沉水植物水生态修复作用及应用边界条件
2021-06-22刘凤茹雒翠张扬徐强叶运顺李红桔
刘凤茹 雒翠 张扬 徐强 叶运顺 李红桔

摘要 我國河湖水体生态系统退化严重,因此修复河湖水体,使其恢复自然状态至关重要。水生植物是水生态系统的重要组成,通过吸收富集、根系微生物作用以及化感作用等吸收、降解水中污染物,净化水体,营造良好的生态环境。沉水植物作为水生植物的一种,在水体生态修复中起着重要作用。通过系统梳理沉水植物对N、P常规污染物、重金属离子和有毒有机污染物的去除效果,国内外应用情况以及沉水植物应用于水体生态修复的边界条件,为水体生态修复中沉水植物的筛选提供参考。
关键词 水生植物;沉水植物;水生态修复;污染物;去除率;边界条件
中图分类号 X173;X52 文献标识码 A
文章编号 0517-6611(2021)09-0066-04
doi:10.3969/j.issn.0517-6611.2021.09.018
Abstract The river and lake water ecosystem in China is seriously degraded,so it is very important to restore the river and lake water body to its natural state.As an important part of the aquatic ecosystem,aquatic macrophytes absorb and degrade pollutants in water through absorption,enrichment,root microbial action and allelopathy,so as to purify the water body and create a good ecological environment.As a kind of aquatic macrophytes,submerged macrophytes play an important role in the ecological restoration of water bodies.By systematically combing the removal effects of submerged macrophytes on conventional pollutants,heavy metals and toxic organic pollutants,domestic and foreign applications,and the boundary conditions of submerged macrophytes used in ecological restoration of water bodies,the article provides the references for the selection of submerged macrophytes in ecological restoration of water bodies.
Key words Aquatic macrophytes;Submerged macrophytes;Water ecological restoration;Pollutants;Removal rate;Boundary conditions
