脂肽微生物的研究进展
2021-07-09陶雪菊苟兴华何成霞
陶雪菊,苟兴华,何成霞
(成都大学 食品与生物工程学院,四川 成都 610106)
0 引 言
微生物脂肽有天然微生物脂肽和合成微生物脂肽两大类.其中,天然微生物脂肽是微生物经过活化培养产生的具有抗菌活性的次级代谢产物;合成微生物脂肽是通过在天然微生物脂肽的结构基础上进行结构修饰而得到[1-2].产生脂肽的微生物有多种,如细菌、放线菌、蓝细菌及真菌等,目前为止,芽孢杆菌是发现产脂肽最多的属类,最早发现的抗菌脂肽也是由芽孢杆菌产生的[3].由于这些微生物具有不同的结构和特性,产生的脂肽也具有不同的抑菌效果[4],如表面活性素类是一种表面活性剂,也具有一定的抗菌能力,伊枯草菌素类具有抗细菌能力,丰原素类具有抗真菌能力.脂肽与其他一些抑菌物质,如与细菌素相比,具有更强的耐热性和耐蛋白水解酶降解能力.脂肽在生物防治、食品加工、石油开采及医药行业有广泛的应用,目前,对脂肽类物质的化学结构及功能研究比较深入,但对脂肽的抗肿瘤、杀虫及溶血栓等生物学活性,有待于进一步研究,这些就激发了许多的科研工作者从不同原料上筛选产脂肽的微生物,以挖掘出新的脂肽菌源,研究不同抗菌物质的生理活性,进而推进脂肽在更多领域的应用[5].
1 抗菌脂肽的结构特征与作用机制
1.1 抗菌脂肽的结构特征
抗菌脂肽一般由7~10个氨基酸残基组成,包含亲水性肽链和亲油性脂肪酸链,其中亲油性脂肪酸链由β-羟基脂肪酸链及β-氨基脂肪酸链两部分组成.微生物脂肽在结构上分为环状脂肽和线形脂肽两大类.环状脂肽又分为脂肪酸成环、接环和离环3种,第一种是脂肪酸链上存在的一个-OH或一个-NH2与肽链上氨基酸的-COOH结合构成酰胺键或者内酯键,导致肽链闭合后形成环状脂肽;第二种是肽环的-NH2直接与肽环相连,肽链的C-端的羧基与N-端氨基酸的-OH或非α-氨基相连形成环状结构称为脂肪酸接环的环状脂肽;第三种是脂肪酸链间接和含有肽键和多种其他酯键、酰胺键等的肽环相连而形成的脂肪酸离环的环状脂肽.线形脂肽又分为螺旋形态和TAN1511(包括TAN1511A、TAN1511B和TAN1511C)两大类,前者为直链结构,后者具有支链结构[6].
研究发现,脂肽类抗菌剂主要包含表面活性素、丰原素、伊枯草菌素、杆菌霉素、抗霉菌枯草杆菌素及制磷脂菌素等[7-8].芽孢杆菌属能产生表面活性素、丰原素和伊枯草菌素3大类脂肽化合物[9].
表面活性素是一类稳定性很高且拥有较高表面活性的表面活性剂,还具有抗真菌、抗细菌、抗病毒和抗肿瘤活性[10].表面活性素肽链典型的氨基酸顺序为Glu(L)-Leu(L)-Leu(D)-Val(L)-Asp(L)-Leu(D)-Leu(L),其分子结构如图1所示[11-12].
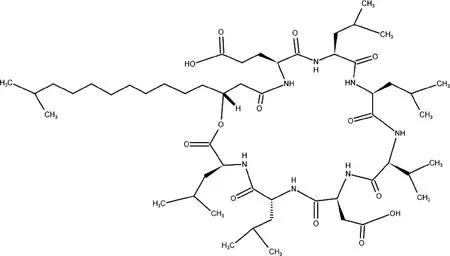
图1 表面活性素分子结构图
丰原素是一类抗真菌抑制剂,特别是丝状真菌产生的丰原素有两种类型,即丰原素A和丰原素B.其中,丰原素A类的肽链第6位上是Ala,丰原素B类的肽链第6位上是Val.丰原素A的分子结构如图2所示[13].
