MAGEA1通过激活Wnt/β-catenin信号通路促进恶性黑色素瘤的转移
2021-06-29许媛媛李永君郝永红胡辉莹方向东
许媛媛,王 晨,贺 赞,王 冬,杨 琼,李永君,郝永红,胡辉莹,方向东,赵 华
1 解放军总医院第一医学中心 皮肤科,北京 100853;2 中国科学院北京基因组研究所 基因组科学与信息重点实验室,北京 100101;3 南部战区总医院(白云院区) 皮肤科,广东广州 510510
恶性黑色素瘤是来源于黑色素细胞的恶性肿瘤,多发生于皮肤,发病率在皮肤恶性肿瘤中占第3位,是皮肤恶性程度最高的肿瘤。近年来,恶性黑色素瘤的发病率持续上升,全球每年约有300000例新病例和60000例死亡病例[1]。早期和原位黑色素瘤,立即手术切除是治疗的首选方法,预后良好。转移性恶性黑色素瘤虽然只占所有恶性肿瘤的1%,但却是最致命的[2],已成为危害人类健康的严重疾病之一。
黑色素瘤相关抗原家族A1(melanoma associated antigen family A1,MAGEA1)是MAGE-A基因亚家族的成员,又名癌症-种系(cancer-germline,CG)基因或肿瘤-睾丸抗原(cancer-testis antigen,CTA)基因。MAGEA1可在多种恶性肿瘤中表达,如黑色素瘤、子宫内膜癌、喉鳞状细胞癌、非小细胞肺癌和胃癌等[3-7],其致病机制包括参与泛素化过程调节细胞的凋亡[8]、启动甲基化影响肿瘤的发生发展等[9]。我们前期的研究已经证实MAGEA1可以通过激活c-Jun促进黑色素瘤的迁移和侵袭[10]。为进一步研究MAGEA1调控黑色素瘤转移恶化进程的分子机制,本文通过对公共数据中原位和转移性黑色素瘤患者的MAGEA1表达量进行差异分析,将过表达MAGEA1的A375和对照细胞系转录组数据富集至KEGG信号通路中,探讨Wnt/β-catenin信号通路在黑色素瘤的恶变过程中发挥的重要作用,并通过MAGEA1差异表达的A375与A2058细胞系进行体外验证,为进一步研究黑色素瘤的恶变机制提供了一定的理论依据。
材料及方法
1主要试剂与材料 A375与A2058细胞系购买自美国ATCC细胞库;Dulbecco’s modified Eagles’s medium (DMEM)购自美国Gibco公司;胎牛血清(FBS)购自澳大利亚AusGeneX公司;Trypsin0.25 EDTA(胰酶)、青霉素/链霉素、Trizol试剂购自美国Life technologies公司;Dimethyl sulfoxide (DMSO)购自美国Sigma公司;RIPA强效裂解液、免疫染色强力通透液购自碧云天公司;BCA Protein Assay Kit BCA法蛋白检测试剂盒购自美国Thermo Fisher公司;TCF报告质粒试剂盒(TOP flash,FOP flash)购自德国MERCK公司;双荧光素酶检测试剂盒购自美国Promega公司;ProLong(R) Gold Antifade Reagent with DAPI、p-GSK-3β(5558)、E-cadherin(3195s) 抗体购自美国Cell Signaling Technology公司;MAGE-A1(sc-71539)、c-Jun(sc-74543)、p-c-Jun(sc-822)、LamiA(sc-20680) 抗体购自美国Santa Cruz公司;β-catenin(YM3403)、c-myc(YT0991)、GSK3β(YM3633) 抗体购自美国ImmunoWay公司;GAPDH(ab128915)、Vimetin(ab8069)、Twist1(ab50887) 抗体购自英国Abcam公司;山羊抗小鼠和抗兔多克隆抗体(ZB-2305,ZB-2301)购自ZSGB-Bio公司;山羊抗小鼠和抗兔荧光抗体、pcDNA3-Neo-EGFP质粒、Lipofectamine2000转染试剂购自美国Invitrogen公司;宽范围彩色预染蛋白质Marker(11~245kU)购自TIANGEN公司;牛血清白蛋白V(BSA)购自瑞士Roche公司;Revert Aid First Strand cDNA Synthesis Kit(反转录试剂盒)购自美国Thermo Scientific™公司;SYBR®Fast通用型购自美国KAPA公司;TCF7 siRNA (Human)、siRNA Negative Control购自Cohesion公司;基质胶Matrigel Matrix购自美国Biosciences公司;苏木素-伊红染色液(H&E):珠海贝索公司;通用SP试剂盒(小鼠/兔链霉卵白素-生物素法检测系统)(SP-9000)、柠檬酸盐修复液pH 6.0、DAB kit、中性树胶购自北京中杉金桥公司。
