刺参腐皮综合征发生的分子调控机制研究进展
2021-06-21李成华
李成华
(1.宁波大学 教育部应用海洋生物技术重点实验室, 浙江 宁波 315832;2.青岛海洋科学技术国家实验室海洋渔业科学与食品产出功能实验室,山东 青岛266071)
海参为无脊椎动物,隶属于棘皮动物门海参纲,具有六亿多年的历史,全世界约有1 200余种,其中印度洋、西太平洋海区是世界上海参种类最多、资源量最大的区域。刺参ApostichopusjaponicusSelenka(1867)作为海参纲的主要经济养殖品种,位列世界八大珍品之一,也是中国主要消费的海参品种,具有悠久的饮食历史和文化。
刺参重要的经济和文化价值极大地推进了其养殖业的发展,20世纪90年代,山东和辽宁沿海地区出现了刺参的规模化养殖;2011年开始,刺参养殖开始逐渐向福建和浙江拓展,成为刺参规模化养殖的新兴地区。目前,中国刺参养殖区主要集中在辽宁的大连、盘锦、锦州、葫芦岛等,山东的烟台、青岛等,福建的霞浦等。根据2020年中国渔业年鉴统计显示,2019年中国刺参养殖产量为1.717×105t,苗种产量为5.25×1010头,经济产值达到300亿元,已成为中国海水养殖业的重要支柱产业[1]。不仅如此,还开拓了以新加坡、中国香港、韩国、日本、美国、菲律宾、印度尼西亚等国家和地区为中心的贸易和消费市场。然而,伴随着养殖规模和密度的扩大,病害问题也日益凸显,其中首当其冲的就是刺参腐皮综合征,每年给刺参养殖业造成的经济损失高达30~40亿元,逐渐成为刺参产业健康发展的瓶颈,对生态环境和食品安全也造成巨大威胁。因此,构建绿色健康的刺参疾病防控策略意义重大。本文中,依托作者团队十余年的研究进展,综述了刺参免疫防御途径和代表性病原致病机制研究进展,同时衔接环境因子对宿主和病原的调控作用,旨在为刺参病害绿色防控和未来研究提供指导和帮助。
1 刺参腐皮综合征流行病学调查及病原鉴定
中国水产科学研究院黄海水产研究所从2002年首次开始针对山东地区养殖刺参疾病进行了流行病学、病原学和防治策略的研究,到目前为止,已对刺参养殖各阶段的易发病有了较为系统的理解。
1.1 流行特征
刺参腐皮综合征(skin ulcer syndrome,SUS)是当前对刺参养殖产业危害最为严重的疾病,又称化皮病、烂皮病等。该病于2003年2月在山东省荣成首次发生,自2004年开始在全国范围内大规模暴发,波及中国从北方到南方的主要刺参产区。
流行病学调查发现,该病多发生于每年水体温度较低的1—4月份(此期水温一般在 8 ℃以下),1—3月份为该病的高峰期[2-3],发病快且涉及养殖区广,一旦发病很快就会蔓延全池,死亡率高达90%以上,属急性死亡。
1.2 病症
该病的发病症状一般分3个阶段,发病初期刺参厌食并伴有“摇头”现象,口部出现局部感染并肿胀,口围膜松弛,触手黑浊,对外界刺激反应迟钝,活动能力和附着力变弱;中期感染的刺参身体收缩、僵直,口部出现小面积溃疡并形成白色斑点,肉刺发白,此时大部分刺参出现排脏现象;感染后期随着病情的加重,刺参体表溃烂部位增多,出现大面积溃烂,最后死亡,并自溶为“鼻涕状”的胶体,附着物上留有一白色印痕(图1)[3-4]。

A—健康刺参;B、C—腐皮综合征I期;D、E—腐皮综合征II期;F、G—腐皮综合征III期;BW—体壁;N—正常组织。
1.3 病原
目前,已经鉴定的刺参腐皮综合征病原主要有以灿烂弧菌Vibriosplendidus为典型代表的弧菌Vibriosp.,以及假交替单胞菌属细菌Pseudoalteromonasnigrifaciens和嗜水气单胞菌Aeromonashydrophlia等。2006年,王印庚等[6]对养殖刺参保苗期的腐皮综合征发病幼参进行了详细的病原学分析,发现假交替单胞菌属细菌和另外一种弧菌属细菌是优势菌。同年,张春云等[3]从患腐皮综合征刺参的病灶部位分离得到灿烂弧菌KL-1,经人工回感染试验,发现该优势菌株引发刺参发病的症状与自然发病刺参的症状相同,其对健康刺参有较强致病和致死作用,并从人工感染发病的刺参个体内分离到大量形态单一的菌株,其形态特征和生化特征与KL-1菌株完全相符,表明灿烂弧菌是刺参腐皮综合征的主要致病菌。从自然发病的幼参病灶部位还分离到一种优势菌FP-1,经人工回接感染试验证实该细菌对健康幼参有致病作用,腹腔注射FP-1,能引起受试刺参半数死亡,且发病刺参出现的症状与自然发病刺参的症状相同,回接感染后再分离得到的FP-3与FP-1为同一种细菌,符合“柯赫法则”;根据普通细菌学表型特征对FP-1进行鉴定,可初步认为细菌FP-1为弧菌属细菌,但对其16S rDNA测序并结合系统发育树的分类学分析结果表明,FP-1为假单胞菌属细菌;进一步分析得知,假单胞菌属细菌具有许多与弧菌属细菌相同的生化特征,如革兰氏阴性杆菌、极生单鞭毛、氧化酶阳性和发酵葡萄糖不产气等特征,综合这两种细菌鉴定方法所得的结果最终证明了FP-1为假单胞菌属细菌。由此可见,对刺参腐皮综合征病原的种属鉴定需要结合分子生物学和生理生化测定等多种方法。另外,芽孢杆菌属细菌也出现在患腐皮综合征刺参的肠道中,他们也被认为是导致腐皮综合征发生的重要致病菌之一。
病毒亦是刺参腐皮综合征的重要病原种类。目前关于病毒引起的刺参病害报道较少,且研究不够深入,主要涉及刺参组织液或病变组织中病毒颗粒形态观察,未对病毒的核酸组成、感染机制和病毒分类进行深入研究。2005年,王品虹等[7]用电镜负染技术检测表皮溃烂及黏液增多的刺参组织提取液,观察到大量具有囊膜近似球形样的病毒粒子,完整的病毒粒子直径为80~100 nm,囊膜厚6~10 nm,核心结构直径35~45 nm,呈六边形;应用超薄切片技术对刺参的触手臂、疣足、触手顶部、背肠血管、呼吸树和肠等组织的病毒感染状况进行观察,发现该种病毒粒子大量存在于所检测的各组织内;感染细胞的超微结构表现为大量细胞器崩解形成空泡结构,并出现“髓袢样”结构等病理变化,根据观测结果,该病毒是一种无包涵体病毒。2008年,Deng等[8]通过超薄切片的电镜观察,并从出现排脏和皮肤溃烂症刺参消化道、呼吸树的细胞胞浆中分离获得直径为75~200 nm的球形病毒颗粒,但在健康的刺参中检测不到病毒颗粒。将健康刺参暴露于含有病毒颗粒的培养基中,无论是否添加细菌悬浮液,其发病症状与可分离病毒的刺参相同,且死亡率为90%~100%;形态学观察表明,病毒形态各异,表明并不是一种病毒引起的病症,且与已报道的贝类病毒具有很大差异,然而遗憾的是并未对该病毒进行分类地位的确定。2010年,Liu等[9]亦从患腐皮综合征的刺参中分离到具有双层囊膜、直径为100~250 nm的球形病毒粒子,提取该病毒的粗提物并感染健康刺参,刺参出现包括触须活动减少、背部乳头状足部衰减、肠壁周围肿胀和腹部溃疡等病症。
2 灿烂弧菌的致病机制
刺参在发育过程中的各个阶段,均易被病原微生物感染,导致刺参疾病的发生。病原菌引起宿主感染的主要毒力因子包括蛋白酶、溶血素、Ⅲ型分泌系统、黏附因子和铁摄取系统等,通过黏附、分泌胞外毒力因子和营养竞争等多个过程,达到其在易感宿主中的入侵和定植。灿烂弧菌是导致刺参腐皮综合征发生的主要病原菌之一,目前的研究表明,灿烂弧菌的黏附因子和胞外产物等过程均与其毒力相关。
2.1 病原菌黏附