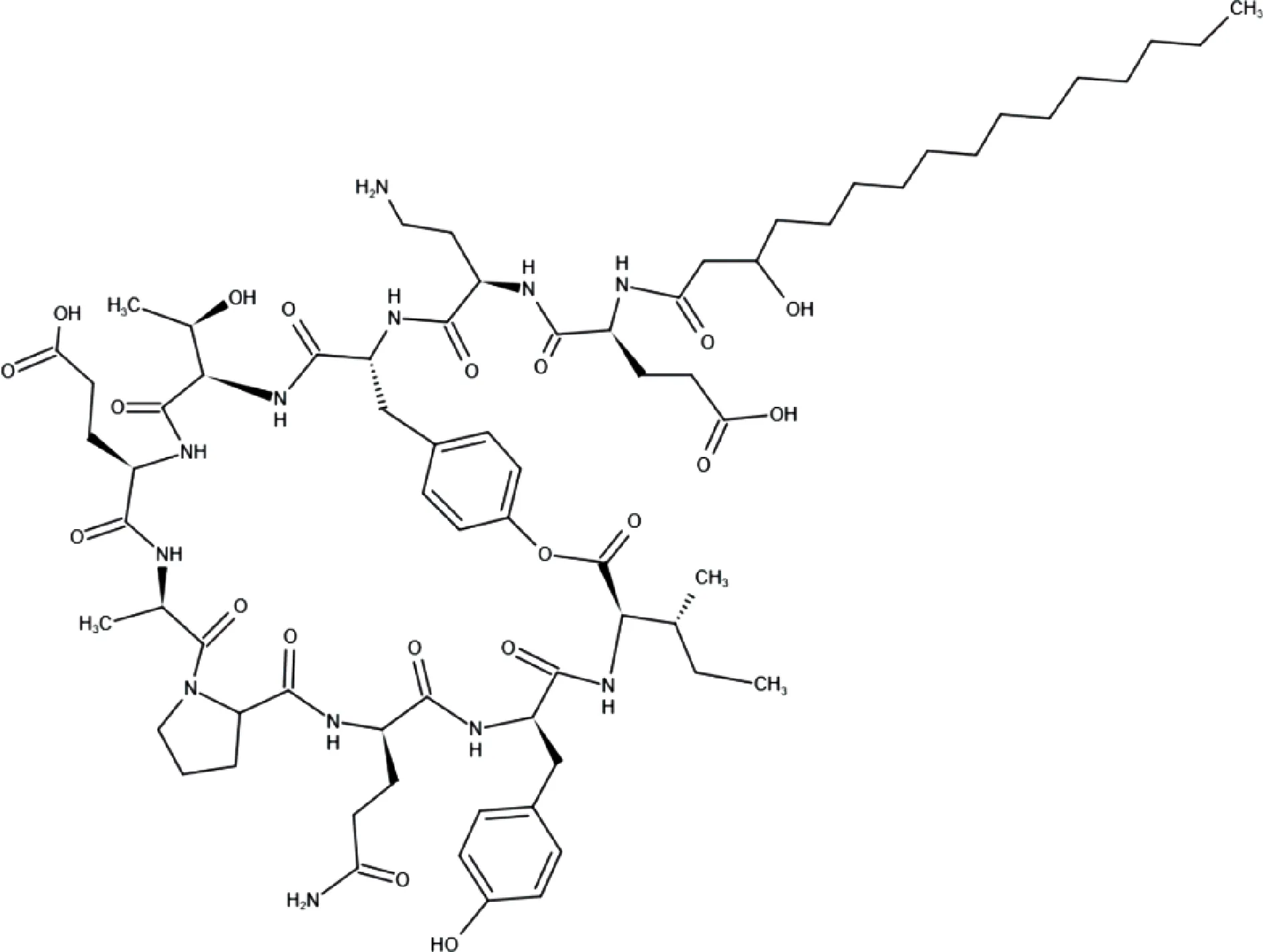
图2 丰原素A分子结构图
伊枯草菌素是一类脂肽化合物,有多种类型,包括伊枯草菌素A、B、C、D和E,芽孢菌素D、F和L,以及抗霉枯草菌素等.其中,伊枯草菌素A的分子结构如图3所示.