我国河湖水体众多,随着社会经济的快速发展,城市化进程不断加快,城市水体污染日益严重,水生态系统严重退化。因此,在控制污染物入河的前提下,对水体进行生态修复,使其恢复自然状态显得尤为重要。水生植物是水体生态系统的重要组成部分,在水体生态修复中起着极其重要的作用。水生植物通过其枝叶和根系吸收污染物,抑制藻类生长,通过根系微生物对污染物质进行降解、转化、分解,去除水体污染物质,同时利用光合作用释放氧气,营造良好的生态环境。沉水植物作为水生植物的一种,是水生态环境的重要组成部分。由于全株生长在水中,生长过程主要从水中吸收营养物质,因此是水环境中重要的污染净化体[1]。黄永杰等[2]研究表明,浮萍、香蒲、水鳖等水生植物对Cu、Pb、Zn等重金属均有很好的富集作用。学者们通过共培养试验已经证实狐尾藻、金鱼藻、眼子菜、黑藻等沉水植物可以通过化感作用抑制某些藻类的生长[3-6]。芽孢杆菌能将有机磷、不溶解磷降解为无机的、可溶的磷酸盐,从而被植物直接吸收利用[7]。
利用沉水植物进行水体生态修复尽管已有众多研究,但由于不同沉水植物对营养盐的耐受程度不同,不同沉水植物适宜种植水深、水流、风浪等条件不同,适宜的生长环境不同,应用边界条件不清晰、管养不到位等,导致现在的水体生态修复效果不佳。笔者通过总结不同沉水植物对不同污染物的去除效果,明确其应用边界,为水体生态修复中沉水植物的筛选提供参考。
1 污染物的去除效果
不同类型水生植物对水体生态修复的作用不同,如挺水植物对藻类的抑制作用使藻类光合作用减弱,沉水植物光合作用对水化学性质具有重要影响[8]。同一类型水生植物对不同浓度的污染水体其净化效果也不同。
1.1 对N、P常规污染物的去除效果
沉水植物对N、P污染物的去除效果研究较多,由于研究条件不一,得出的污染物去除率不尽相同,但不同研究者得出的沉水植物污染物去除能力强弱基本一致。
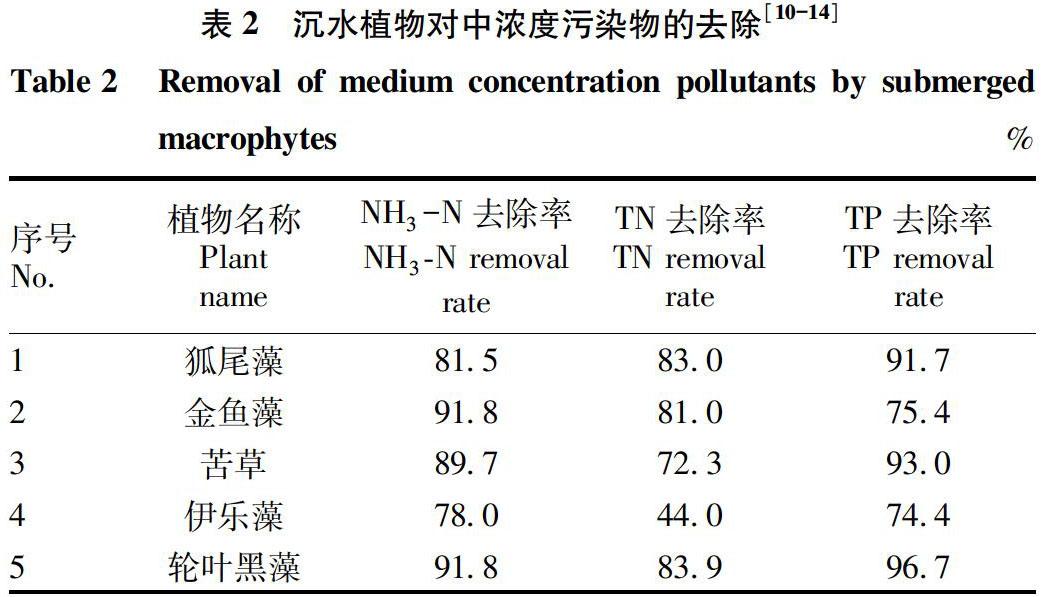
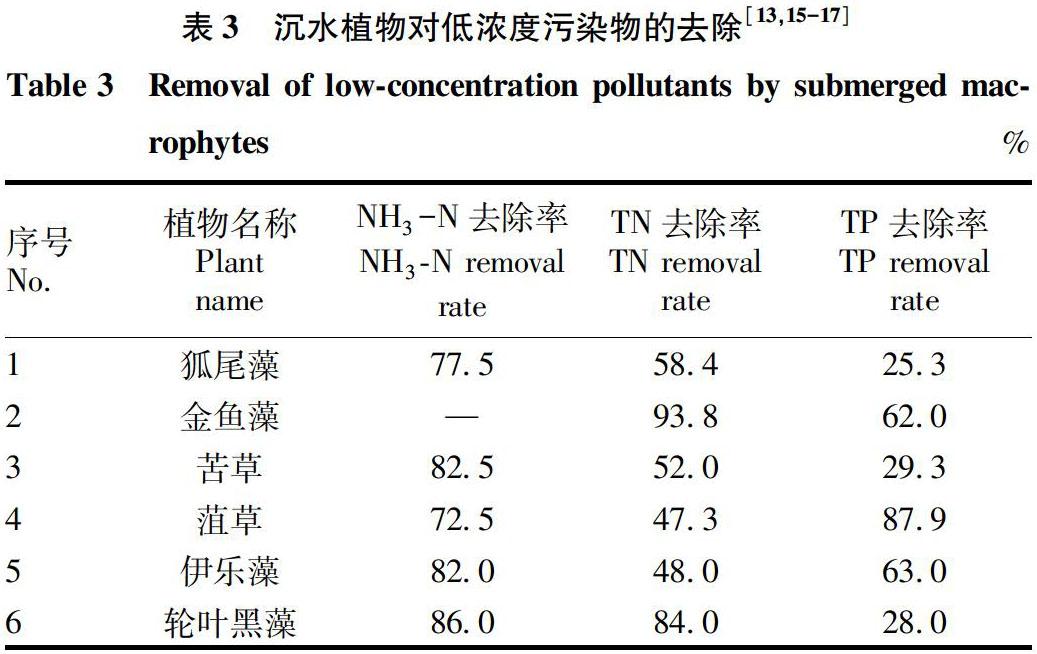
表1~3通过总结常见沉水植物对不同浓度N、P污染物的去除效果,为水体生态修复中沉水植物的选择提供参考。表中高、中、低浓度污染物的划分参考《城市黑臭水体污染程度分级标准》以及地表水环境质量标准(GB 3838—2002),定义高浓度污染物范围为NH3-N>8 mg/L、TN>8 mg/L、TP>3.0 mg/L;中浓度污染物范围为NH3-N=2~8 mg/L、TN=2~8 mg/L、TP=0.4~3.0 mg/L;低浓度污染物范围为NH3-N<2 mg/L、TN<2 mg/L、TP<0.4 mg/L。
由表1可知,金鱼藻对高浓度NH3-N的去除率最高,达93.5%,其次为伊乐藻、苦草,去除率均在90.0%以上;伊乐藻对高浓度TN的去除率最高,达90.1%,其次依次为苦草、金鱼藻、菹草、狐尾藻、轮叶黑藻;金鱼藻对高浓度TP的去除率最高,达96.6%,其次为伊乐藻和苦草,去除率在90.0%以上,菹草和轮叶黑藻对TP的去除率较低,在90.0%以下。由此可知,金鱼藻对高浓度NH3-N和TP污染水体的净化效果较好,伊乐藻对高浓度TN污染水体的净化效果较好。