黏附作用对病原菌的致病力有着至关重要的作用,其具有抵抗黏液冲刷、细胞纤毛运动和肠蠕动等过程的清除作用,有利于病原菌在宿主体表和体内定居。Dai等[10-11]分别采用5-(4,6-二氯三嗪基)氨基荧光素[5-(4,6-dichlorotriazinyl) aminofluorescein,5-DTAF)]的化学标记法和绿色荧光蛋白(Green fluorescent protein,GFP)的生物标记法,对灿烂弧菌进行了荧光标记,进而对标记菌在刺参体内的分布进行了追踪(图2),结果表明,灿烂弧菌在感染初期可侵染到刺参的呼吸树、触手、肠道、肌肉和体腔液等各个组织中,随着感染时间延长至48 h和96 h,荧光标记的灿烂弧菌可在肠道和呼吸树中被检测到,而在其他组织中菌体的量极少。因此推断,呼吸树和肠道是灿烂弧菌感染刺参的主要组织器官。

图2 荧光显微镜下观察刺参呼吸树(A)、肠道(B)、体壁(C)、触手(D)和肌肉(E)中GFP标记的灿烂弧菌[11]
弧菌的黏附因子一般可分为两大类:菌毛黏附素和非菌毛黏附素,前者包括鞭毛和纤毛等,后者包含脂多糖和外膜蛋白等。Dai等[12]采用基因过表达技术研究了灿烂弧菌中FliC(鞭毛蛋白亚基)是否参与菌体黏附,结果表明,fliC基因过表达菌株的生物膜形成能力和运动能力均明显高于野生型菌株,且fliC基因的过表达导致灿烂弧菌对刺参肌肉、呼吸树和肠道组织中的黏附效率增加,而对体壁和触手的黏附率无明显影响,因此,FliC介导的黏附具有明显的组织特异性。除了使用FliC聚合而成的鞭毛丝作为黏附因子,二氢脂酰胺脱氢酶(dihydroacrylamide dehydrogenase,DLDs)是目前已知的灿烂弧菌另一黏附因子。Dai等[10]制备了DLD1和DLD2的多克隆抗体,并对菌体表面的DLD1和DLD2蛋白进行封闭,DLD1、DLD2抗体封闭后的菌体对刺参体腔细胞的黏附率分别为7.5%、12.5%,均显著低于野生型菌株的黏附率25%,封闭后的菌体对刺参各组织的黏附率亦均有所下降,对肠组织的黏附更具有特异性。
2.2 灿烂弧菌的致病因子
2.2.1 金属蛋白酶Vsm 金属蛋白酶是多种病原菌中公认的重要致病因子。灿烂弧菌中的vsm基因(VS_RS05940)编码金属蛋白酶,是灿烂弧菌中第一个被鉴定的胞外毒力因子[13-14]。研究者分别向同等刺参体腔细胞中加入70、90、100 μL纯化的重组Vsm蛋白(50 mmol/L),发现加入不同量Vsm蛋白后的细胞存活率约下降至对照组的86%、79%和72%,表明灿烂弧菌的Vsm对刺参体腔细胞具有明显的细胞毒性[15]。
2.2.2 铁离子吸收 铁是宿主和病原微生物必需的营养素。铁离子的氧化还原电位是导致铁离子产生生物毒性的主要原因,因此,细胞内铁离子的浓度和分布受到严格的控制。考虑到几乎所有病原微生物对铁的绝对需求,宿主通过限制入侵病原微生物的生物可利用性铁成为宿主先天免疫系统的重要组成部分。病原微生物在宿主体内夺取铁离子的能力及对铁离子的利用,已经成为多种病原微生物致病过程的重要组成部分。
研究者从铜绿假单胞菌培养物中经过Cu2+的亲和层析获得了对Fe3+具有高亲和力的荧光铁载体pyoverdine,体外抑制试验表明,pyoverdine(100 μL, 50 mmol/L)可显著抑制灿烂弧菌在2216E培养基中的生长;采用pyoverdine对刺参养殖海水进行预处理,可显著降低灿烂弧菌感染刺参的效率,实验室条件下浸泡感染试验显示,pyoverdine对仿刺参具有90%以上的相对保护效率,这意味着铁离子对灿烂弧菌感染刺参并导致刺参发病过程具有重要的作用[15]。进一步研究表明,在灿烂弧菌中至少存在两条铁吸收通路[16-17]。第一条通路是铁载体介导的Fe3+吸收,灿烂弧菌可以分泌异羟肟酸盐类型的铁载体,通过铁载体络合环境中的Fe3+形成铁载体- Fe3+复合物,铁载体-Fe3+复合物进而可以被锚定在细胞膜上的识别蛋白IutA受体识别,并将铁离子转运到细胞内供菌体自身生长及致病所需;第二条通路是亚铁离子吸收通路,在灿烂弧菌体内,酪氨酸代谢通路中的关键酶羟苯丙酮酸二加氧酶(hydroxyphenylpyruvate dioxygenase,Hppd)催化形成尿黑酸(homogentisic acid,HGA), 在特定条件下HGA聚合形成多聚物HGA-黑色素(HGA-melanine),HGA-黑色素具有铁离子还原能力,可在菌体外将Fe3+还原为Fe2+,再通过位于细菌周质空间的FeoA和FeoB介导的亚铁离子转运系统,将Fe2+转运至菌体内部,形成了灿烂弧菌中亚铁离子摄取途径[18](图3)。

图3 灿烂弧菌体内存在的铁吸收通路
多条铁摄取通路的存在有利于细菌抵抗不利的水体和宿主环境。具有全局性调控作用的铁离子吸收调节蛋白(ferric uptake regulator, Fur)是细菌响应环境中铁离子水平,进而调节细菌基因表达并做出响应的关键调节因子。利用实时荧光定量PCR技术检测低铁环境下灿烂弧菌中fur基因的表达,在灿烂弧菌培养环境下加入铁离子螯合剂8-羟基喹啉,灿烂弧菌fur基因的表达显著降低,在RNA水平上验证了灿烂弧菌的Fur确实具有响应环境中铁离子浓度的功能[19]。
2.2.3 溶血素 4-羟苯丙酮酸双加氧酶(4-Hydroxyphenylpyruvate dioxygenase,4-HPPD)是灿烂弧菌中研究较为详细的灿烂弧菌溶血素,也是铁离子依赖型的非血红素加氧酶,其在酪氨酸分解代谢途径中起关键作用。Liang等[20]首先测定了Vshppd的表达,在刺参体腔液的刺激下,Vshppd基因表达在60 min内显著上调60多倍;用绵羊血细胞进一步测试发现,灿烂弧菌Vshppd蛋白具有明显的溶血活性,并具有明显的剂量依赖性;MTT法测定结果显示,Vshppd蛋白(3~4 μg/μL)对刺参细胞具有明显的毒性作用,且随着蛋白质量浓度的增加,刺参体腔细胞的存活率逐渐下降;为了进一步验证Vshppd在致病过程中的作用,利用基因框内缺失原理,构建了Vshppd基因敲除的灿烂弧菌减毒菌株(MTVs),用野生型(WTVs)和MTVs灿烂弧菌分别感染刺参后,感染了野生型菌株的刺参表现出与腐皮综合征相关的相同症状,并且表皮腐烂面积大,而感染MTVs的灿烂弧菌对刺参的致死率大大降低,突变型菌株感染的刺参只在体表上偶有症状不明显的白点,同时刺参的半致死剂量(LD50)由野生型的5.129×106CFU/mL升高到突变型的2.606×1010CFU/mL,证实了灿烂弧菌Vshppd在病原胁迫中的重要致病作用。
2.2.4 分泌系统 一般认为,细菌有Ⅰ~Ⅳ型分泌系统,不同的病原菌通过特有的分泌系统将毒力因子分泌到细胞外发挥作用,从而导致宿主发病。目前已有研究表明,灿烂弧菌中含有与鞭毛同源性较高的fT3SS,其分泌的效应蛋白参与了灿烂弧菌的致病过程。Zhuang等[21]借助自杀性质粒pDM4,采用基因框内缺失突变的方法构建了灿烂弧菌T3SS ATPase突变菌株ΔvscN,结果表明,T3SS ATPase的敲除导致该菌的LD50升高,该突变体聚集到刺参体腔细胞表面的能力明显降低,且潜在效应因子Hop不能如野生型菌株中的Hop一样定位于刺参体腔细胞内,而且突变体中Hop的表达在RNA和蛋白质水平上均明显降低。这与该学者之前得出的重组的rHop蛋白对刺参体腔细胞具有明显细胞毒性的结果相一致[22]。这表明,Hop是灿烂弧菌的Ⅲ型分泌系统效应因子,也是迄今为止唯一得以鉴定的灿烂弧菌Ⅲ型分泌系统效应因子。
3 刺参应对灿烂弧菌感染的免疫调节机制
