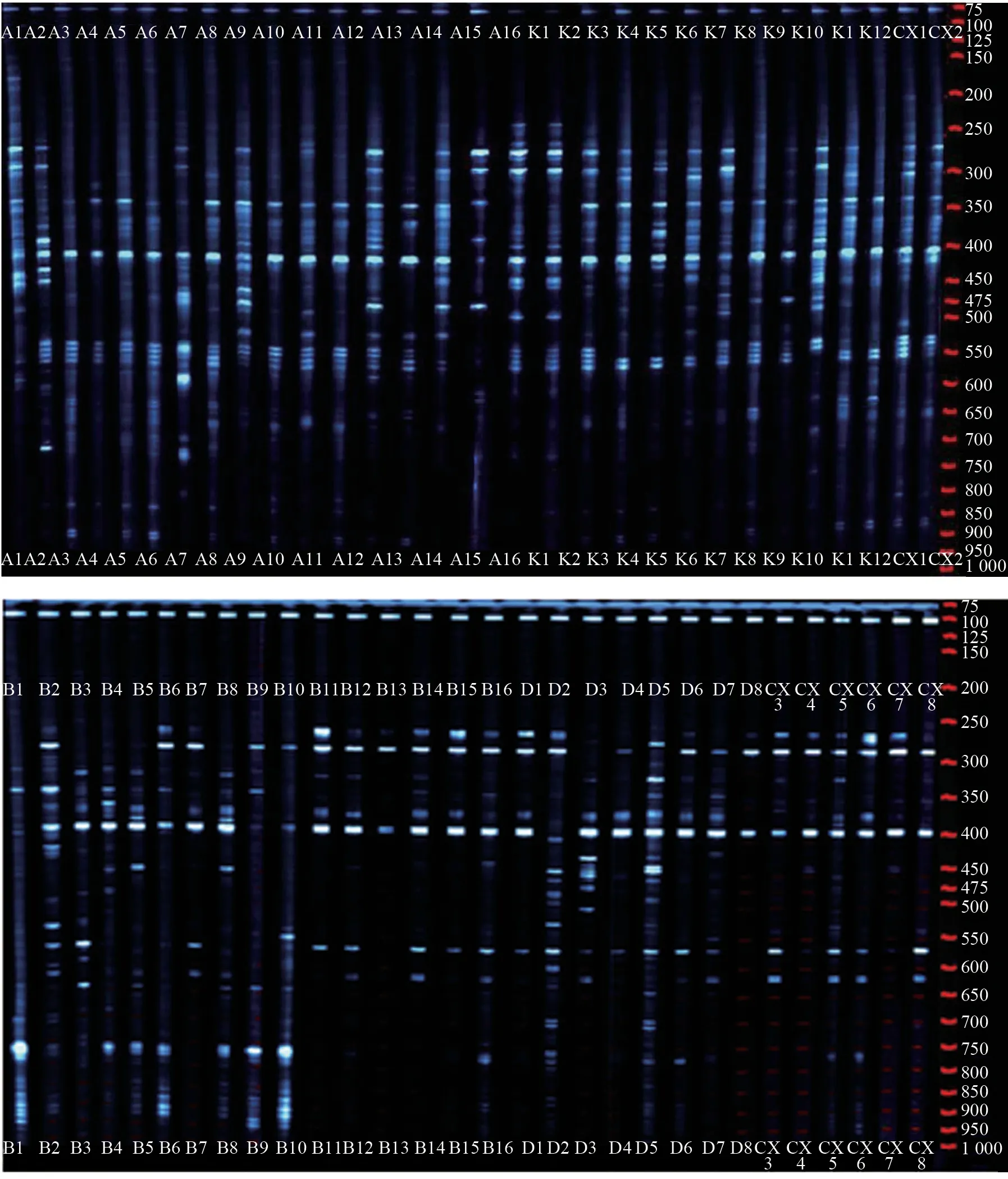
图4 使用引物UBC808 扩增的60 份女贞种质材料的ISSR 图谱(PAGE)Fig.4 ISSR map of 60 L.lucidum germplasm materials amplified with primer UBC808(PAGE)
2.4.2 ISSR 标记聚类分析
根据遗传相似系数矩阵,对60 份女贞种质材料进行UPGMA聚类,结果如图5所示。由图5可见,以相似系数0.298 为界,可将供试样品分为i 和ii 2 大类。i 类中,A8、A10 首先聚在一起,之后与A12、A14 聚在一起,然后与A4、A5 以及A3 聚在一起,A11 和K12 相聚再与前面的种质聚在一起;B6、B8 亲缘关系最近,然后与CX6 相聚;B9、B10 相聚再与B1 相聚。其余的种质聚成ⅱ类。根据表1中女贞种质来源可知,不同种源地的种质可以聚在一起,如A8 和A10;同一种源地的种质不一定聚在一起,同为鄢陵来源的A3、A4、A5聚在ⅰ类,A6、A7 聚在ⅱ类。
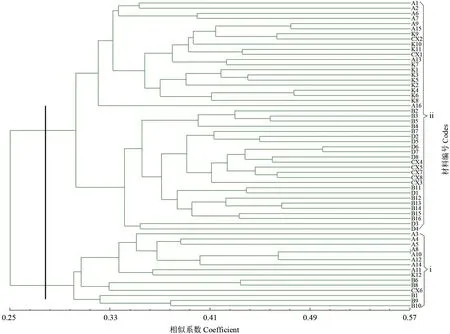
图5 60 份女贞种质材料基于ISSR 的遗传相似性的UPGMA 聚类Fig.5 UPGMA cluster analysis based on ISSR genetic identities among 60 L.lucidum materials
3 结论与讨论
女贞叶肉组织显微结构、低温半致死温度和抗寒表型观测结果均显示优株A4 的抗寒性强且稳定,是良好的抗寒女贞育种材料。供试女贞材料的ISSR 聚类结果反映了其亲缘关系,也表明不能按照优株地理距离聚类。ISSR 聚类结果与抗寒性表现相符,表明亲缘关系近的女贞种质具有相似的抗寒表现。
3.1 ISSR 聚类分析
赵峰等[16]将RAPD 技术应用于女贞种内遗传多样性分析,将7 个居群的121 份女贞材料按照地理来源分为4 大类群。李传代[17]应用ISSR 分子标记将66 份女贞种质分为4 类,亲缘关系远近与地理位置远近的格局不完全吻合。本试验中60份女贞供试材料来自河南、山东、河北、四川等地,ISSR 聚类结果显示大部分女贞种质可按照地域聚在一起,但存在不同取样地的女贞材料未按照地理距离聚类的情况,亲缘关系与取样地域不完全吻合。D5、D6、D7 均为B3 结实播种所得后代,亲缘关系近,聚在一起,说明ISSR 聚类结果反映了其亲缘关系,印证了ISSR 聚类的可信性。本试验中取样材料虽然来自不同省份,但这些省份大多数不是女贞的原产地,当地女贞多是自外地引种而来。李传代等[15]从云南、贵州、江西、湖南、江苏女贞居群中取样,得到的Nei’s 基因多样性指数均值为0.200 5,Shannon 信息指数均值为0.315 1,这2 个衡量遗传多样性的指标均高于本试验结果。表明本试验中所选取女贞种质材料间的遗传变异小于野生分布种群,这与试验取样不是来自野生居群有关,这在高粱[18]、向日葵[19]、大豆[20]等物种上得到证实。