由表2可知,金鱼藻和轮叶黑藻对中浓度NH3-N的去除率较高,均达91.8%,其次为苦草,去除率为89.7%,相比之下狐尾藻和伊乐藻对中浓度NH3-N的去除率较差;对中浓度TN的去除率从大到小依次为轮叶黑藻>狐尾藻>金鱼藻>苦草>伊乐藻,轮叶黑藻对TN的去除率最高,达83.9%,伊乐藻对TN的去除率最低,仅为44.0%;对中浓度TP的去除率从大到小依次为轮叶黑藻>苦草>狐尾藻>金鱼藻>伊乐藻,轮叶黑藻对TP的去除率最高,达96.7%,去除率最低的为伊乐藻,仅为74.4%。由此可知,轮叶黑藻对中浓度污染水体的净化效果最好。
由表3可知,对于低浓度NH3-N的去除效果为轮叶黑藻>苦草>伊乐藻>狐尾藻>菹草;低浓度TN的去除效果为金鱼藻>轮叶黑藻>狐尾藻>苦草>伊乐藻>菹草;低浓度TP的去除效果为菹草>伊乐藻>金鱼藻>苦草>轮叶黑藻>狐尾藻。由此可知,轮叶黑藻对低浓度NH3-N和TN的去除效果较好,菹草对低浓度TP的去除效果较好。
以上数据表明,同一沉水植物对不同浓度污染物的去除效率不同,如狐尾藻,在表1中NH3-N浓度>8 mg/L时,去除率为89.9%;在表2中NH3-N浓度为2~8 mg/L时,去除率为81.5%;在表3中NH3-N<2 mg/L时,去除率为77.5%。这是因为植物对水中污染物的浓度有一定的耐受范围,在该范围内,去除率随着浓度的升高而增加,马剑敏等[18]也得出相似的结论。因此,在实际应用中,需根据水体污染程度及污染类型选择合适的植物进行水体生态修复。
1.2 对重金属离子的去除效果
沉水植物可吸收重金属离子,其体内的谷胱甘肽、植物螯合素、金属硫蛋白、有机酸或氨基酸等含巯基物质可降低重金属离子的毒性[19],通过络合及区域化作用,使重金属离子在植物体内留存并富集,从而降低其对环境的污染[20]。Keskinkan等[21]研究了金鱼藻对Pb、Zn和Cu的富集能力,结果表明金鱼藻对Cu(Ⅱ)、Zn(Ⅱ)、Pb(Ⅱ)的最大富集能力分别为6.17、13.98、44.80 mg/g。Bunluesin等[22]研究表明黑藻对Cd具有很好的吸附能力,能在20 min达到吸附平衡,最大吸附能力为15.0 mg/g。Sivaci等[23]研究发现菹草对不同浓度(0、4、8、16、32 和 64 mg/L)的Cd溶液的吸收效果均很好,而狐尾藻仅对较低浓度(0、4和8 mg/L)的Cd吸收效果好。Lesage等[24]研究表明穗花狐尾藻对Co、Ni和Zn的富集能力分别为2.3、3.0和6.8 mg/g,对Cu的吸收能力为29.0 mg/g。陈国梁[25]研究了沉水植物对As的富集效果,富集能力从强到弱依次为苦草(1.80 mg/g)>黑藻(1.71 mg/g)>菹草(1.66 mg/g)>狐尾藻(1.64 mg/g)>金鱼藻(1.10 mg/g)。冉艳淋等[26]研究了4种常见沉水植物(苦草、狐尾藻、金鱼藻、黑藻)对As的富集特征,发现沉水植物对As的富集能力与水环境中As的浓度、形态、共存离子如磷酸盐、Zn离子等因素有关,其对As的耐受性及富集能力可以通过合适的化学、生物方法增强,从而提高沉水植物对含As废水的净化能力。另有学者研究了沉水植物活体及粉对重金属的修复效果,结果表明沉水植物活体对重金属的去除能力大于粉[27]。表4列举了几种沉水植物对重金属的富集系数。