病原菌在进化过程中发展了多种策略以攻击宿主,其中最重要的一种方式就是通过毒力蛋白对宿主细胞造成损伤。与此同时,疾病的发生还与宿主的免疫防御息息相关。作为无脊椎动物的刺参只存在先天性免疫,缺乏脊椎动物中的获得性免疫。无脊椎动物主要通过一类被称为模式识别受体(pattern recognition receptors,PRRs)的免疫识别分子识别入侵病原,进而启动细胞免疫和体液免疫抵抗或清除病原[23-24]。
3.1 刺参细胞免疫
3.1.1 体腔细胞分类 国内外学者对棘皮动物体腔细胞的研究已经长达30年之久。随着研究的人员、目的、手段等不同,刺参体腔细胞多样且不断变化,其体腔细胞类型至今无统一的标准,体腔细胞和血细胞的确切关系也未见描述。Eliseikina等[25]在研究刺参和瓜参Cucumariajaponica时发现,体腔细胞可以分成祖原细胞、吞噬细胞、有空泡细胞、小(幼)桑椹细胞、桑椹细胞(Ⅰ、Ⅱ和Ⅲ型)、晶体细胞和震颤细胞。刘晓云等[26]利用电镜技术将刺参体腔细胞分为大颗粒细胞、小颗粒细胞、透明细胞和淋巴样细胞。而有关刺参血细胞的论述尚未见报道,有学者认为,体腔细胞相当于血细胞,血液类似于体腔液。李华等[27]将刺参体腔细胞分为6类,即淋巴样细胞、球形细胞、变形细胞、纺锤细胞、透明细胞和结晶细胞,其中以淋巴样细胞、球形细胞和变形细胞为主,透明细胞、纺锤细胞数量次之,结晶细胞数量最少,并依据细胞形态、运动能力、细胞内颗粒大小,将球形细胞又分为3个类型,即Ⅰ型球形细胞(颗粒小,细胞无运动能力)、Ⅱ型球形细胞(颗粒较小,细胞伸出刺状伪足)和Ⅲ型球形细胞(颗粒较大,细胞能做阿米巴运动)(图4)。

A—淋巴样细胞;B—球形细胞,光镜下球形细胞呈球形,Ⅰ型球形细胞胞核被胞质中大量的小颗粒遮住,细胞无运动能力,直径为5~10 μm(a),Ⅱ型球形细胞胞核被胞质中大量的小颗粒遮住,细胞外质伸出很多刺状伪足,活体观察犹如“太阳”状,细胞直径为7~12 μm,刺状伪足长为4~7 μm(b),Ⅲ型球形细胞,直径为6~13 μm,静止时呈球形,细胞核被许多颗粒状小球体遮住(c),活体观察细胞可做阿米巴运动,伸出钝状伪足(d);C—变形细胞;D—透明细胞;E—纺锤细胞;F—结晶细胞。
3.1.2 体腔细胞的免疫功能
1)趋化作用。趋化作用是指细胞沿浓度梯度向化学刺激物做定向移动。当病原微生物入侵机体后,受损细胞会释放某些化学物质,吸引大量的炎症细胞,并表现为局部炎症反应,此时镜下观察受损组织可见大量炎症细胞浸润现象。通过对腐皮综合征刺参体壁病理切片观察,发现其患病组织具有类似于高等动物的炎症现象,出现明显的白细胞增多、细胞浸润、组织空泡和肌肉纤维断裂等炎性组织学特点,同时在患腐皮综合征的刺参体壁中检测到了炎症标志分子NO,其含量比正常组刺参显著上调1.54倍,总NOS(TNOS)显著上调1.50倍,诱导型NOS(iNOS)显著上调1.24倍[28]。
2)细胞杀伤功能。细胞杀伤功能是刺参体腔细胞免疫功能之一,又被称为细胞毒性。吞噬细胞和桑椹细胞均可产生并释放杀菌物质来分解外来物质,如脂肪酶、过氧化物酶和丝氨酸蛋白酶等。研究发现,天然状态的刺参体腔液在体外状态下处理灿烂弧菌有明显的杀伤作用,而热处理过的刺参体腔液在体外状态下处理灿烂弧菌杀伤能力明显减弱,并且这种效果随着灿烂弧菌浓度的升高而减弱,表明刺参体腔细胞的杀伤功能可能是通过蛋白酶活性来实现的[15]。
3)吞噬功能。细胞吞噬作用是刺参体腔细胞免疫的主要防御机制,体腔液中的吞噬细胞能够有效地识别并吞噬外源粒子,然后将其降解或直接排出体外。大多数免疫反应都是由具有吞噬作用的变形细胞介导的。2016年,Wang等[29]将整合素抗体与刺参体腔细胞孵育,测定了体腔细胞对荧光微球菌的吞噬能力,证明了刺参体腔细胞膜上的整合素蛋白具有促进细胞吞噬的功能。2019年,Yue等[30]鉴定了雷帕霉素靶蛋白复合物2型(mechanical target of rapamycin complex 2,mTORC2)蛋白亚基Rictor对刺参体腔细胞内吞具有正调控作用。为进一步明确mTORC2介导的细胞内吞机制,2021年,Lv等[31]进而检测了Rictor被干扰后,调节内吞作用的AGC激酶家族成员SGK1、PKCα和AktSer473位点的表达水平,结果显示,Rictor被干扰抑制后,SGK1和PKCα的表达水平及AktSer473位点的磷酸化水平均受到显著抑制,而Akt含量未发生变化,这表明,mTOR抑制剂雷帕霉素处理则不能影响SGK1和Akt的表达,但能够显著抑制PKCα的表达水平及AktSer473位点的磷酸化水平。上述结果揭示了Akt的磷酸化修饰可能参与mTORC2介导的刺参体腔细胞内吞,而Rictor可能独立于mTORC2调节SGK1。
4)细胞凋亡。细胞凋亡是细胞为维持内环境稳定,由基因调控的、自主有序的生理性死亡。目前,在刺参中已扩增得到多个细胞凋亡相关基因。2016年,Shao等[32]通过RACE技术扩增得到了4个刺参caspases同源基因,即caspase-2、caspase-3、caspase-6和caspase-8,qPCR分析发现,这4个caspases基因在刺参各个组织中的分布特征及灿烂弧菌诱导后的表达水平均差异显著,表明不同的caspases基因在免疫反应中发挥不同功能。随着研究的深入,发现刺参可通过多种途径诱导其体腔细胞凋亡的发生以对抗灿烂弧菌的感染。
①细胞凋亡线粒体途径的调控:灿烂弧菌感染可促进刺参组织蛋白酶B活性增加,影响线粒体膜电位的变化并促进细胞色素c的释放,进而激活下游效应因子caspases-3以执行细胞凋亡[33-34];通过刺参活体体腔注射Bax(Bcl-2-associated X) siRNA并结合拯救试验,进一步证实了Bax能促进线粒体介导的细胞凋亡[35],而Bcl-2(B-cell lymphoma 2)则能抑制细胞色素c的释放进而抑制细胞凋亡[36];此外还发现,刺参亲环素A可作为NF-κB辅助因子促进其核移位,进而调控Bcl-2表达并诱导细胞凋亡发生[37]。
②细胞凋亡死亡受体途径的调控:2020年,Zhao等[38]首次阐述了刺参中存在以死亡受体介导的细胞凋亡,caspase-8作为死亡受体介导的凋亡途径中的关键启动子,其具有 FADD样DED结构域,能够与死亡受体蛋白FADD结合,形成死亡复合体,进而诱导下游caspase基因表达,诱导细胞凋亡。
5)细胞自噬。细胞自噬是指细胞通过双层膜包裹胞质物、入侵病原或损伤的细胞器、蛋白质等待降解物形成自噬体,并与溶酶体融合形成自噬溶酶体将包裹物消化降解,从而实现细胞自身的能量物质代谢和细胞器更新的胞内过程[39-40]。其作为一种维持细胞内环境稳态、实现自我更新的进化保守机制,同时也是免疫防御机制的重要组成部分,在机体免疫防御中发挥重要功能[41]。通过对刺参基因组分析发现,刺参中也存在众多自噬相关基因,且自噬标志性蛋白已在相关文献中报道[ 42],表明刺参中存在细胞自噬。但关于刺参在病原感染下的自噬完整通路,还有待深入研究。
6)细胞焦亡。细胞焦亡是一种程序性细胞死亡,具有与细胞坏死相似的形态学特征,如细胞质膜上会有孔的形成、细胞肿胀、细胞质膜破裂,随后内容物释放到细胞质外,引发组织急性炎症反应等[43]。经典的炎症小体主要通过细胞质内的模式识别受体(如NLR家族蛋白等)识别多种病原体或宿主源性的危险信号来组装,如NLRC4炎症小体,主要由细胞内的模式识别受体NLRC4、接头蛋白ACS和炎性caspase-1前体组装而成,可以激活炎性caspase-1蛋白[44]。在刺参中,存在模式识别受体NLRC4[45]或NLRP3[46],但在其基因组中未找到GSDMD蛋白和IL-1β。刺参caspase-1可促进HEK293细胞GSDMD和IL-1β的表达[47],表明刺参中可能也存在细胞焦亡,但还需要更深入的研究来进一步证实。