3.2 抗寒女贞优株筛选
大量研究结果表明,采用电导法测定一系列温度下植物组织的电导率,采用Logistic 方程进行回归分析,通过计算得到的植物组织低温半致死温度可以作为抗寒性指标[4-5,21-23]。本试验中通过比较不同女贞材料枝条的低温半致死温度,发现其抗寒性存在明显差异,优株A4 的低温半致死温度达到-15.1 ℃,而抗寒性较弱的种质K1 的低温半致死温度为-4.2 ℃。除K1、K4 外,其余优株均比朱凤荣等[24]测定的河南新乡女贞的低温半致死温度(-5.316 ℃)低,一定程度上说明本试验中所筛选优株的抗寒性整体较强。叶片解剖结构也是反映抗寒性的指标[25]。优株A4 休眠枝条的低温半致死温度为-15.1 ℃,与K1、K3 相比,其叶肉组织解剖结构中栅栏组织细胞层数多、排列紧密,栅栏组织和海绵组织厚度的比值大,说明A4抗寒性强与叶片组织结构有关。将ISSR 聚类结果与2015—2017年的抗寒表型观测结果相比较可知,ISSR 聚类结果中,ⅰ类种质的抗寒表型多为Ⅰ、Ⅱ、Ⅲ级,其中A4、A12 的抗寒表型为Ⅰ级,仅A14的抗寒表型为Ⅳ级。ⅱ类种质的抗寒表型以Ⅲ、Ⅳ、Ⅴ级为多,也包括部分抗寒表型Ⅱ级的女贞种质。将ISSR 聚类结果与低温半致死温度相比较可以看出,ⅰ类种质包含15 个优株,其中10 个优株的低温半致死温度在-10 ℃以下,抗寒能力强,A4 的低温半致死温度最低,而ⅱ类种质的低温半致死温度较高。结合2015—2017年女贞抗寒表型观测结果可发现,ⅰ类女贞种质抗寒性较强,ⅱ类女贞种质抗寒性较弱。在进行抗寒女贞杂交育种时,应选择抗寒强的ⅰ类女贞种质为亲本。
尽管本试验中所选取抗寒指标在多数情况下与抗寒表型观测结果相符,但部分材料存在出入,如A12 的抗寒表型为Ⅰ级,但其低温半致死温度较高(-11 ℃),说明抗寒女贞筛选的复杂性,单凭几年时间的观测进行抗寒优株筛选,其结果不可靠。由于抗寒表型具有不稳定的特点,各种抗寒性筛选指标均有其局限性,单一指标难以准确全面地反映植物的抗寒性[26],将抗寒表型、生理指标、植物组织结构与由分子标记得出的亲缘关系相结合,来筛选抗寒女贞优株,可增强筛选结果的可靠性,得出更为科学的结论。2016年1月,试验地最低温度达到-19.7 ℃,优株A4 的主干和1年生、2年生枝基本未受冻害,其余女贞地上部均呈现不同程度冻害。优株A4 在自然极寒天气的表现与低温半致死温度、叶片解剖结构等抗寒指标及ISSR 聚类结果相互印证,说明其抗寒性较强,可作为抗寒女贞种质在华北地区推广。
植物的抗寒性是其南种北引的主要限制因子,而植物是否真正适应引种地环境,需要几十甚至上百年时间的检验。本研究虽持续7 a,但筛选出的抗寒常绿女贞优株的抗寒表现仍有待时间的检验。除种内选育外,开展种间杂交育种是未来培育抗寒女贞优株的研究方向。