由表4可知,沉水植物对水体中重金属的富集系数远远大于对沉积物中重金属的富集系数。不同沉水植物对同一重金属的富集能力不同,同一沉水植物对不同重金属的富集能力也不同。高海荣等[1]研究表明菹草对Pb、Cd的富集作用较强;黑藻对Pb、Cu、Zn均有较高的富集,而对Cd的富集能力却较弱;穗花狐尾藻对Zn的富集能力最强;小茨藻对Cd的富集最强;龙须眼子菜对Zn、Cu有较大的富集,而对Pb、Cd的富集相对较弱。谢佩君等[28]研究表明黑藻对Cu的生物富集系数(BSAF)是苦草的2.9倍、金鱼藻的2.7倍,对Pb的富集能力从强到弱依次为苦草>黑藻>金鱼藻。晏丽蓉[29]研究表明沉水植物对Cd、Pb、Zn的富集能力从强到弱依次为苦草>黑藻>金鱼藻,这与乔云蕾等[20]的研究结果(苦草中Cd的富集量是黑藻的1.90倍,是金鱼藻的3.02倍)一致。综上可知,狐尾藻、黑藻、菹草、小茨藻和龙须眼子菜对Zn的富集能力最强,金鱼藻对Cu和Cd的富集能力最强,苦草对Cd的富集能力最强。在实际应用中,需根据具体的重金属类型及水体状况选择合适的沉水植物净化污染水体。
1.3 对有毒有机污染物的去除效果
沉水植物除了对传统N、P污染物和重金属离子有较好的去除效果外,对酚类、苯类、有机氯类等有毒有机污染物也有一定的去除效果,利用沉水植物的这种能力,可有效对受有毒有机污染物污染的水体进行修复。
Zhang等[30]研究了几种沉水植物对烷基酚的吸收效果,结果表明,狐尾藻对壬基酚的积累能力最高,达3 050 μg/kg,伊乐藻(1 490 μg/kg)、金鱼藻(1 160 μg/kg)次之,菹草对壬基酚的积累能力最低,仅为854 μg/kg;辛基酚在沉水植物中的积累量与壬基酚不同,伊乐藻中积累量最高,达3 630 μg/kg,其次为狐尾藻(3 090 μg/kg),菹草和金鱼藻对辛基酚的积累量较少,分别为1 210、1 140 μg/kg。Heinis等[31]研究了轮藻对4-壬基苯酚(NP)的吸收,结果发现,在30和300 μg/L的NP溶液中,轮藻的最大吸收量分别为11.5和139.0 mg/kg,降解50%的NP需要10.3 d,降解95%的NP需要189 d;在30和300 mg/L NP溶液中暴露20~48 d,轮藻累计的最大干重NP浓度分别為2.7和27.4 mg/kg。
于方磊[32]采用正交设计法研究3种水生植物普生轮藻、豆瓣菜和菹草对水体中苯的净化作用,结果表明在温度为18 ℃、处理时间为4 h、生物量为3 g时,普生轮藻对苯的去除率最高,达35.26%;温度为18 ℃、处理时间为4 h、生物量为1 g时,豆瓣菜对苯的去除率最高,达69.71%;温度为8 ℃、处理时间为4 h、生物量为2 g时,菹草对苯的去除率最高,达55.45%。王燕[33]研究了轮叶黑藻、伊乐藻、金鱼藻3种沉水植物对二甲戊灵的去除效果,结果表明当二甲戊灵浓度为0.1 mg/L时,去除率最高,去除率从大到小依次为轮叶黑藻(89.53%)>伊乐藻(84.28%)>金鱼藻(83.96%)。
Garrison等[34]研究了伊乐藻对p,p′-DDT及其异构体o,p′-DDT的降解,结果发现其降解半衰期为1~3 d,使用伊乐藻对碳14标记的p,p′-DDT进行的降解试验表明,高达22%的DDT类似物与植物共价结合。从以上研究可知,沉水植物对多种有毒有机污染物均有一定的富集能力,在提供一定景观效果的同时也可有效对污染水体进行修复。
2 沉水植物水生态修复国内外应用情况
由于沉水植物对水体污染物具有良好的去除效果,在水体生态修复中得到越来越多的应用。
2.1 国内应用情况