3.2 刺参免疫因子功能
3.2.1 Toll样受体信号通路 Toll样受体(toll-like receptor,TLR)属于固有免疫病原模式识别受体,可识别入侵的病原微生物蛋白、核酸和脂类及其在反应过程中合成的中间产物和代谢产物,如革兰氏阴性菌的脂多糖(LPS)、革兰氏阳性菌的肽聚糖和病毒的双链RNA等,这些都属于分子结构高度保守的病原相关分子模式(pathogen-associated molecular pattern,PAMP)。TLR通过对PAMP的识别,快速激活包括接头蛋白、信号复合体和转录因子复合体负责的细胞内信号级联反应,最终导致机体产生促炎细胞因子、抗炎细胞因子及趋化因子。
刺参Toll样受体成员分子已基本鉴定完成(图5),其中包括TLR受体2个成员分子(TLR3 和 Toll),TLR通路正调控蛋白3个[关键接头分子-髓样分化因子88(MyD88)、坏死因子受体相关因子6(TRAF6)和MyD88下游的激酶-IL-1R相关激酶1、4 (IL-1R-association kinase 1、4, IRAK 1、4)],以及NF-κB的2个亚基(Rel和p105)。
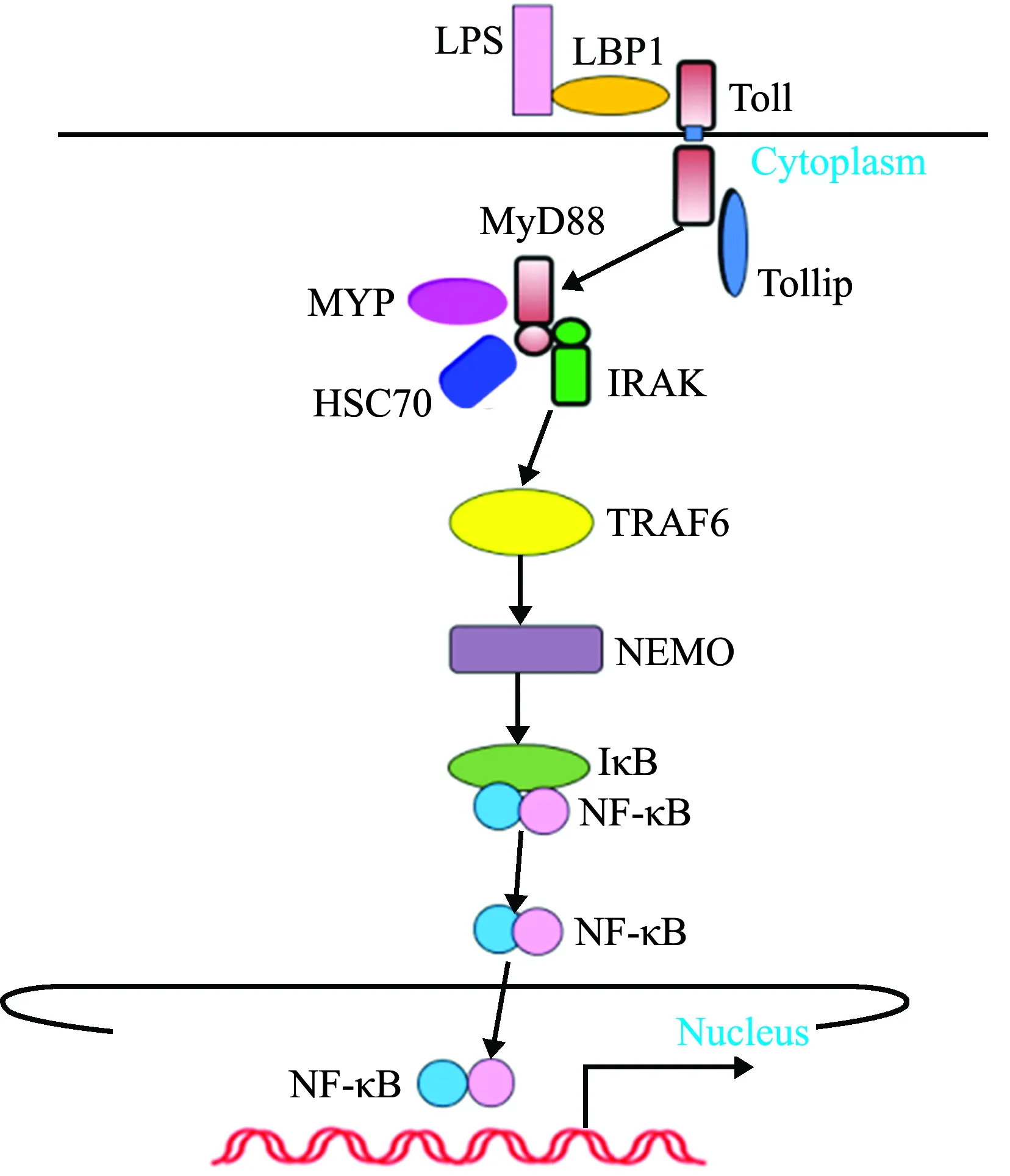
图5 刺参Toll样受体模式图
1)TLR3和Toll的免疫功能分析。2013年,Sun等[48]首次从刺参体腔细胞中克隆得到了两个TLRs基因TLR3和Toll,SMART结构预测表明,TLR3 和 Toll氨基酸序列均具有典型的TLR结构,即胞外亮氨酸重复序列LRR区、跨膜区(transmembrane, TM)和胞内TIR(toll/interleukin-1 receptor)区,说明TLR在无脊椎动物进化过程中是高度保守的;系统进化分析发现,刺参TLR3和 Toll 分别与脊椎动物类V-TLR、原口动物 P-TLRs聚为一支,表明刺参中存在两种不同类型的TLR;采用病原革兰氏阳性菌、革兰氏阴性菌、真菌和病毒类似物,如肽聚糖(peptidoglycan, PGN)、脂多糖(lipopolysaccharides, LPS)、酵母聚糖(zymosan A)和聚肌胞苷酸(polyinosinic-polycytidylic acid, Poly I:C)胁迫刺参体腔细胞,荧光定量分析表明,这两个基因的表达模式在刺参不同组织中针对不同的刺激有所不同,即在PGN刺激下,TLR3和Toll基因在体腔细胞中的表达变化最显著,在LPS和Poly I:C刺激下,两个基因在管足中表达变化最显著,在Zymosan A刺激下,两个基因在呼吸树中表达变化最显著。由此推测,刺参TLR3和Toll 在对抗细菌感染过程中发挥着重要作用且功能上可能存在差异。
2)MyD88的鉴定及功能解析。MyD88是TLR信号转导中触发先天免疫下游级联反应的关键接头分子。Lu等[49]分离并鉴定了MyD88基因全长cDNAs序列,该因子与哺乳动物、果蝇直系同源物具有非常高的结构保守性,预测的MyD88氨基酸序列中,存在一个典型的死亡域(death domain,DD)和一个保守的Toll / IL-1R(TIR)结构域;在灿烂弧菌攻毒试验中,刺参MyD88和TRAF6基因的表达水平均显著升高,但TRAF6升高的幅度更大,持续时间更长。上述结果表明,MyD88和TRAF6可能在刺参抗菌反应中起重要作用。
在不同物种中TLR信号通路的激活通常涉及不同的相互作用蛋白。Lv等[50]通过GST-pull down试验鉴定了与刺参MyD88结合的4个蛋白,分别为MYP(major yolk protein)、IRAK1、HSC70(heat shock cognate protein 70)和TIR结构域蛋白;通过免疫共沉淀试验发现,GST-HSC70和GST-MYP融合蛋白可分别与刺参MyD88结合,激光共聚焦显微镜观察显示,MyD88、HSC70和MYP均共定位于细胞质,这表明,HSC70和MYP可以与MyD88相互作用;该研究者进一步通过转染MyD88、HSC70、MYPsiRNA检测TLR信号通路的变化以探究HSC70和MYP是作为MyD88信号激活过程中所必需的辅助分子还是直接激活分子,结果显示,MyD88基因被成功干扰后,IRAK1和p105 mRNA显著降低,且MyD88蛋白表达显著降低;此外,在HSC70和MYP基因敲除后,IRAK1和p105 mRNA均显著降低;Western blot分析显示,MyD88、MYP和HSC70基因敲除48 h后,p50蛋白表达水平显著降低。这表明,MYP和HSC70是作为MyD88依赖的信号通路的辅助分子参与信号级联反应的。
3)IRAK1和IRAK4的功能解析。Lu等[51]通过RACE技术,获得了刺参IRAK1基因全长(GenBank:KJ918751),SMART分析显示,IRAK1氨基酸序列包含一个长为93个氨基酸的保守的死亡结构域和一个285个氨基酸的典型的中心激酶结构域,在激酶结构域中还包含ATP结合位点和丝氨酸/苏氨酸蛋白酶激活位点;进一步对刺参体腔细胞吞噬作用的检测发现,IRAK1基因沉默能够显著促进刺参体腔细胞对灿烂弧菌的吞噬作用。
Cui等[52]克隆了刺参IRAK4基因全长,预测IRAK4氨基酸序列中具有典型的死亡结构域(10~113 aa)和激酶结构域(160~426 aa);在灿烂弧菌攻毒和LPS胁迫下,刺参IRAK4基因表达显著上调,IRAK4基因被干扰后,TLR信号通路中下游成员IRAK1、TRAF6和p105在体内和体外试验中,均显著下调表达;在体外/体内沉默IRAK4后,刺参体腔细胞的凋亡水平分别增加了1.82倍和1.95倍。这表明,刺参IRAK4参与了Toll 样受体信号通路的级联激活应答。
4)NF-κB亚基的鉴定及功能分析。Wang等[53]鉴定了刺参中NF-κB的两个亚基Rel和p105;预测的刺参Rel蛋白具有其他物种NF-κB蛋白的保守性,在N端包含一个279个氨基酸(50~328 aa)的RHD结构域,Rel还包含两个保守的基序,其中一个为NF-κB蛋白共有序列RFRYPCEG,是所有NF-κB蛋白与DNA结合所必需的DNA识别环,NF-κB蛋白包含核定位信号(KRKR),是NF-κB蛋白核定位必不可少的结构域;预测的刺参p105蛋白在N端区域包含一个RHD同源结构域(53~348 aa)和一个带有6个锚蛋白重复序列的C端IκB样结构域和死亡结构域,p105还具有保守的DNA结合基序和核定位位点,当p105蛋白被磷酸化时,随后的26S蛋白酶体降解并激活了p50亚基活性,使得核蛋白结合位点得以暴露,促进p50的核转移。
Wang等[53]进一步测定了刺参经LPS胁迫0、10、30、60、120、240、360 min后体腔细胞中Rel、p50和p105基因的表达规律,结果显示,p105基因的下调最为显著,在10 min时与对照水平有显著差异,在胁迫60 min后恢复到基线水平;在LPS胁迫10 min后,p50的mRNA表达显著增加,达到最高水平,并在大约60 min后恢复到正常水平,这表明,p50的增加是由于刺参p105的快速降解引起的;Western blot分析表明,LPS胁迫显著降低了胞浆中p105和p50蛋白的表达水平,LPS刺激120 min后,细胞核中p50蛋白表达显著增加,LPS处理后360 min,胞浆中p105和p50蛋白水平有所恢复,但细胞核中未检测到p50蛋白,同时还发现,在未经LPS刺激的体腔细胞中,Rel蛋白主要存在于细胞质中,而在LPS胁迫的刺参体腔细胞中,Rel蛋白向细胞核转运,表达模式与p50蛋白相似。