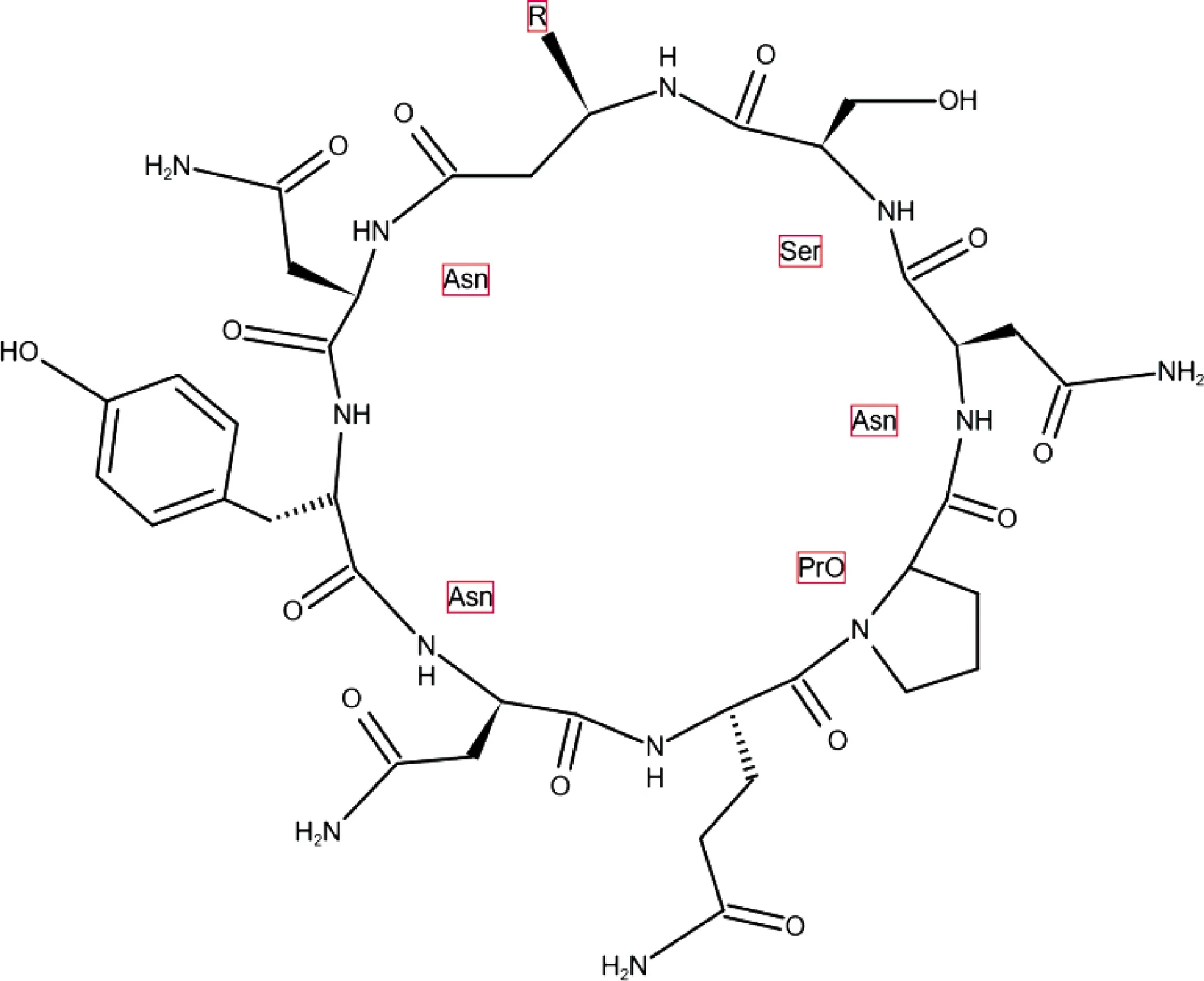
图3 伊枯草菌素A分子结构图
微生物产生的抗菌脂肽种类较多、结构较复杂、性质差异也较大.但是其中一部分脂肽化合物会有诸多类似物,且结构相类似,性质上也相近,因此在筛选及分离纯化上存在困难,而且产量较低,这就给大规模的分离纯化工艺带来了麻烦.
大部分的芽孢杆菌都能分泌具有抗菌活性的物质,主要包括表面活性素、杆菌霉素、乳酸链球菌肽、丰原素和伊枯草菌素等(见表1).对脂肽的筛选鉴定及其活性探索也主要集中于芽孢杆菌类,霉菌和乳酸菌的研究次于芽孢杆菌,且这些能产脂肽物质的菌株主要是革兰氏阳性菌,有的具有广谱抗菌性,一部分只具有抗真菌特性等.