杭州西湖茅家埠湖区采用苦草、狐尾藻、黑藻、金鱼藻、微齿眼子菜和篦齿眼子菜6种沉水植物进行水体生态修复,沉水植物的平均盖度达36.8%,苦草、金鱼藻、狐尾藻生长扩散成为优势种,其平均盖度分别为56%、66%、71%,水质得到一定改善,TN、NO3-N呈下降趋势[35]。沉水植物为山东省南四湖(南阳湖、独山湖、昭阳湖、微山湖)敞水区的主要水生植物,水生植物物种数、生物多样性指数、生物量与水体透明度表现出正相关(P<0.05)[36]。深圳市龙岗区龙岗中心城龙潭公园采用狐尾藻、苦草、轮叶黑藻等沉水植物修复公园水体,效果显著,修复后水体水质达地表Ⅳ类,目前已稳定运行一年多。云南滇池、异龙湖[37]、河北白洋淀[38]、深圳荔枝湖、香蜜湖等水体均采用沉水植物对水体进行生态修复,取得了良好的效果。
2.2 国外应用情况
德国49个北部温带浅水湖泊采用沉水植物进行生态修复,建立生态模型,并检验沉水植物对浅水湖泊水体生态修复的作用,结果表明通过种植沉水植物,湖泊水体在春季和夏季均能保持良好的水质,不会出现富营养化[39]。在西班牙地中海沿岸的Albufera deValència自然公园浅水湖泊中种植狐尾藻、金鱼藻、菹草等沉水植物,成功恢复了水体生态环境[40]。沉水植物穗状狐尾藻和黑藻是韩国利用最多的2种沉水植物,在众多水体中均有应用[41]。Havens等[42]通过探索沉水植物在不同水位条件下的恢复策略,成功恢复了美国佛罗里达州的一个大型亚热带湖泊中的沉水植物。
3 沉水植物水生态修复应用边界条件
沉水植物作为水体生态修复的重要环境因子,对水体生态修复起着至关重要的作用。尽管目前已有众多应用,但由于其具有众多应用边界条件,在应用中往往由于超过其边界条件导致沉水植物大面积死亡、生态系统崩溃的现象,因此,明确沉水植物的边界条件至关重要。如营养盐浓度/污染负荷、透明度/水深、底质、鱼类和其他水生动物、藻类(浮游藻类和附着藻类)、水流/风浪等[43]。
3.1 营养盐浓度/污染负荷
不同沉水植物对营养盐的耐受浓度不同,同一沉水植物对不同浓度的同一污染物其去除效果也不同。当营养盐达到一定浓度时,沉水植物会逐渐死亡,因此污染负荷过高的水体不宜采用沉水植物進行水生态修复。如郭俊秀[44]研究表明氨氮浓度从1.5 mg/L开始诱导抗氧化酶超歧化氧化酶(SOD)、过氧化物酶(POD)活性,对轮叶黑藻的正常生长产生胁迫。颜昌宙等[45]研究表明氨氮浓度为4 mg/L开始诱导抗氧化酶SOD、POD活性,当达到16 mg/L 时,黑藻在20多天内全部死亡。菹草对氨氮的耐受范围为0~2 mg/L,当氨氮浓度超过4 mg/L时菹草生长速率明显降低,形成逆境胁迫,产生氨害,影响菹草的正常生理活动,抑制其正常生长[46]。王斌等[47]研究表明竹叶眼子菜比较适合寡营养和中营养水质条件,过高的营养条件(氨氮>2.5 mg/L,TP>0.15 mg/L)对竹叶眼子菜是一种胁迫,可抑制其生长。
3.2 透明度/水深
水体透明度对沉水植物的生长起着重要作用,因水体透明度影响真光层,而水生植物的生长需要光照。根据相关研究[48-49],只有当水体真光层深度接近水深的情况(比值>0.8),恢复沉水植物才有可能,而水体的真光层深度与透明度有关,为2.0~2.5倍水体透明度;水深越浅,水体透明度恢复越快;1.2 m左右水深的水体透明度恢复快,适宜采用沉水植物进行水体生态修复,超过1.2 m水深不适宜采用沉水植物进行水体生态修复。
3.3 其他