5)NF-κB亚基的其他调控因子。Shao等[54-55]研究发现,在刺参一氧化氮合酶(NOS)、精氨酸酶(arginase)和精胺酶(agmatinase)的启动子区分别存在多个NF-κB结合位点;通过共转染鲤鱼上皮瘤细胞(endothelial progenitor cells,EPC)发现,刺参Rel是NOS的关键转录因子,序列分析表明,-375~366 bp是NF-κB的结合位点,对启动NOS转录起重要作用,而共转染arginase启动子后,荧光素酶的转录活性极显著下降,表明转录因子Rel可能负调控arginase基因的转录;此外,共转染agmatinase启动子后,荧光素酶活性无显著变化,表明转录因子Rel对agmatinase启动子的活性无影响。Jiang等[56]研究发现,刺参纤维相关蛋白FREP的表达也受转录因子Rel的调控,有趣的是,FREP基因被干扰后,Rel的mRNA表达水平显著降低[58],表明FREP可以正反馈调节Rel表达。此外,通过GST-pull down和免疫荧光技术发现,刺参亲环素A(CypA)能够与Rel互作,进一步通过干扰技术和Western blot技术分析表明,CypA作为辅助因子促进NF-κB的核移位[37]。
3.2.2 刺参JAK/STAT信号通路 目前,有关刺参JAK/STAT信号通路的研究相对缺乏,仅STAT5被报道。Shao等[57]克隆出cDNA全长为2 643 bp的刺参STAT5基因序列,预测该基因编码787个氨基酸组成的蛋白,STAT5蛋白由STAT结合结构域、STAT α结构域、STAT DNA结合结构域和SH2结构域组成;经灿烂弧菌和LPS胁迫后,刺参STAT5的mRNA表达水平显著上调,转录因子FOXP的mRNA水平也显著上调,而SOCS2的表达趋势与STAT5相反;进一步干扰STAT5的表达后,FOXP的mRNA表达水平显著下调,表明STAT5可能正调控FOXP转录,而SOCS2的mRNA表达水平则极显著上调,表明STAT5负调控SOCS2表达。关于刺参JAK/STAT信号通路的其他成员,还有待进一步研究。
3.3 刺参肠道微生物与疾病发生
肠道微生物作为生物体一个重要的“器官”,主要通过调控黏膜免疫系统的发育与成熟、维持肠道屏障的完整性、调节肠道微环境的pH、影响脂质的合成与吸收、改变氧化二甲胺与LPS的含量等途径调控宿主健康状态,介导宿主多种系统疾病的发生和发展。因此,刺参肠道菌群的组成与营养消化、生物屏障、免疫防御等生理过程密切相关,对机体健康生长具有极其重要的作用。
3.3.1 刺参肠道微生物群落组成、结构和功能 刺参肠道菌群丰富而多样,研究发现,其肠道内细菌数量为1.85×105~ 2.17×109CFU/g[58],其中大多数为异养细菌[59],以变形菌门Proteobacteria、厚壁菌门Firmicutes、疣微菌门Verrucomicrobia、拟杆菌门Bacteriodetes为主(图6),以及一定比例的梭杆菌门Fusobacteria、酸杆菌门Acidobacteria和浮霉菌门Planctomycetes等,通过生理生化的方法鉴定发现,刺参肠道组织和内容物菌群由弧菌属Vibrio、假单胞菌属Pseudomonas、节杆菌属Arthrobacter、棒杆菌属Corynebacterium、不动杆菌属Acinetobacter、微球菌属Micrococcus、奈瑟氏球菌属Neisseria、黄杆菌属Flavobacterium、黄单胞菌属Xanthomonas、柄杆菌属Caulobacter和产碱菌属Alcaligenes等细菌组成,弧菌属和假单胞菌属为肠道中的优势菌群[60]。
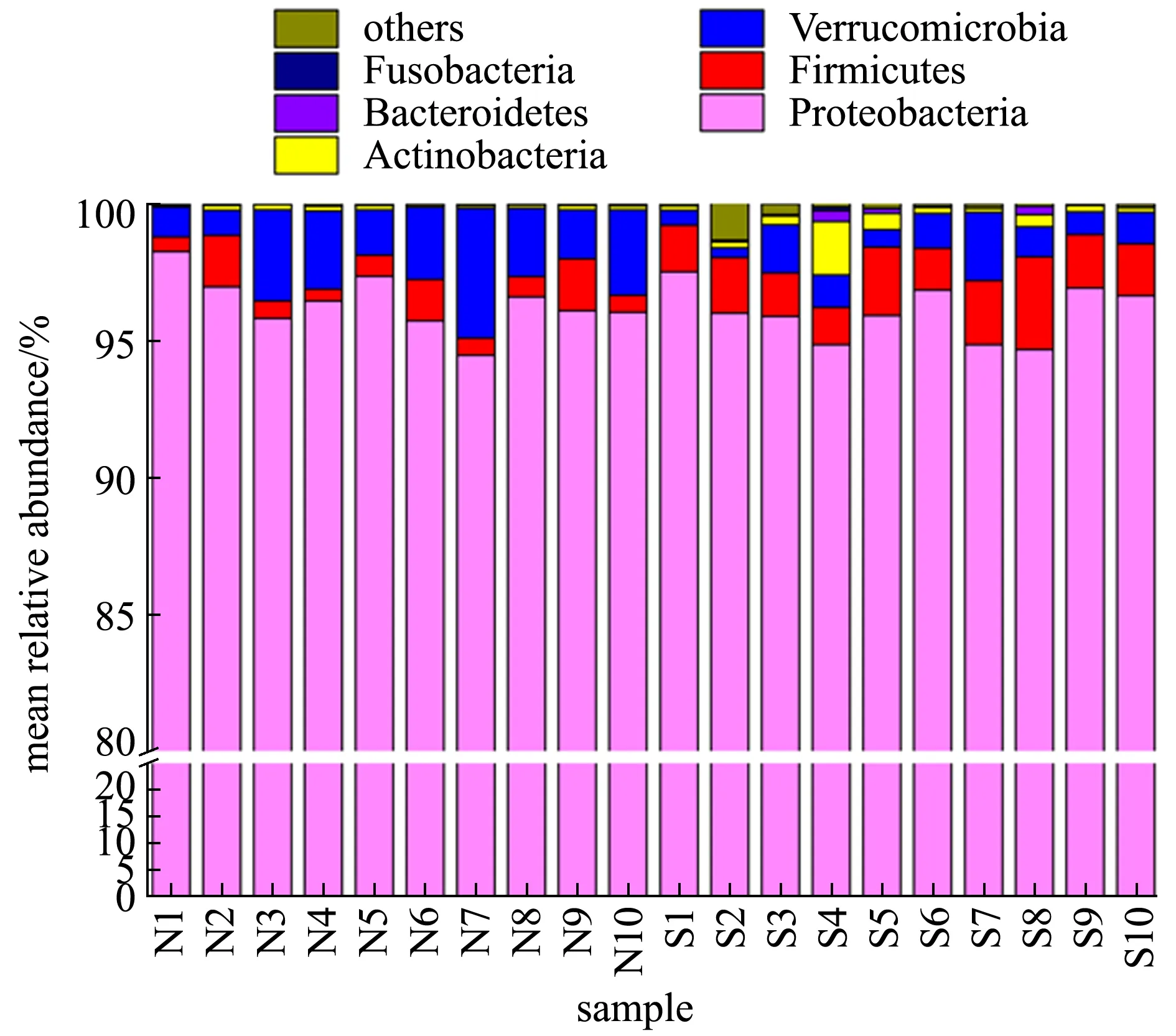
图6 健康和自然发生腐皮综合征状态下刺参肠道菌群群落组成[60]
对于刺参肠道中的可培养细菌,张文姬等[61]通过限制性内切酶片段长度多态性(RFLP)技术分析了2216E、硫代硫酸盐-柠檬酸盐-胆盐-蔗糖琼脂(TCBS)和葡萄糖琼脂培养基上分离的刺参肠道可培养菌,共获得了假单胞菌属、弧菌属、芽孢杆菌属Bacillus、希瓦氏菌属Shewanella、交替假单胞菌属Pseudoalteromonas、不动杆菌属、气球菌属Aerococcus、发光菌属Photobacterium、葡萄球菌属Staphylococcus和克锡勒氏菌属Kushneria等细菌,并证明假单胞菌属为优势菌属。对冬季刺参肠道菌群进行平板稀释涂布培养,再根据形态学特征进行区分,然后对所分离的菌株进行核糖体DNA 扩增片段限制性内切酶分析(ARDRA),证明冬季刺参肠道中优势菌为厚壁菌门芽孢杆菌纲芽孢杆菌属的巨大芽孢杆菌B.megaterium、苏云金芽孢杆菌B.thuringiensis,以及灿烂弧菌和假单胞菌属的施氏假单胞菌P.stutzeri[58]。应用变性梯度凝胶电泳(PCR-DGGE)技术分析表明,刺参肠道优势菌依次为变形菌门、拟杆菌门和柔膜菌门,其中γ-变形菌纲丰度最高,相对含量在 32.0%~33.6%之间[62]。研究者从刺参肠道内容物中分离到11株细菌,通过16S rDNA V3区基因测序,将其归类为厚壁菌门的梭菌属和变形菌门的假单胞菌属[63]。
总结分析发现,不同研究人员采用不同研究方法得到的结果也不尽相同,这主要是由于纯培养的方法、以16S rDNA为基础的DNA指纹图谱技术和高通量测序技术均会受到培养条件、模板质量、测序效率及生物信息学分析等因素的影响,也会对菌群分析造成限制。但不同的研究结果均证实,变形菌门、厚壁菌门和拟杆菌门在刺参肠道菌群中均具有较高的丰度,为刺参肠道菌群中的优势菌群。
王印庚等[64]研究发现,刺参肠道的位置不同,其肠道菌群的结构和功能也不同。刺参的肠道分为前肠、中肠及后肠,前肠主要负责食物的消化,后肠则发挥对营养物质的吸收和代谢作用[65],肠道功能的差别与肠道内菌群的差异密切相关。应用高通量测序方法分析刺参前中后肠的菌群结构,发现前肠的细菌多样性高于后肠,乳球菌属Lactococcus、芽孢杆菌属为前肠和中肠的共有优势菌属,黄杆菌属Formosa和乳球菌属Lactococcus为后肠的优势菌属,前肠和中肠中的细菌群落多样化可能在食物的消化分解中扮演着重要角色[66]。而采用细菌培养法研究显示,弧菌属和假单胞菌属为刺参肠道的主要优势菌属,后肠的菌群种类呈现出多样化,而前肠菌群种类略显单一[67]。利用PCR-DGGE技术分析发现,池塘养殖刺参前肠、中肠、后肠内容物的细菌群落主要为α-变形菌纲、β-变形菌纲、δ-变形菌纲和拟杆菌纲等的细菌,其优势菌主要为γ-变形菌纲[62]。对刺参中肠和后肠肠道菌群高通量测序发现,中肠和后肠的肠道群落主要以变形菌门、厚壁菌门、拟杆菌门、疣微菌门和放线菌门为主,但是群落组成也根据肠道位置的不同而有所差异,在患腐皮综合征的刺参样品中,中肠和后肠的放线菌门丰度均显著降低,后肠中与弧菌相关的菌群相互作用更紧密,其肠道菌群助力弧菌对宿主的侵染,因此,后肠的肠道菌群能更及时准确地反映刺参病害的发生[68]。