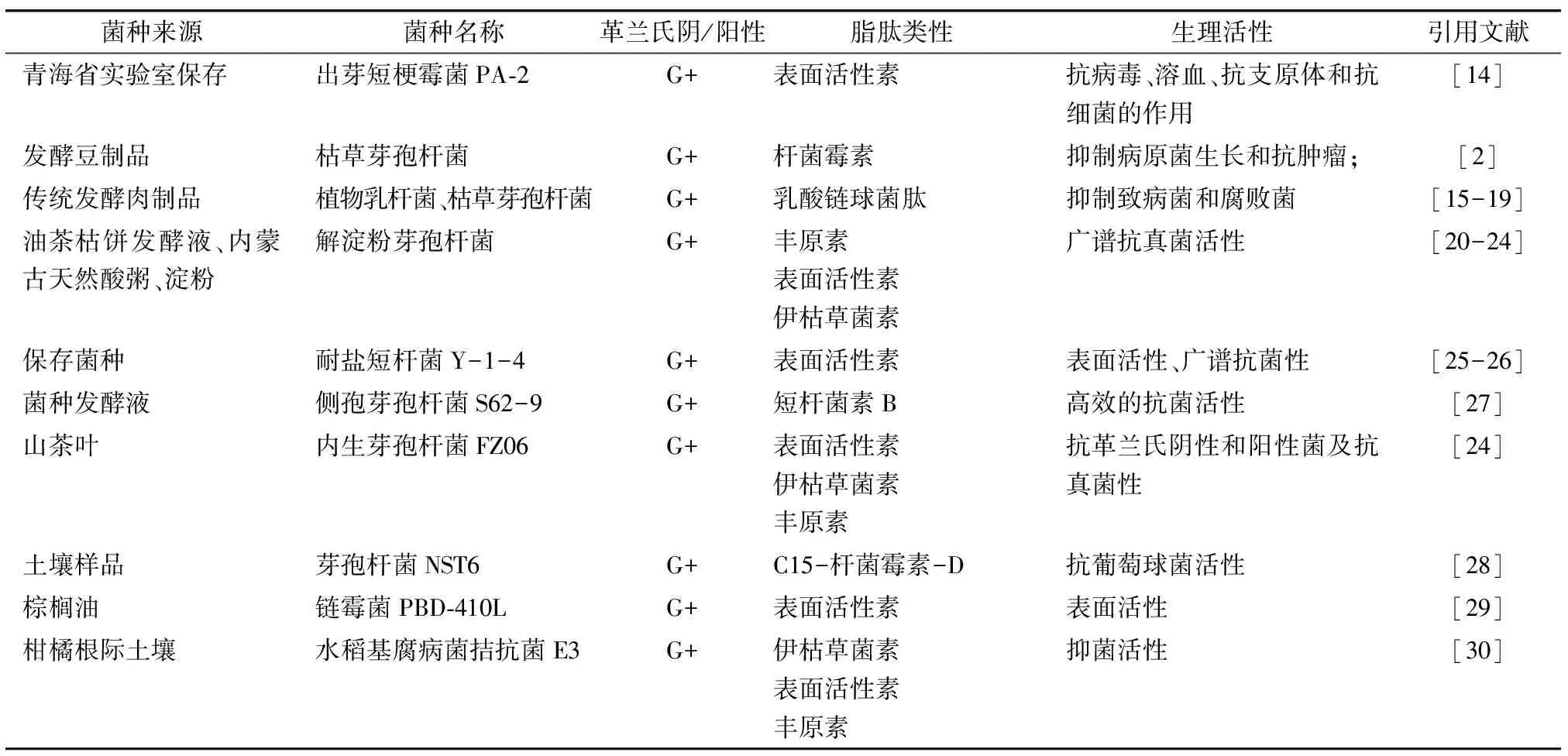
表1 部分脂肽及微生物类型及活性概述
1.2 抗菌脂肽的作用机制
1.2.1 抗菌脂肽与细菌细胞壁的作用
抗菌脂肽通过穿透细菌磷脂膜外面的细胞壁来抑制细菌的生长,借助其静电引力来穿过细胞壁,抗菌脂肽与细胞壁上多糖之间的强静电作用,与脂质之间的疏水作用以及脂质与抗菌脂肽间的氢键增多也会使抗菌脂肽活性增强,进而提升其抑菌率[31].例如,人源抗菌脂肽LL-37和抗菌脂肽SMAP-29被用来抑制肺病原菌,研究发现,其作用时主要借助抗菌脂肽与细菌细胞壁上多糖之间的相互作用而影响其生物学活性[32].其中,荚膜多糖主要存在于周围介质中,在静电引力下,与细菌膜外面的脂多糖带相结合后,抗菌脂肽就会引起膜结构的异常变化,利用其较强的亲和力能从带有负电荷的脂多糖处转移一部分阴离子,造成该部位的细胞壁紊乱,改变其稳定性后,抗菌脂肽便能穿过该区域来达到抑制细菌的目的.对于革兰氏阳性菌来说,细胞壁表面的磷壁酸本身带有的负电荷,可与抗菌脂肽的正电荷借助静电引力并与之结合,使抗菌脂肽能够顺利通过革兰氏阳性菌的细胞壁,再靠近细胞膜,并对细胞膜发挥作用[11].