河湖水体沉水植被恢复成和败很大程度上取决于水生动物特别是鱼类群落结构的组成。草食性鱼类以水体中的水草为食物,大量放养草食性鱼类加速了沉水植被的衰退,水生植物种类大量减少[50-51],水体的天然净化能力大大降低,浮游植物大量繁殖,透明度明显降低[52]。
水流能增加水环境中O2、CO2以及营养物质的交换[53]。研究表明,在较低的流速范围内(0~0.01 m/s),沉水植物光合作用速率与流速成正比,但当流速超过此范围,沉水植物的光合作用受到明显抑制[54]。
除上述提到的沉水植物应用的边界条件外,应用沉水植物进行水体生态修复的最重要的前提条件是只能应用在已经控源截污的水体中。
4 展望
沉水植物对水体生态系统修复效果已进行了大量研究,应用沉水植物修复水体生态系统已经成为一种趋势,在国内外均有大量应用案例。但由于沉水植物应用于水体生态修复时有较多的边界条件,且不同沉水植物对营养盐的耐受程度、适宜种植水深以及水流速度等均不同,因此在种植前需明确待修复水体的污染物浓度、底质、透明度、水流、风浪等条件,选择合适的种类进行水体生态修复。
另外,在使用沉水植物进行水体生态修复时,需注意季节间的搭配。沉水植物冬季种较少(菹草、伊乐藻),夏季种则很多。在水体生态修复中注意冬季种与夏季种搭配使用,保证草型水生态系统的稳定延续。最后,在使用沉水植物进行水体生态修复时,需注意先锋物种的使用。轮叶黑藻、伊乐藻属于先锋物种,耐污能力较强、净化效果较好,但生长繁殖快,不易控制,在水体中容易形成优势物种,造成管养困难,且影响其他植物的生长。因此,在景观水体尤其是底泥营养物质丰富的水体生态修复中不建议使用轮叶黑藻和伊乐藻,在条件较恶劣的水体如三面光河道可适当使用,因其难以在三面光水体中快速繁殖,较好控制。
参考文献
[1] 高海荣,陈秀丽,赵爱娟,等.5种沉水植物对重金属富集能力的对比研究[J].环境保护科学,2016,42(4):101-105.
[2] 黄永杰,刘登义,王友保,等.八种水生植物对重金属富集能力的比较研究[J].生态学杂志,2006,25(5):541-545.
[3] KRNER S,NICKLISCH A.Allelopathic growth inhibition of selected phytoplankton species by submerged macrophytes[J].Journal of phycology,2002,38(5):862-871.
[4] HILT S(NEE KRNER),GHOBRIAL M G N,GROSS E M.In situ allelopathic potential of Myriophyllum verticillatum (Haloragaceae) against selected phytoplankton species[J].Journal of phycology,2006,42(6):1189-1198.
[5] MULDERIJ G,SMOLDERS A J P,VAN DONK E.Allelopathic effect of the aquatic macrophyte,Stratiotes aloides,on natural phytoplankton[J].Freshwater biology,2006,51(3):554-561.
[6] 李小路,潘慧云,徐洁,等.金鱼藻与铜绿微囊藻共生情况下的化感作用[J].环境科学学报,2008,28(11):2243-2249.