3.3.2 刺参肠道微生物菌群的变化与疾病的发生 肠道微生物从功能上可以分为共生、益生和病原微生物三大类,其中主要是细菌,也包括真菌、病毒和噬菌体,它们在宿主肠道中保持着一种动态平衡。庞大的肠道微生物群体通过与宿主的长期协同进化,已成为一个与宿主密不可分的后天获得的重要“器官”。肠道微生物这一“器官”发挥的功能多种多样,包括物质代谢、生物屏障、免疫调控及宿主防御等,肠道微生物不仅帮助宿主从食物中吸收营养,还能够合成氨基酸、维生素、抗生素等供宿主利用,并可将产生的毒素加以代谢,减少对宿主的毒害。因此,肠道微生物和宿主存在着互利共生的关系,对于维持宿主的健康发挥着重要作用。
Zhang等[60]对健康刺参和腐皮综合征自然发病刺参肠道菌群的组成和特性分析发现,健康刺参和患腐皮综合征刺参的肠道菌群结构有显著差异,健康刺参的优势菌种类为变形菌门细菌(平均相对丰度为96.37%),其次为疣微菌门(2.47%)和厚壁菌门(0.96%),而腐皮综合征患病组的优势菌种类为变形菌门细菌(平均相对丰度为96.01%),其次是厚壁菌门(2.02%)和疣微菌门(1.22%),相比之下,疣微菌门和厚壁菌门的相对丰度在两组中表现出相反的趋势,这表明细菌结构的变化可能与疾病的发生相关。同时,两种不同状态下刺参肠道菌群优势菌目的相对丰度也存在显著差异,患病组的乳酸杆菌目相对丰度明显高于对照组,而疣微菌目相对丰度显著低于对照组。与健康刺参相比,患病个体的肠道菌群的多样性显著降低;通过对腐皮综合征刺参肠道菌群指示菌的筛选,发现红细菌科的细菌和格氏乳球菌等与该病的发生密切相关;对肠道菌群所介导的功能进行预测,发现在腐皮综合征患病组中肠道菌群介导的与疾病感染和信号转导相关的信号通路增强,如免疫系统中的NOD样信号转导通路等(图7)。

图7 健康和患病状态下刺参肠道菌群所介导的功能差异[60]
3.3.3 环境因子协助肠道微生物促进疾病发生 随着季节的变化,环境理化因子也不断发生变化,导致环境中微生物的组成和数目发生动态变化,最终影响刺参肠道中微生物的组成和数量,进而影响疾病发生。有研究发现,低温对刺参肠道菌群结构有显著影响,在一定范围内,随着温度的降低,对肠道菌群的影响作用越强[69]。对高温(20 ℃)下刺参肠道菌群结构分析发现,高温可以通过改变肠道菌群的结构,协助灿烂弧菌对刺参的侵染[70]。对不同水温下刺参发病率分析表明,在14~26 ℃,随着水温的升高,刺参发病率明显增加,病情严重程度也显著上升,然而,水温高于20 ℃时,病菌感染3 d处理组的刺参发病率和发病程度与对照6 d处理组无显著性差异,甚至呈下降趋势[71]。这表明,温度较低时,病菌是刺参发病的主导因素,温度升高加剧了病菌的感染性。
Zhang等[70]对病原菌灿烂弧菌感染、机械损伤、高温、亚硝酸盐和氨氮处理后的刺参肠道菌群结构分析发现,环境因子胁迫下肠道菌群的数量下降,不同的环境胁迫条件导致α-变形菌纲相对丰富(30.31%~70.74%)较正常组(24.69%)显著增加,而厚壁菌门、放线菌门和δ-变形菌纲相对丰度减少;在细菌科分类水平上,除机械损伤组外,不同环境胁迫条件下,红杆菌科相对丰度(47.81%~68.57%)较正常组(19.76%)显著增加,弧菌科相对丰度(3.90%~11.11%)较正常组(0.71%)显著增加,冷杆菌科Psychromonadaceae相对丰度(2.19%)较正常组(1.30%~7.28%)也显著增加,相反,OM60和瘤胃球菌科相对丰度较正常组下降;利用基于16S的细菌群落功能预测工具(PICRUSt)进一步分析显示,环境因子胁迫下刺参肠道菌群介导的功能与正常组相比,不同的环境因子胁迫组肠道菌群介导的细胞通讯途径(局灶性黏附、紧密连接和黏附连接)、免疫系统(NOD样受体信号途径)、脂质代谢、聚糖生物合成和代谢等通路显著降低(图8)。
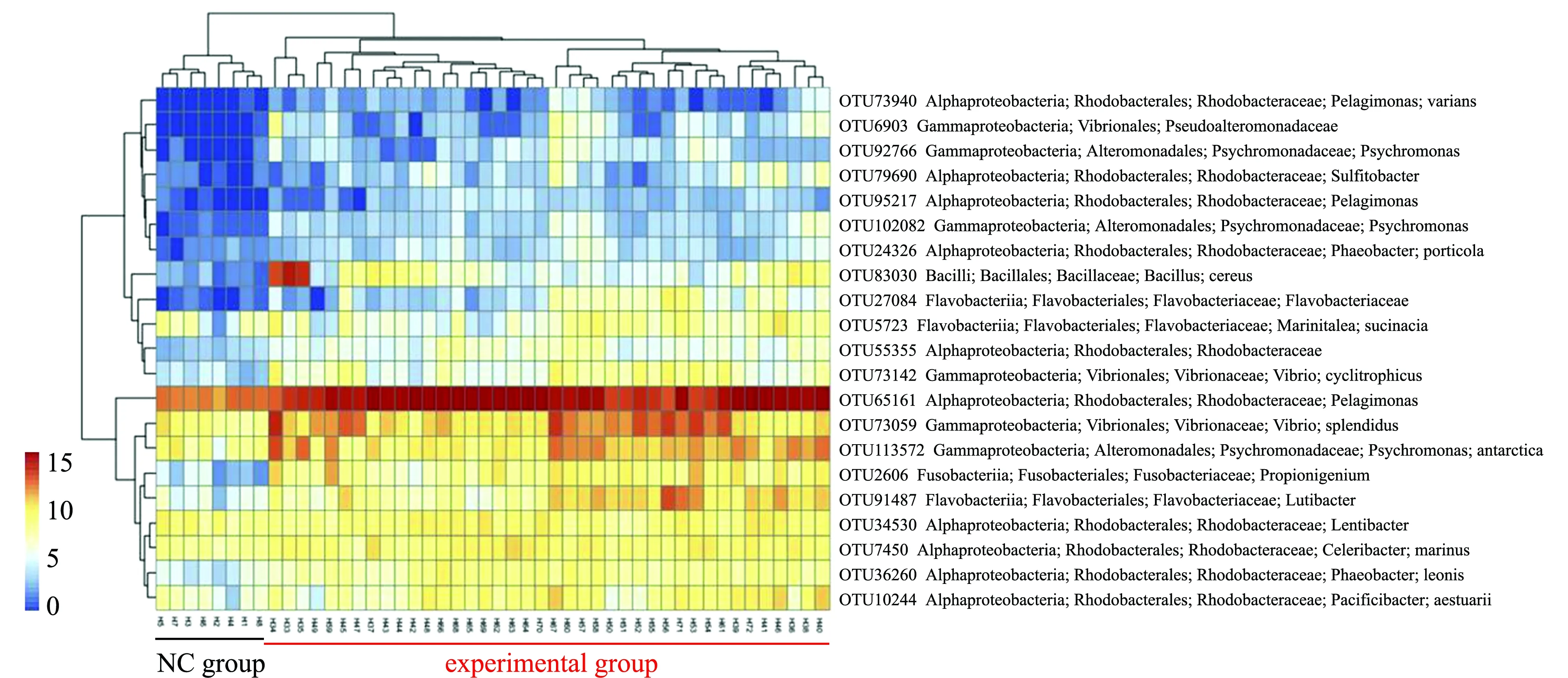
图8 不同环境胁迫条件下刺参28个细菌指标的相对丰度变化[70]
基于IndVal大于0.8,对不同环境因子胁迫下导致刺参腐皮综合征的后肠肠道指示菌进行筛选,获得了21个具有显著指示病害发生的OTU,主要隶属于红细菌科[70]。有研究报道红细菌科细菌是导致水生动物疾病的病因之一,如蟹的乳状病[72]。同时,在各个环境胁迫下,刺参肠道中蜡样芽孢杆菌Bacilluscereus、黄杆菌科、灿烂弧菌和嗜环弧菌Vibriocyclitrophicus丰度均增加;进一步利用RT-qPCR定量各环境因子胁迫组中灿烂弧菌的数量,发现其均高于单独的灿烂弧菌处理组,而机械损伤组灿烂弧菌丰度最高[70]。这说明环境因子在不同程度上协助了灿烂弧菌在刺参肠道中的定植。
4 刺参病害生态防治
4.1 刺参抗病育种
随着刺参产业规模的不断拓展,种质退化、生长缓慢、养殖周期长、抵御环境变化能力差和病害频发,以及商品参品质下降等一系列制约或潜在制约产业发展的问题也日益凸显。为有效解决产业面临的上述问题,辽宁、山东沿海各养殖单位纷纷培育出刺参苗种。目前,已培育的刺参新品种有山东省海洋生物研究院与好当家集团有限公司联合培育的“鲁海1号”、中科院海洋所与山东烟台东方海洋通过产学研结合选育的“东科1号”、山东省海洋资源与环境研究院与多家单位联合培育的刺参新品种“崆峒岛1号”、大连海洋大学培育的“水院1号”、山东安源水产公司培育的“安源1号”和中国水产科学研究院黄海水产研究所与青岛瑞滋集团有限公司合作培育的“参优1号”等。 其中,“参优1号”是中国首个具有抗逆性状的刺参新品种,于2017年通过国家水产原种与良种委员会评审(品种登记号为GS-01-016-2017[73])。历经12年培育的新品种,具有抗病力强、生长速度快和耐高温等优点,成功选育填补了中国抗逆刺参良种培育的空白。刺参“参优1号”抗灿烂弧菌能力强,在6月龄时灿烂弧菌侵染后成活率提高11.68%,可显著提高刺参抗化皮病的能力;生长速度快,池塘养殖收获时其平均体质量提高38.75%,可显著提高刺参的产量和经济效益;成活率高,池塘养殖收获时成活率提高23%以上。
4.2 免疫增强剂
免疫增强剂是指单独或同时与抗原使用均能增强机体免疫应答的物质, 其通过提高养殖动物的非特异性免疫力以增强机体对病原的抵抗力。免疫增强剂的主要作用方式包括提高养殖动物的非特异性免疫、提高养殖动物的特定生长率和抑制病原菌[74]。免疫增强剂对宿主免疫力提升和病原抵抗力的作用主要通过细胞免疫和体液免疫方式来实现。由于免疫增强剂具有比抗生素更安全、比疫苗作用范围广等优点,越来越受到重视,特别是对于提高以非特异性免疫为主的无脊椎动物(对虾、贝类及海参)的免疫力尤为重要。目前,水产养殖中常用的免疫增强剂主要有中草药、多糖类物质、益生菌、动植物与细菌提取物及化学合成物质类等物质。
4.2.1 中草药制剂 中草药对于增强刺参机体免疫力、预防及治疗病原微生物引起的刺参病害,以及促进刺参生长等具有良好效果,同时又因其无抗药性、无残留、无副作用、不引发药源性疾病及抑菌杀菌效果明显而备受人们关注。2009年,Wang等[75]首次研究了黄芪的根及其多糖对刺参非特异性免疫应答的增强作用,使用3%的黄芪细粉(CP)、超细粉(SP)和0.3%的黄芪多糖(APS)喂养刺参60 d后,属于体液反应的溶菌酶活性、活性氧ROS的水平均明显增加;用黄芪超细粉和黄芪多糖喂养60 d后,分别采用灿烂弧菌进行刺参攻毒试验,刺参累计出现症状的比率从对照组的66.67%下降到16.67%和33.34%,而用黄芪细粉喂养60 d后的刺参累计出现症状的比率与对照组相同。之后闫茂仓等[76]对22 味中草药进行了灿烂弧菌体外抑制试验,筛选到黄连、黄芩、连翘和黄柏对灿烂弧菌具有较好的抑菌和杀菌效果。贾晨晨等[77]研究了菊芋全粉对平均体质量为4.7 g稚参的免疫增强作用,结果显示,在灿烂弧菌感染下,15 g/kg菊芋全粉可显著增强刺参免疫因子活力。