1.2.2 抗菌脂肽与细菌细胞膜的作用
从抗菌脂肽卤素-1与各种膜模型发生相互作用而引起的脂肽结构变化,可以得出肽模式的作用机制,该抗菌脂肽氨基酸序列在特定位置修饰,会导致两亲性的增加,蛋白质插入受限,进而致使抗菌性及溶血活性发生变化,导致对模型膜的不同反应[33].抗菌脂肽的作用方式的机制涉及脂肽与细胞膜之间膜孔的形成,抗菌脂肽与膜的相互作用在其活性机制中起到关键作用,并且抗菌脂肽主要以地毯式排列于脂质与水的表面.有抗菌脂肽存在时,脂质体的负电位降低,多肽的插入扰乱了有序排列的脂质,同时破坏细菌的细胞膜,降低其有效参数,便于水分子渗透过膜,并使得胞浆内物质流出,引起靶细胞的死亡[34].
但由于存在静电排斥,阳离子抗菌脂肽不会与阳离子两亲体相互作用,而只会和阳离子聚集体系相互作用,不与阳离子单体发生作用[35].抗菌脂肽在高度疏水的介质中趋于α-螺旋,而在水介质中则采用随机螺旋结构,螺旋度对抗菌脂肽与膜的亲和力有影响,并且脂质囊泡可以促使肽排列成更多的螺旋结构[36].但这种螺旋结构的疏水相互作用很弱,需要具有大面积的聚集态来形成跨膜离子通道[27],由于肽链的疏水性会随着肽链的缩短而延长,且与抗真菌活性间为协同作用,因此双头脂肽和脂三角相比脂五肽就会形成更小的、更具动态的聚集体,作为胶束或单体的阴离子两亲体与抗菌脂肽因为静电作用,膜电势遭到破坏,脂肽构象变化,最后导致细胞质膜分离和细胞质物质的损失,进而无法维持正常的生命活动而死亡[37].
1.2.3 抗菌脂肽与细菌胞质内靶目标的作用
抗菌脂肽与膜的结合不是直接将细胞杀死,而是其进入细胞后攻击靶细胞的一个必要过程.先通过细胞壁和膜进入细胞体内,后出现核物质浓缩而发生异常间隔,进而杀死细胞.抗菌脂肽的抑菌机制因为其代谢产物的多样性而多种多样,如芽孢杆菌菌株w176产生的细胞上清液(CFS),有多种不同的代谢物,包括大环内酯抗生素、抗霉枯草菌素、聚酮合酶抗生素和表面活性素.当青霉菌与CFS共同培育后,其亚细胞结构发生了液泡化变化.用CFS处理马尾松后,核糖体生物会胁迫相关基因转录水平发生变化,分析得出CFS是一种新的可用于防治柑橘绿霉病的生物制剂[38].除此之外,一些枯草芽孢杆菌能够产生纤维素酶、分泌蛋白酶及脂肽类抑菌物质,这些抑菌物质会使病原菌的菌丝产生膨大畸形,从而抑制病原真菌的生长[39],并且这些产生抑菌物质的微生物和病原菌之间会存在对养分和空间的竞争,利用自身生长发育产生拮抗剂来尽可能地消耗可以利用的营养物质,从而抑制病原体的生长[40].其中,抗性诱导系统是一种常见的抑菌机制,用拮抗剂来诱导宿主菌的防御反应,促使目标基因的表达,进而激活不同的化学物质,进行防治感染[41-42].