[7] 许航,陈焕壮,熊启权,等.水生植物塘脱氮除磷的效能及机理研究[J].哈尔滨建筑大学学报,1999,32(4):69-73.
[8] 高光.伊乐藻、轮叶黑藻净化养鱼污水效果试验[J].湖泊科学,1996,8(2):184-188.
[9] 张倩妮,陈永华,杨皓然,等.29种水生植物对农村生活污水净化能力研究[J].农业资源与环境学报,2019,36(3):392-402.
[10] 赵迪.刺苦草等水生植物对富营养化程度不同水体净化作用研究[D].合肥:安徽农业大学,2010:52.
[11] 金树权,周金波,包薇红,等.5种沉水植物的氮、磷吸收和水質净化能力比较[J].环境科学,2017,38(1):156-161.
[12] 赵贺芳.挺水与沉水植物对景观水体净化的研究[J].赤峰学院学报(自然科学版),2018,34(6):104-107.
[13] 童昌华,杨肖娥,濮培民.富营养化水体的水生植物净化试验研究[J].应用生态学报,2004,15(8):1447-1450.
[14] 刘晓波,高奇英,朱文君,等.苦草与金鱼藻对水体污染物的去除效果[J].给水排水,2018,54(S2):82-88.
[15] 王文林,王国祥,李强,等.菹草伊乐藻群落对富营养化水体水质的净化效果[J].南京师大学报(自然科学版),2006,29(4):111-116.
[16] 赵贺芳.漂浮植物与挺水植物对景观水体的研究[J].赤峰学院学报(自然科学版),2018,34(7):36-38.
[17] 王丽卿,李燕,张瑞雷.6种沉水植物系统对淀山湖水质净化效果的研究[J].农业环境科学学报,2008,27(3):1134-1139.
[18] 马剑敏,靳同霞,李今,等.伊乐藻、苦草和菹草对磷急性胁迫的响应[J].水生生物学报,2008,32(3):408-412.
[19] 李文学,陈同斌.超富集植物吸收富集重金属的生理和分子生物学机制[J].应用生态学报,2003,14(4):627-631.
[20] 乔云蕾,李铭红,谢佩君,等.沉水植物对受重金属镉、锌污染的水体底泥的修复效果[J].浙江大学学报(理学版),2016,43(5):601-609.
[21] KESKINKAN O,GOKSU M Z L,BASIBUYUK M,et al.Heavy metal adsorption properties of a submerged aquatic plant (Ceratophyllum demersum)[J].Bioresource technology,2004,92(2):197-200.
[22] BUNLUESIN S,KRUATRACHUE M,POKETHITIYOOK P,et al.Batch and continuous packed column studies of cadmium biosorption by Hydrilla verticillata biomass[J].Journal of bioscience and bioengineering,2007,103(6):509-513.
[23] SIVACI A,ELMAS E,GM F,et al.Removal of cadmium by Myriophyllum heterophyllum Michx.and Potamogeton crispus L.and its effect on pigments and total phenolic compounds[J].Archives of environmental contamination and toxicology,2008,54(4):612-618.
[24] LESAGE E,MUNDIA C,ROUSSEAU D P L,et al.Sorption of Co,Cu,Ni and Zn from industrial effluents by the submerged aquatic macrophyte Myriophyllum spicatum L.[J].Ecological engineering,2007,30(4):320-325.
[25] 陈国梁.沉水植物对砷的富集特征及机理研究[D].杭州:浙江大學,2014:143.
[26] 冉艳淋,陈国梁.几种常见沉水植物对砷富集的研究进展[J].生物工程学报,2020,36(3):407-415.
[27] 张饮江,易冕,王聪,等.3种沉水植物对水体重金属镉去除效果的实验研究[J].上海海洋大学学报,2012,21(5):784-793.
[28] 谢佩君,李铭红,晏丽蓉,等.三种沉水植物对Cu、Pb复合污染底泥的修复效果[J].农业环境科学学报,2016,35(4):757-763.
[29] 晏丽蓉.几种沉水植物对底泥中镉、铜、铅、锌修复作用的研究[D].金华:浙江师范大学,2013:59.