中草药虽然可提高刺参的免疫力,然而其在刺参养殖中的使用需谨慎。姚刚等[78]进行了五倍子、乌梅、石榴皮、黄芩和甘草5种中草药的单剂和复方药剂对稚参的急性毒性试验,结果表明,中草药的浓度和配伍是中草药使用过程中最为重要的两个参数,高浓度的中草药对稚参表现出了一定的急性毒性。因此,中草药的使用必须在安全浓度范围内,方可用作刺参细菌性疾病的防治药物。
4.2.2 免疫多糖类 大量的药理和临床研究结果表明,从生物体中提取的天然多糖,具有免疫增强作用强、安全性高和应用广谱等优点,是理想的免疫增强剂。目前水产养殖中使用的多糖类免疫增强剂通常有微生物多糖、甲壳素、壳聚糖和植物多糖等。已用于刺参免疫增强剂的多糖有β-葡聚糖、甘露寡糖、壳聚糖和海藻多糖等(表1),其中β-葡聚糖在刺参免疫增强中的作用得以广泛研究。Zhao等[79]研究表明,饲料中添加一定浓度的β-葡聚糖,对刺参生长、免疫应答和抗灿烂弧菌感染等活性有显著提高,投喂含浓度为1.25 g/kg β-葡聚糖饲料的刺参比投喂含2.50 g/kg β-葡聚糖饲料的刺参生长速度更快,体腔细胞的呼吸爆发活性更高,且死亡率更低,表明β-葡聚糖能够显著降低刺参在灿烂弧菌感染下的死亡率,且具有明显的浓度依赖性。Wei等[80]等报道了来源于浒苔的多糖PEP2具有提高刺参体腔细胞的吞噬能力(PC)、呼吸爆发活性,以及体腔液中酸性磷酸酶(ACP)、过氧化氢酶(CAT)和超氧化物歧化酶(SOD)活性的能力,并显著降低灿烂弧菌感染的刺参累计死亡率。

表1 应用于刺参免疫增强的多糖类免疫增强剂
4.2.3 免疫增强类菌剂 已有研究表明,健康动物体内或养殖环境中具有产酶能力的微生物是益生菌的有效来源。目前已获得了多株潜在的益生菌,并测定了益生菌对刺参生长、消化、免疫及抗病力的影响。杨志平等[84]从健康刺参(体质量为10~30 g)肠道中分离出50株细菌,并对该菌株产淀粉酶、蛋白酶、脂肪酶和纤维素酶等的能力进行了测定,结果获得了3株产酶能力高且不产生溶血的菌株BC26、BC228和BC232,通过注射模式和浸泡感染模式进行了这3株菌的安全性测定,结果表明无论何种感染方式,注射或浸泡感染使用浓度为107CFU/mL 的菌悬液,或投喂含109CFU/g的菌体作为饲料添加剂,都不会引起刺参发病,初步表明这3株菌具有可在刺参养殖中应用的潜力。Yan等[85]筛选到了另一株芽孢杆菌BacillusbaekryungensisYD13,使用浓度为104、106、 108CFU/g的YD13添加到基础饲料中喂养刺参,104、106CFU/g浓度的B.baekryungensisYD13表现出了较强的免疫激活作用,喂养60 d后,刺参体腔液LSZ、ACP及血清ALP、SOD、CAT酶活性均有不同程度的升高,喂养结束后,采用灿烂弧菌进行感染试验,结果表明,104、106、 108CFU/g浓度的B.baekryungensisYD13可使刺参的死亡率下降至15%、5%和30%,均显著低于对照组的50%。
酵母菌也是刺参养殖中常用的免疫增强剂。Liu等[86]筛选获得了对刺参具有免疫激活效果的酵母Metschnikowiasp.C14并投喂刺参,即使在其较低的水平,即104CFU/g时也获得了100%的相对保护率。Ma等[87]分析了仙人掌有孢汉逊酵母菌HanseniasporaopuntiaeC21对刺参的保护效应,H.opuntiaeC21可显著提高刺参体腔细胞的吞噬能力、体腔液的LZM、T-NOS、SOD、AKP和ACP活性,以及刺参体腔细胞裂解上清液中LSZ、T-NOS、AKP和ACP的活性,采用含有H.opuntiaeC21的饲料喂养刺参45 d后,使用灿烂弧菌进行感染试验,结果表明,H.opuntiaeC21可以对灿烂弧菌感染的刺参产生100%的保护率,刺参的死亡率由对照组的50%降低至H.opuntiaeC21饲养组的0%。Wang等[88]使用3种不同浓度(105、106、107CFU/g)的海洋红酵母RhodotorulabenthicaD30添加到饲料中并投喂刺参,刺参体腔液中的淀粉酶、纤维素酶和褐藻酶活性均有所提高,且随着D30浓度的增加,消化酶的活性亦有所增加,喂养后的刺参使用灿烂弧菌感染,结果表明,用浓度为105、106CFU/g的R.benthicaD30喂养后的刺参死亡率均为10%,用浓度为107CFU/g的R.benthicaD30喂养后的刺参死亡率为0%,而喂养基础饲料的对照组刺参死亡率为54.45%。
综合芽孢杆菌和酵母菌作为刺参免疫增强剂的研究,不难发现,在使用相同量的菌剂条件下,酵母菌明显具有更强的免疫增强作用,且菌剂对刺参的免疫刺激作用具有菌株特异性和剂量依赖性。这一特点在Yang等[89]的研究中表现得更为突出。Yang等同时研究了B.cereusG19、B.cereusBC-01和ParacoccusmarcusiiDB11 菌株对刺参的免疫增强作用,采用含有B.cereusG19的饲料喂养刺参后,其体腔细胞的吞噬能力、呼吸爆发活性和AKP酶活性增强,采用含有B.cereusBC-01的饲料喂养刺参后,其体腔细胞的呼吸爆发活性和AKP酶活性增强,采用含有P.marcusiiDB11的饲料喂养刺参后,其体腔细胞的PC、AKP和SOD酶活性增强。因此,可以得出结论,补充B.cereusG19和BC-01的饲料可以显著改善刺参的生长性能和体腔细胞的免疫反应,而补充P.marcusiiDB11的饲料可以积极改善刺参的生长性能和体腔细胞的免疫反应。
此外,不同的细菌复合物也被用于刺参免疫增强作用的研究。使用枯草芽孢杆菌B.subtilisYB-1和B.cereusYB-2的益生菌混合物[90],以及使用乳酸菌、鞘氨单胞菌和醋酸杆菌的组合[91],均获得了比使用单菌更好的免疫刺激效果。使用枯草芽孢杆菌BacillussubtilisT13喂养刺参30 d后,B.subtilisT13对刺参的特定生长率有显著影响,饲料中加入109CFU/g的B.subtilisT13可显著提高刺参体腔细胞的PC、呼吸爆发活性及T-NOS、TCC、SOD酶活性,使用含有109CFU/g的B.subtilisT13的饲料喂养30 d,在灿烂弧菌感染后,刺参的累计死亡率为20.0%,显著低于对照组刺参的56.2%,而使用105、107CFU/g的T13饲养后,刺参的累计死亡率均为50.0%[92]。使用蜡样芽孢杆菌B.cereusEN25喂养刺参30 d,B.cereusEN25对刺参的特定生长率及TCC、ACP活性均无显著性影响,但可显著提高刺参的PC、呼吸爆发活性和T-NOS活性,饲料中添加浓度为107CFU/g的EN25的刺参在灿烂弧菌感染后死亡率为33.3%,远低于对照组刺参的64.2%[93]。
4.3 拮抗菌剂