2 抗菌脂肽类微生物源
2.1 芽孢杆菌属
枯草芽孢杆菌是一类耐酸碱、耐热、好氧或兼性好氧并产芽孢的革兰氏阳性杆状细菌[43].其分布广泛、抗逆性强,且枯草芽孢杆菌含有适合用于合成抗生素的基因,一些菌株可以产生蛋白酶和脂肪酶等消化酶,合成多种生物活性物质及产生抗菌脂肽等[44-45].抗菌脂肽包括伊枯草菌素、表面活性素和丰原素3类[46],其产生抗菌物质分为两个途径:其一是通过非核糖体合成脂肽类抑菌物质,也是主要的抑菌物质之一[47];另外一种是通过核糖体合成蛋白类抑菌物质,这些蛋白类多是可以裂解细胞壁的酶类,导致细胞裂解,最后导致死亡[48].
胡美忠等[49]筛选到一株能产生抗菌脂肽的芽孢杆菌,且该抗菌脂肽抑菌谱广,耐酸碱性和热稳定性好.多粘类芽孢杆菌对致病菌和腐败菌有显著的抑制作用,该菌产生的抑菌物质为蛋白类的细菌素,目的是作用于有害菌的细胞壁及细胞膜,进而达到抑菌效果[50];地衣芽孢杆菌FJAT-4 产生的抗菌脂肽为表面活性素和丰原素两大类物质,该抑菌脂肽会致使辣椒、香蕉和番茄尖孢镰刀菌等多种植物致病菌,尤其是尖孢镰刀菌的菌丝体发育畸形,从而影响该菌的正常生长[51].此外,甲基型芽孢杆菌产生的多种抑菌物质单独使用和协同发挥的抑制使用效果不一,多种抑菌物质共同加入到含致病菌发酵液中进行发酵后,产生的上清液和沉淀里抑菌带的明显程度也有所差异,即表现出不同程度的抑菌活性,这为脂肽的抗菌机制的研究提供了依据,并且还能在发酵过程中产生胞外抑菌物质,对霉菌和致病菌都有一定的抑制作用,因此是一类具有潜在价值的微生物[52-53].
2.2 乳酸菌属
乳酸菌是一种益生菌,因具有发酵产酸的特性,而被广泛应用到酸奶、发酵面包及酵素饮料等食品中.乳酸菌还能产生一些抗菌脂肽及蛋白类物质,在食品的生物防腐等方面存在着重要的作用,在抗菌脂肽方面也存在极大的开发潜力.
依据抗菌脂肽的抑菌谱、抑菌机制和耐热性等可以将抗菌脂肽分为以乳酸链球菌素为代表的羊毛硫抗菌脂肽、非羊毛硫抗菌脂肽、大分子耐热性多聚肽以及含有多糖、阳离子和脂质的多肽这4大类[17].杨云喜等[18]筛选了一株产抗菌脂肽的乳酸菌,该抗菌脂肽对真菌无抑菌活性,但是对细菌具有广谱抑菌性,通过优化能够产生抗菌脂肽的培养基成分及培养条件,为其大规模地应用提供了基础.赵乙桢等[19]从新鲜牛奶中分离到一株能产生广谱抑菌性抗菌脂肽的鼠李糖乳杆菌ZJ225.赵冬兵等[54]从板鸭中筛选得到了一株魏斯氏菌,其产生的抗菌脂肽对较强酸碱都具有稳定性.章检明等[17]在91株乳酸菌中筛选出了7株产生抗菌脂肽的乳酸菌,有魏斯氏菌属、鼠李糖乳杆菌属、葡萄球菌属及微球菌属4种,其中鼠李糖乳杆菌属能产生具有广谱抑菌性,且性质较稳定的多肽.