[30] ZHANG Z,WU Z B,HE L.The accumulation of alkylphenols in submersed plants in spring in urban lake,China[J].Chemosphere,2008,73(5):859-863.
[31] HEINIS L J,KNUTH M L,LIBER K,et al.Persistence and distribution of 4-nonylphenol following repeated application to littoral enclosures[J].Environmental toxicology and chemistry,1999,18(3):363-375.
[32] 于方磊.三种水生植物对苯的净化作用研究[D].太原:山西大学,2012.
[33] 王燕.沉水植物对二种除草剂耐受性及二甲戊灵去除研究[D].苏州:苏州大学,2018:91.
[34] GARRISON A W,NZENGUNG V A,AVANTS J K,et al.Phytodegradation of p,p′-DDT and the Enantiomers of o,p′-DDT[J].Environmental science & technology,2000,34(9):1663-1670.
[35] 蔺庆伟.杭州西湖西进水域沉水植物斑块镶嵌格局优化及稳定化研究[D].新乡:河南师范大学,2015:99.
[36] 客涵.南四湖沉水、浮叶植物群落结构与水环境因子相关研究[D].济南:山东大学,2014:70.
[37] 赵风斌.异龙湖沉水植物群落恢复影响因子与恢复技术研究[D].上海:上海海洋大学,2017:130.
[38] 徐杰,何萍,王钦,等.夏季白洋淀沉水植物分布与水环境因子的关系[J].湿地科学,2013,11(4):488-494.
[39] HILT S,ALIRANGUES NUEZ M M,BAKKER E S,et al.Response of submerged macrophyte communities to external and internal restoration measures in north temperate shallow lakes[J].Frontiers in plant science,2018,9:1-24.
[40] RODRIGO M A,ROJO C,ALONSO-GUILLN J L,et al.Restoration of two small Mediterranean lagoons:The dynamics of submerged macrophytes and factors that affect the success of revegetation[J].Ecological engineering,2013,54:1-15.
[41] SON D,CHO K H,LEE E J.The potential habitats of two submerged macrophytes,Myriophyllum spicatum and Hydrilla verticillata in the river ecosystems,South Korea[J].Knowledge & management of aquatic ecosystems,2017,418:1-11.
[42] HAVENS K E,SHARFSTEIN B,BRADY M A,et al.Recovery of submerged plants from high water stress in a large subtropical lake in Florida,USA[J].Aquatic botany,2004,78(1):67-82.
[43] 谭凯婷,柳君侠,王志红,等.沉水植物修复富营养化景观水体的研究进展[J].水处理技术,2019,45(6):15-18,27.
[44] 郭俊秀.营养盐对沉水植物生长指标和抗氧化酶系统的影响[D].呼和浩特:内蒙古农业大学,2008:60.
[45] 颜昌宙,曾阿妍,金相燦,等.不同浓度氨氮对轮叶黑藻的生理影响[J].生态学报,2007,27(3):1050-1055.
[46] 纪庆亮.三种水生植物氨氮耐受性和冬季净水效果研究[D].南京:南京林业大学,2010:71.
[47] 王斌,李伟.不同N、P浓度条件下竹叶眼子菜的生理反应[J].生态学报,2002,22(10):1616-1621.
[48] 秦伯强,张运林,高光,等.湖泊生态恢复的关键因子分析[J].地理科学进展,2014,33(7):918-924.
[49] ZHANG Y L,ZHANG B,MA R H,et al.Optically active substances and their contributions to the underwater light climate in Lake Taihu,a large shallow lake in China[J].Fundamental and applied limnology/Archiv für hydrobiologie,2007,170(1):11-19.
[50] 苏胜齐,姚维志.沉水植物与环境关系评述[J].农业环境科学学报,2002,21(6):570-573.
[51] 邱东茹,吴振斌,况琪军,等.不同生活型大型植物对浮游植物群落的影响[J].生态学杂志,1998,17(6):22-27.
[52] 陈洪达.养鱼对武汉东湖生态系的影响[J].水生生物学报,1989,13(4):359-368.
[53] 吴振斌.水生植物与水体生态修复[M].北京: 科学出版社,2011:418.
[54] WESTLAKE D F.Some effects of low-velocity currents on the metabolism of aquatic macrophytes[J].J Exp Bot,1967,18(2):187-205.