2生物信息学分析 从TCGA公共数据中原位和转移黑色素瘤患者RNA-Seq数据,提取MAGA1表达数据(FPKM),通过Graphpad软件绘制原位和转移黑色素瘤患者FPKM散点图,并进行t检验。MAGEA1过表达A375(A375-MAGEA1)和对照细胞系(A373-con)的RNA-seq数据,使用Limma(R语言包)进行差异表达分析,并将结果进行KEGG富集分析(在线分析网站www.genome.jp/kegg)。
3克隆与质粒构建 聚合酶链反应(polymerase chain reaction,PCR)从正常人血液扩增出MAGEA1蛋白的cDNA,并将其克隆到pcDNA3-Neo-EGFP质粒中。将针对MAGEA1的短发夹状RNA(short hairpin RNA,ShRNA)克隆到pRNATU6.1-Neo-EGFP质粒中,构建成载体pRNATU6.1-Neo-EGFPshMAGEA1。表1提供了shRNA序列。为了避免shRNA可能的脱靶,我们设计了siRNA来下调黑色素瘤细胞系中MAGEA1的表达。表1提供了两个最有效的siRNA(siRNA3和siRNA4)的序列。所有构建的载体均经限制性内切酶酶切和标准DNA测序验证。

表1 shRNA与引物信息Tab. 1 shRNA and primers sequences
4细胞培养和转染 A375细胞株来源于皮肤原位黑色素瘤组织,A2058细胞株来源于恶性黑色素瘤患者转移淋巴结。对照细胞株(A375-con,A2058-con)和过表达MAGEA1稳定细胞株(A375-MAGEA1,A2058-MAGEA1),敲低MAGEA1稳定细胞株(A375-shMAGEA1-1,A375-shMAGEA1-2,A2058-shMAGEA1-1,A2058-shMAGEA1-2,A375-MAGEA1-NC,A2058-MAGEA1-NC,A375-MAGEA1-siTCF7,A2058-MAGEA1-siTCF7)和阴性对照细胞株(A375-scramble,A2058-scramble)均通过Lipofectamine2000转染构建。用G418筛选转染细胞2周,分离培养单个集落。所有细胞均用90% dMEM、10% FBS和0.1mg/mL双抗配制的培养液培养,置于37℃,5% CO2恒温培养箱中培养。
5β-catenin/TCF转录报告分析 将105个细胞(A375-con,A2058-con,A375-MAGEA1,A2058-MAGEA1,A375-scramble,A2058-scramble,A375-shMAGEA1-1,A375-shMAGEA1-2,A2058-shMA GEA1-1,A2058-shMAGEA1-2)接种于48孔板中,加入250µL无双抗培养基,37℃培养24 h,细胞密度达90%~95%时进行转染。在TopFlash或FopFlash转染后36~48 h,使用双荧光素酶检测试剂盒进行检测并记录结果。
6蛋白质的提取和检测 收取细胞(A375-con,A2058-con,A375-MAGEA1,A2058-MAGEA1),加入适量4% SDS吹打混匀,于95℃金属水浴锅煮沸30min;4℃,13500r/min,离心5min;将上清移至新的离心管得到总蛋白。细胞中核浆蛋白分离:细胞经过处理后,4℃,1000r/min离心5min,弃上清,加入适量的裂解液A(20mmol/L HEPES,pH7.0,10mmol/L KCl,2mmol/L MgCl2,0.5% Nonidet P-40,2mmol/L Na3VO4,2mmol/L NaF,2mmol/Lβ-甘油磷酸纳,1% Protease inhibitor cocktail),匀速缓慢吹打多次。4℃,15000 g离心5min。将上清组分转移到新的离心管中,4℃,15000 g离心5min,得到的上清即为细胞质蛋白组分。用裂解液A清洗沉淀2次,4℃,15000 g离心5min,得到的沉淀即为细胞核蛋白组分。各加适量的强效裂解液超声破碎细胞核,能量和时间:15%能量,开2s,停4s共超声10s。离心机4℃,15000 g离心5min,将上清溶液转移至新的管中,即为提取的细胞核蛋白组分。用BCA法检测蛋白浓度,最后使用Western blot检测Wnt/β-catenin信号通路与EMT相关蛋白表达差异。