2.3 霉 菌
相比其他属类的微生物而言,对霉菌产生抗菌物质的研究较少.有研究者探索了部分霉菌的抑菌活性,通过优化出芽短梗霉菌PA-2的发酵温度、转速及装液量等,得出该菌产生的脂肽物质为表面活性剂,具有较好的发泡和乳化作用[14,55].李志田等[56]筛选鉴定出了一株浑圆链霉菌QH-16,经离体测定,其对多种细菌及植物病原菌具有很好的抑菌作用.张萍华等[57]探索了黑翅土白蚁巢中链霉菌发酵产物的抗菌活性,并对其抗菌成分进行研究,发现其对金黄色葡萄球菌具有很好的抑制作用,具有开发成为新型微生物源杀菌剂的潜力.
毛霉是一类高效微生物,能够在单一培养基中利用可再生来源——碳源,来生产生物糖脂、生物乳化剂和表面活性剂等,毛霉在相同的培养基和不同的物理条件下实现了生物表面活性剂和乳化剂的生产.此外,可以肯定的是,生产的表面活性剂和乳化剂满足可持续性和附加值的基本要求,因为它们是获得各种工业部门日常生产的化合物的主要原料[58].
3 抗菌脂肽的应用
3.1 在食品中的应用
随着社会的发展,消费者对食品安全的要求在不断提高,但是滥用及违规使用添加剂等造成的添加剂类食品安全事故也时有发生.此外,还有食品自身的变化导致的污染,如黄曲霉,其寄主范围非常广,辣椒、玉米、棉花以及花生等多种作物都会被其污染,不仅会造成巨大的经济损失,还在污染作物的过程中产生黄曲霉毒素,人和畜禽误食后会引发中毒,严重的可能会致癌和导致死亡[59].在这样的环境背景下,类似于抗菌脂肽等生物制剂得到了消费者的青睐.
抗菌脂肽在食品行业的主要应用是作为生物防腐保鲜剂和食品添加剂.抗菌脂肽的表面活性剂具有乳化作用,可用作食品加工中的发泡剂和乳化剂,以改善食品的质构、色泽和口感.另外,抗菌脂肽的抑菌活性又可作为防腐保鲜剂应用于食品中.张学雯等[60]分离鉴定了黄曲霉拮抗菌B10-6-1拮抗物种类,并对其结构等进行了研究,这为黄曲霉的有效控制提供了依据.
3.2 在饲料生产中的应用
随着消费者对肉类的需求不断增加,促进了各地养殖业的发展,但是由于养殖地分布各地,控制上就容易出现问题.抗生素饲料的滥用会导致动物体内的抗生素通过畜产品、水、土壤等传递给人类,导致人体产生大量耐药菌株,对人类的健康造成危害.因此,绿色且安全、无残留及不易产生耐药性的新型生物饲料添加剂备受关注,已成为诸多科研工作者的研究方向.
抗菌脂肽能够成为新型饲料与其结构有关,其结构是由易于消化吸收的脂肪酸链和氨基酸肽链两部分组成.抗菌脂肽可以部分取代用于饲喂的抗生素,避免抗生素在动物体内过多蓄积而沿食物链危害人体健康.另外,抗菌脂肽在抑制革兰氏阳性菌和部分革兰氏阴性菌的同时,又能够促进其生长,且此过程不会被肠道吸收,只在消化道内起作用,因此一般无残留,效果稳定,也不易产生细菌耐药性[11].
3.3 在医药行业的应用
为了治疗细菌感染,抗生素被大量应用,其作用效果是通过破坏细菌的DNA、细胞膜和细胞活性以及用其来结合细菌的一些化合物来杀死细菌.抗菌脂肽能与两亲性生物可降解聚合物自行组装而成,形成的HnMc胶束能有效地结合细菌膜[61],从而杀灭广谱细菌[62].抗菌脂肽因为菌源不同,以及自身结构的不同导致了抗菌性能的差异.其中,肽链中脂肪酸长度的改变导致伊枯草菌素具有多样性,可以防止多种生物性植物病原菌,丰原素具有治疗皮肤真菌病及具抗真菌的作用[63-64].
抗菌脂肽因其抗黏附性能,可以抑制生物膜的产生和细菌黏附在感染部位的能力[1],因此利用这种能力生产抗生素和治疗性疫苗,可以治疗革兰氏阳性菌及病毒的感染.有一种环状类型的抗菌脂肽,由非极性的脂肪酸链通过结合极性的氨基酸脂肪链组成,其在多重耐药病原细菌方面具有独特的抑菌能力.正是因为代谢产生的脂肽能够较强地抑制革兰氏阳性菌的生长,如抑制结核分枝杆菌与肺炎双球菌等细菌,可应用于疫苗的生产中[6].有研究表明,含有抗菌脂肽的蛋白质与不含抗菌脂肽的比较,在无外部佐剂的情况下能够诱导强烈的病毒跨膜蛋白的特异性抗体反应,此类重组基因的蛋白质有望成为一种疫苗[65].脂肽在国外已被应用于预防和治疗药物,甚至有研究表明,微生物脂肽还能在预防癌症方面有所应用.