灿烂弧菌是刺参养殖的主要病原菌,目前已经报道的拮抗菌剂的筛选多是针对灿烂弧菌展开的(表2)。从来自大连三山岛的健康刺参体表及肠道中分离得到37株细菌,采用十字交叉划线法和纸片法筛选出对灿烂弧菌具有拮抗作用的菌株28株,其中7株菌均是通过竞争营养物质与生存空间或者产生有抑菌活性的胞外产物两种方式协同抑菌获得的,其中假单胞菌Pseudoalteromonassp.CG-6-1主要分泌蛋白类等极性的生物大分子,其胞外产物经过80%的硫酸铵沉淀时获得的沉淀物抑菌活性最高,产生的抑菌圈直径可达到25 mm[94]。使用同样的筛选策略,对刺参体表的细菌进行了分离和拮抗活性的筛选,获得一株枯草芽孢杆菌B.subtilisCG,并通过正丁醇提取、硅胶G薄层层析、官能团显色反应和红外光谱分析,最终推断主要的拮抗物质为氨基糖苷类化合物[95]。从红岛刺参养殖区泥样中分离出27株可培养细菌,其中包含一株灿烂弧菌LJ08,采用科赫法则回归感染表明,LJ08对幼参具有致病性,泥样中分离获得的可培养细菌经安全性、抑菌活性和毒性检测,筛选出对灿烂弧菌LJ08有明显拮抗作用的柠檬黄假交替单胞菌PseudoalteromonascitreaLJ03、海水芽孢杆菌BacillusaquimarisLJ04和白色食琼脂菌AgarivoransalbusLJ06[96]。

表2 拮抗菌的筛选及拮抗特性
以刺参腐皮综合征主要致病菌假交替单胞菌P.nigrifaciens、灿烂弧菌V.splendidus、副溶血弧菌V.parahaemolyticus和溶藻弧菌V.alginolyticus为指示菌,对山东东营的刺参养殖池塘水样和底泥样品中的可培养细菌进行了拮抗菌的分离鉴定,获得了拮抗菌贝莱斯芽孢杆菌BacillusvelezensisDY-6,该菌对受试致病菌均有较好的抑制作用;进一步对该菌的安全性进行了评估,使用高浓度的B.velezensisDY-6菌体浸泡刺参,在试验期间浸浴组和投喂组刺参状态良好,无发病和死亡现象[97]。这表明B.velezensisDY-6具有良好的抑菌能力,对刺参养殖过程中的细菌病害具有较好的防治潜力。
从健康的虎斑乌贼体内筛选到一株可以拮抗刺参病原灿烂弧菌V.splendidusAJ01的弧菌Vibriosp.V33,将Vibriosp.V33的上清液加入灿烂弧菌V.splendidusAJ01培养液中,可抑制38%的灿烂弧菌生长;将Vibriosp.V33的无菌上清液于100 ℃加热5 min,其拮抗活性未发生显著变化,这提示弧菌Vibriosp.V33产生的拮抗为非蛋白类物质;为了量化Vibriosp.V33对灿烂弧菌的拮抗作用,在自然海水中,将Vibriosp.V33与V.splendidusAJ01共孵育24 h后,与对照组相比,V.splendidusAJ01的数量大大减少,数量约降低89%;进一步对Vibriosp.V33上清液中所含的拮抗物质活性从分子量、热稳定性、极性等角度进行了表征,结果表明,Vibriosp.V33分泌的拮抗物质具有热稳定、水溶性及分子量小等特点[98]。
另外,对Vibriosp.V33和V.splendidusAJ01分别进行铁吸收的测定,发现Vibriosp.V33具有更高的铁吸收效率,因此,推测Vibriosp.V33主要是通过拮抗灿烂弧菌的铁吸收过程实现对灿烂弧菌V.splendidusAJ01生物量的抑制[102]。分别使用Vibriosp.V33、V.splendidusAJ01及二者的混合菌进行刺参感染试验,结果表明,Vibriosp.V33浸泡感染的刺参未出现死亡和病症,而V.splendidusAJ01浸泡感染的刺参累计死亡率为70%,采用二者混合菌共浸泡感染的刺参累计死亡率为40%[98]。这说明,Vibriosp.V33是一株安全性较高的菌株,且其对灿烂弧菌感染的刺参具有43%的相对保护率。
从健康刺参肠道微生物中筛选出对V.splendidusAJ01具有拮抗活性的蜡状芽孢杆菌BacilluscereusLS2,B.cereusLS2的上清液对V.splendidusAJ01的生长具有较强的抑制作用,抑制率可达到86.83%;采用活性追踪试验进一步表征了该拮抗物质的性质,B.cereusLS2分泌的拮抗物质为相对分子质量小于3 000的小分子物质,且对胰蛋白酶敏感,对温度具有较高的稳定性;采用6 mol/L的盐酸沉淀B.cereusLS2的无菌上清液,得到的沉淀物仅溶于甲醇中,表明该物质具有脂溶性;原位酸水解和茚三酮显色试验显示,未经水解的沉淀物呈现紫色,而水解后的沉淀物呈现橙黄色,因此,该拮抗物质具有多肽的性质[99]。结合所测定的性质推断,蜡状芽孢杆菌LS2产生的拮抗灿烂弧菌物质为小分子的脂肽。
4.4 噬菌体在控制刺参病害微生物中的应用
噬菌体是一类能够感染细菌、真菌、螺旋体或放线菌等微生物的病毒总称。噬菌体本身不能繁殖,必须在活菌内寄生。噬菌体是自然界中存在的最丰富的物种之一,据估计其数量高达1031数量级[103]。相较于普通真核细胞病毒,噬菌体的培养更加简便、经济、快捷且其对化学和物理因素较稳定,易于获得纯培养。噬菌体具有极高的宿主特异性,对人和动物没有感染性,只寄生于其易感染的宿主细胞内,因此,可作为一种理想的抗菌生物制剂替代抗生素[104-105]。
目前,关于噬菌体在抑制病原菌感染刺参过程中的应用已有较多的报道。Zhang等[106]利用溶藻弧菌噬菌体研究其对刺参的保护效果发现,噬菌体感染复数为10、1和0.1的刺参(10 g±2 g)存活率分别为73%、50%和47%,这一结果与没有噬菌体的对照组(3%的存活率)具有显著差异,而用两种抗生素治疗(5 mg/L多西环素和10 mg/L卡那霉素)后的存活率分别为80%和47%,显示抗生素和噬菌体具有类似的治疗效果。Li等[107]用3种噬菌体单独或以1∶1∶1的比例分别对灿烂弧菌进行体外抑制试验,与未处理组相比,对所有被噬菌体处理的灿烂弧菌生长有显著的抑制作用,且混合噬菌体(噬菌体鸡尾酒)比3种噬菌体单独使用具有更明显的抑制效果;进一步进行噬菌体鸡尾酒与抗生素及单一噬菌体对刺参的保护效果研究中发现,未添加抗生素的刺参存活率为18%,抗生素处理的为82%,混合噬菌体处理的为82%,噬菌体pps-1、pps-2和pps-3处理的分别为65%、58%和50%。这表明,利用噬菌体,特别是用不同噬菌体的组合来控制刺参中的弧菌感染,可能是一种可行的方法。
5 存在问题及展望
疾病的发生是宿主免疫力和病原致病力斗争的结果和呈现形式,在这之中环境因子发挥着重要的调节作用。因此,宿主、病原体和环境是决定疾病发生的关键三要素,也是建立刺参疾病绿色防治的理想靶点。目前,在刺参免疫防御机制方面取得了重要进展,鉴定了机体免疫调控的多种信号通路及其调控途径,获得了具有潜在应用价值的效应分子,选育了抗病力强的“参优1号”刺参新品种,但在病原致病机制研究方面进展相对迟缓,主要聚焦在灿烂弧菌溶血素、铁吸收等毒力因子发掘和功能方面,而涉及环境因子的研究更多地集中在某一特定环境因子的变化对宿主免疫指标的影响,对于环境因子如何调控疾病发生的研究几乎是空白。因此,今后应在以下几方面开展深入研究。
1)为实现刺参产业健康可持续发展,亟须强化宿主免疫力、病原致病力和环境因子三者研究的深度和交叉融合。在刺参免疫防御研究方面,应拓宽细胞免疫研究的深度和广度,强化体腔细胞分型和功能研究,建立稳定传代的刺参特定体腔细胞系。基于病原-宿主互作理论,阐明病原微生物激活免疫信号通路,实现免疫逃逸的分子机制。
2)在病原致病机制方面,加强灿烂弧菌以外的其他病原致病因子的发掘和功能研究,突出不同病原感知宿主免疫细胞/效应分子的分子和细胞学机制,强化宿主对病原毒力因子表达的调控作用研究。
3)在环境调控疾病发生研究方面,阐明关键环境因子对宿主免疫防御能力和病原毒力因子表达的调控关系,明确环境污染物介导刺参疾病发生的分子机制,阐释环境微生物(包含病原微生物)在宿主体内的迁移、定植和功能。
致谢:郭明老师和段雪梅老师参与了本文的资料搜集与写作工作。