7免疫荧光染色 将圆形盖玻片浸泡于浓盐酸中2 h,室温干燥后置于75%乙醇中浸泡30min。烘干后进行细胞(A375-con,A2058-con,A375-MAGEA1,A2058-MAGEA1)爬片,接种适当密度细胞,培养24 h后用4%多聚甲醛进行细胞固定30min。透化液于室温下处理15min后,使用5% BSA/PBS溶液封闭30min。室温下一抗孵育2 h,室温下二抗避光孵育1 h,PBS漂洗后进行封片,切片用激光共聚焦显微镜观察并采集图像保存。
8实时荧光定量PCR分析 使用Trizol裂解细胞(A375-MAGEA1-NC,A2058-MAGEA1-NC,A375-MAGEA1-siTCF7,A2058-MAGEA1-siTCF7),加入氯仿和异丙醇提取细胞总RNA并进行浓度测定和质量检测,反转录合成cDNA(Thermo Scientific公司试剂盒)。采用CFX96实时荧光定量PCR检测系统进行基因表达分析(所有基因的引物序列见表2)。

表2 实时荧光定量PCR引物序列Tab. 2 Sequences of real-time quantitative PCR primers
9细胞侵袭和迁移实验 接种细胞(A375-MAGEA1-NC,A2058-MAGEA1-NC,A375-MAGE A1-siTCF7,A2058-MAGEA1-siTCF7)前1 d,将人工基质胶与预冷的DMEM培养基(无血清和双抗)按照1∶7的比例以总量40 µL铺在Transwell小室的上室面进行预处理(迁移实验不需要此步),接种细胞数为2×104(迁移实验)和2×105(侵袭实验),细胞培养24 h (迁移实验)和48 h(侵袭实验)后进行固定和HE染色,最后将小室置于显微镜下观察、拍照并计数。
10统计学分析 统计学分析采用SPSS19.0软件,以上实验至少重复3次,多组比较采用单因素方差分析,P<0.05为差异有统计学意义。
结 果
1MAGEA1可能会影响Wnt/β-catenin信号通路进而引起黑色素瘤的转移 通过分析TCGA公共数据中原位和转移性黑色素瘤患者MAGEA1的表达情况发现,MAGEA1在转移性黑色素瘤患者(n=359)的MAGEA1 RNA表达水平远高于原位黑色素瘤患者(n=102)(图1A),差异有统计学意义(P<0.001),表明MAGEA1可能与黑色素瘤转移密切相关。将MAGEA1过表达A375(A375-MAGEA1)和对照细胞系(A373-con)的RNA-seq数据,进行KEGG信号通路富集分析发现,Wnt/β-catenin信号通路中FRP的表达显著下调,GSK3β表达轻度下调,c-myc、fra-1和cycD表达显著上调,TCF/LEF和c-Jun表达轻度上调(图1B)。因此,推测MAGEA1可能通过影响Wnt/β-catenin信号通路进而引起黑色素瘤的转移。
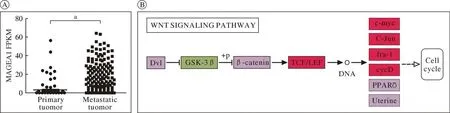
图1 MAGEA1的表达差异及对Wnt/β-catenin信号通路的影响(aP<0.05)A:TCGA公共数据中原位与转移黑色素瘤患者的MAGEA1表达;B:MAGEA1过表达A375细胞系中基因的改变情况(红色表示显著上调的基因,粉色表示轻度上调的基因,绿色表示下调的基因)Fig.1 Difference of MAGEA1 expression and its effect on Wnt/β-catenin signal pathway(aP<0.05)A: MAGEA1 expression in orthotopic and metastatic melanoma patients in TCGA public data. B: MAGEA1 overexpression gene changes in the A375 cell line (red indicates significantly up-regulated genes, pink indicates slightly up-regulated genes, and green indicates down-regulated genes)