3.4 在农业中的应用
现在农业上受到多种病虫害的威胁,不仅影响了农产品的质量,还大大降低了农产品的产量和农户的收入[66].目前,大部分种植者的观念仍是化学防治效果显著,因此广泛应用化学物质来防治农业病虫害,但是,这些化学用品可能会对生态环境和人类生活产生负面影响.因此,近年来生物防治被广大研究者所推崇,被认为是一种安全和可持续的方法,其中芽孢杆菌能够产生一些抗菌脂肽类物质,具有广谱抗菌活性,因此被广泛用于农业的生物防治[67].枯草芽孢杆菌已被选为脂肽类抗生素的生物农药的菌株之一,它能产生对烟草病毒及辣椒病毒有防治效果的脂肽类抗生素表面活性素[46],能用于控制柑橘绿色和蓝色霉菌[68],以及防治辣椒病害、黄瓜病害及油菜菌核病[26].另外,解淀粉芽孢杆菌产生的有抑菌作用的活性物质C14-杆菌霉素-D和C15-杆菌霉素-D,可以用于合成新型生物农药,以解决黄曲霉污染防治的瓶颈问题[59].
4 讨论与展望
现在已经发现的产生抗菌脂肽芽孢杆菌种类颇多,但其他属类的研究较少.另外抗菌脂肽的同系物种类繁多,其发酵液成分复杂,很难大规模生产和浓缩纯化出足量的纯品,甚至许多产物还没有一种标准的检测方法.因此,为了更加高效地利用这些微生物来产生抗菌物质,应该对其结构进行深入研究,才能找到提高其产量的方法[55].从目前来看,脂肽微生物的研究可以从以下几方面进行:
1)抗菌脂肽的抑菌机制仍然主要是对细胞壁、细胞膜及靶细胞的作用,可以根据其结构进行更深入研究,探索其具体的基因作用位点、抑菌机制和应用范围等.可以通过从基因水平来探究抗菌脂肽的结构,目的是要获得相关的转录调控因子,如用自杀载体等方式来获得突变菌株,抗菌脂肽产量会有所下降,再控制相关的正向调控因子发挥正调控作用,对突变基因进行回补后,脂肽产量会有所恢复[69].另外,还存在阻遏蛋白对脂肽进行调控,当分别含有两种阻遏蛋白的菌株,其控制基因突变后再利用载体回补,这两种控制基因中的任意一个发生突变,另一个阻遏蛋白的表达量有所上调,说明这两种阻遏蛋白相互抑制,在整个系统中协调发挥作用,基因突变后阻遏蛋白表达量的增加伴随着抗菌脂肽产量的增加,为正调控因子,阻遏蛋白的增加伴随着抗菌脂肽产量的减少为负调控,正向调控的蛋白可以抑制阻遏蛋白而起到正向调控的作用[70].因此,产脂肽微生物的筛选鉴定及脂肽的分离纯化方面可以从基因的调控入手,缩小筛选范围、提高实验效率.
2)抗菌脂肽在应用上还存在一些问题,如黄曲霉毒素容易污染食品而造成食品安全问题,尽管目前为止己筛选到很多抑制黄曲霉生长的化合物,但几乎没有应用到实践中来,因此需要注意与实践相结合,拓宽应用范围[71].
3)将抗菌脂肽用于临床上还存在许多问题,抗菌脂肽本质是蛋白质,应用过程中容易被降解,需要包埋或修饰处理,而且在毒力学和药效学等方面研究较少,需要进一步的探讨脂肽类物质对人体的安全问题的影响,提供一些支持性的研究报告.
4)基因工程是抗菌脂肽的主要获得手段,其成本较高,生产效率较低,因此应该加大科学研究,如优化发酵条件、工艺设计,改良菌种、载体及降低成本等,逐渐向研发出性能更好、规模更大的生物制剂的方向发展[72].