2MAGEA1激活Wnt/β-catenin信号通路,促进黑色素瘤细胞上皮-间充质转化 将TOP flash/FOP flash质粒转染入稳定的MAGEA1过表达(A375-MAGEA1,A2058-MAGEA1)和 敲 低(A375-shMAGEA1,A2058-shMAGEA1)细胞系,并用双荧光素酶报告系统检测分析TOP与FOP荧光素酶活性的比值。过表达MAGEA1时,TOP/FOP比值增加,即β-catenin的转录活性增加(P<0.05);敲低MAGEA1,TOP/FOP比值下降(图2A),差异有统计学意义。为了进一步证实这一结果,对黑色素瘤细胞核和细胞质中β-catenin的表达进行检测,过表达MAGEA1后,细胞核中的β-catenin表达上升,细胞质中表达变化不明显(图2B)。免疫荧光方法分析A2058细胞中β-catenin的入核情况(图2C),MAGEA1过表达后β-catenin出现了明显的入核现象。而且c-myc和β-catenin随着MAGEA1的表达增高上调,而GSK-3β和p-GSK-3β表达下调(图2D),表明MAGEA1可激活Wnt/β-catenin信号通路。同时,MAGEA1的过表达可以下调E-cadherin表达,上调Vimetin和Twist1的表达(图2E)。初步验证了MAGEA1对黑素瘤细胞EMT标志物表达的影响。
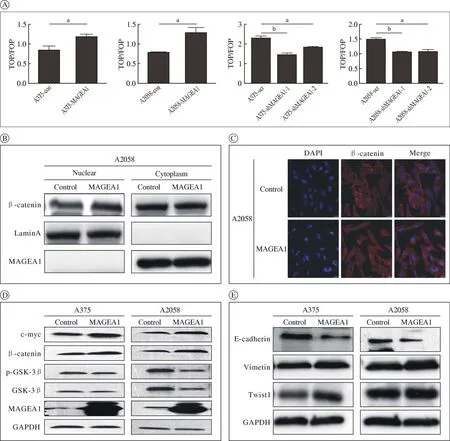
图2 MAGEA1对Wnt/β-catenin信号通路相关蛋白与EMT相关蛋白的影响(aP<0.05;bP<0.01)A:稳定过表达和敲低MAGEA1的A375和A2058细胞中TOP/TOP的活性值;B:过表达MAGEA1细胞中细胞核和细胞质中βcatenin的表达情况;C:过表达MAGEA1细胞株中免疫荧光检测β-catenin的表达情况(蓝色代表细胞核,红色代表β-catenin);D:过表达MAGEA1细胞中c-myc、β-catenin、GSK-3β和p-GSK-3β蛋白的表达情况;E:过表达MAGEA1细胞中E-cadherin、Vimetin和Twist1蛋白的表达情况Fig.2 Effect of MAGEA1 on Wnt/β-catenin signaling pathway related proteins and EMT related proteins (aP<0.05; bP<0.01)A: TOP/TOP activity values in A375 and A2058 cells stably overexpressing and knocking down MAGEA1; B: The expression of βcatenin in the nucleus and cytoplasm of cells overexpressing MAGEA1; C: Immunofluorescence detection of β-catenin expression in overexpression MAGEA1 cell line (blue represents cell nucleus, red represents β-catenin); D: Overexpression of c-myc, β-catenin, GSK-3β and p-GSK-3β protein expression in MAGEA1 cells; E: Overexpression of E-cadherin, vimetin and twist1 proteins in MAGEA1 cells
3MAGEA1通过c-Jun激活Wnt/β-catenin信号通路促进黑色素瘤的迁移和侵袭 敲低TCF7后c-Jun的RNA表达显著降低(P<0.001) (图3A),c-Jun的磷酸化水平显著降低,而c-Jun总的表达水平保持相对稳定(图3B),差异均有统计学意义,表明Wnt/β-catenin信号通路的活性确实受到抑制。迁移和侵袭结果表明,过表达MAGEA1的A375和A2058细胞株中敲低TCF7后,细胞的迁移(图3C)和侵袭(图3D)能力显著下降(P<0.05),差异有统计学意义。提示MAGEA1通过c-Jun激活Wnt/β-catenin信号通路促进黑色素瘤的迁移和侵袭。
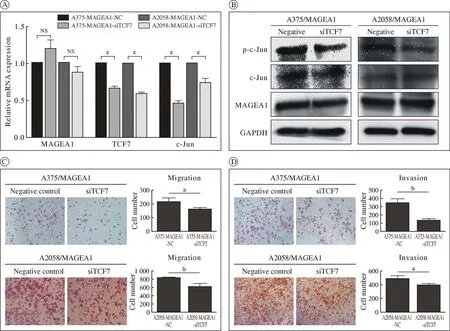
图3 MAGEA1通过c-Jun激活Wnt/β-catenin信号通路的验证以及对黑色素瘤细胞表型产生的影响(aP<0.05;bP<0.01;cP<0.001)A:转染siTCF7后对过表达MAGEA1细胞部分基因mRNA表达的影响;B:转染siTCF7后对过表达MAGEA1细胞部分基因蛋白表达的影响;C:敲低TCF7后对过表达MAGEA1细胞株迁移的影响;D:敲低TCF7后对过表达MAGEA1细胞株侵袭的影响Fig.3 MAGEA1 was used to verify the activation of Wnt/β-catenin signal pathway by c-Jun and its effect on the phenotype of melanoma cells(aP<0.05; bP<0.01; cP<0.001)A: Effect of siTCF7 transfection on mRNA expression of some genes in cells overexpressing MAGEA1; B: The effect of transfection of siTCF7 on the protein expression of some genes in cells overexpressing MAGEA1; C: The effect of knocking down TCF7 on the migration of overexpressing MAGEA1 cell lines; D: The effect of knocking down TCF7 on the invasion of overexpressing MAGEA1 cell lines
讨 论
恶性黑色素瘤是皮肤恶性程度最高的肿瘤,患者的5年生存率取决于诊断时的疾病分期,一旦出现转移,死亡率极高,因此研究恶性黑色素瘤的发病和转移机制非常重要。MAGEA1是第一个黑色素瘤抗原基因(MAGE)家族基因,迄今为止研究者已经发现至少56个MAGE家族基因[11],Caballero等[12]首先通过分析血液和组织证明黑色素瘤患者的MAGEA1家族发生了突变。更多的研究表明MAGEA1可以通过多种方式调节肿瘤的生物学行为,在乳腺癌和卵巢癌细胞中MAGEA1可与FBXW7相互作用,调节NOTCH1受体的表达而影响肿瘤细胞的增殖和迁移[8],MAGEA1也可以与SKIP蛋白和HDAC1相互作用,抑制肿瘤细胞的转录[13],MAGEA1的去甲基化可诱导非小细胞肺癌的进展[14],但MAGEA1参与黑色素瘤转移的机制尚无报道。我们对MAGEA1调节黑色素瘤恶变过程的内在机制进行了系列研究探索,并首次证明MAGEA1可以通过Wnt/β-catenin信号通路参与黑色素瘤转移过程。
Brasseur等[3]和Barrow等[15]分别证明转移黑色素瘤患者组织中MAGEA1的表达在RNA和蛋白水平均有所升高,这与我们实验和数据分析的结果一致,即转移黑色素瘤患者MAGEA1表达远远高于原位黑色素瘤患者。Wnt/β-catenin信号通路是调控细胞发育的重要级联反应之一,并且与肿瘤紧密相关。越来越多的证据表明Wnt/β-catenin信号通路可以促进肿瘤的转移[16],Wnt/β-catenin信号在肿瘤尤其是结直肠癌中发挥的作用最为突出[17-20],其他肿瘤中Wnt/β-catenin信号也可出现异常,如肺癌、肝细胞癌、乳腺癌、胃癌[21-26]等。目前已经报道多种基因可以通过Wnt/β-catenin信号通路影响肿瘤的转移,Kindlin-2通过Wnt/βcatenin信号传导促进肝细胞癌的侵袭和转移[22],DDAH1通过Wnt/β-catenin信号通路介导胃癌细胞的侵袭和转移[25],YPEL3通过Wnt/β-catenin信号通路抑制鼻咽癌细胞的上皮间质转化和转移[27]。我们发现Wnt/β-catenin信号通路在黑色素瘤转移中起着重要作用。已有的研究表明β-catenin的上调会导致黑素瘤转移增加[28-30],与我们的研究结果一致。然而,Lim和Nusse[31]则认为Wnt/βcatenin信号传导的丢失可能有助于黑素瘤的进展,β-catenin在黑素瘤中的作用有待进一步证实。黑素瘤的发生发展是多个信号通路参与的过程,Wnt/β-catenin信号通路亦可与其他信号通路发生相互作用。MAGEA1通过Wnt/β-catenin信号通路对黑素瘤细胞产生的影响是直接还是间接作用我们将继续探索。
综上所述,MAGEA1作为一种与恶性肿瘤转移相关的基因,在恶性黑色素瘤中可通过激活Wnt/β-catenin信号通路促进细胞的转移。
