缺铁胁迫下垂丝海棠幼苗响应外源糖处理的生理特性
2021-06-20石福辉缐旭林王秀耿喜红唐雯王延秀
石福辉 缐旭林 王秀 耿喜红 唐雯 王延秀
摘要:以8葉龄垂丝海棠实生苗为试验材料,通过喷施不同浓度的葡萄糖与蔗糖,研究外源糖对其缺铁胁迫下生理的影响。结果表明,垂丝海棠叶片的相对叶绿素(SPAD)含量、相对含水量、超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、亚铁离子浓度(Fe2+)在 -Fe处理下相比于对照显著下降,在外施葡萄糖和蔗糖后,其含量相比于-Fe处理显著上升,且具有浓度效应,随处理浓度的增加呈先升后降趋势;而脯氨酸(Pro)含量随着处理浓度的增加呈上升趋势,各处理的Pro含量均显著高于对照。此外,缺铁处理下丙二醛(MDA)相比于对照显著上升,在外施葡萄糖和蔗糖后其升幅显著下降,且存在浓度效应,随处理浓度的增加呈先升后降趋势。依据主成分得分排序,外源葡萄糖和蔗糖对垂丝海棠缺铁胁迫缓解能力由高到低均为:CK > -Fe+2 > -Fe+4 > -Fe+1 > -Fe+8 > -Fe。研究发现,2 mM的外源葡萄糖和蔗糖均能通过调节渗透物质、增强抗氧化酶活性、增加生物膜的稳定性、提高亚铁离子含量以达到缓解缺铁胁迫的效应,且蔗糖处理的缓解效应更好。
关键词:垂丝海棠;缺铁胁迫;葡萄糖处理;蔗糖处理;主成分分析
中图分类号:S661.4 文献标志码:A 文章编号:1001-1463(2021)05-0006-09
doi:10.3969/j.issn.1001-1463.2021.05.003
Physiological Characteristics of Malusa halliana Seedlings under Iron Deficiency Stress in Response to Exogenous Sugar Treatment
SHI Fuhui, XIAN Xulin, WANG Xiu, GENG Xihong, TANG Wen, WANG Yanxiu
(College of Horticulture, Gansu Agricultural University, Lanzhou Gansu 730070, China)
Abstract:Malusa halliana seedlings of 8 leaf age were used as experimental materials to study the effects of exogenous sugar on physiological characteristics of apple rootstock under iron deficiency stress by sprayed with different concentrations of glucose and sucrose. The results showed that the vertical wire relative haitang leaf chlorophyll content(SPAD), relative water content, superoxide dismutase(SOD), peroxidase(POD), catalase(CAT), concentration of ferrous ions(Fe2+) under -Fe treatment compared with the control(CK) decreased significantly, after the glucose and sucrose, compared with its content -Fe a marked increase in the processing, and has the concentration effect, with the increase of the concentration showed a trend of rose first and then fell. Proline content(Pro) showed an increasing trend with the increase of treatment concentration, and the Pro content of each treatment was significantly higher than CK. In addition, malondialdehyde(MDA) significantly increased when compared with CK under iron deficiency treatment, and its increase significantly decreased after the application of glucose and sucrose, and there was a concentration effect, which a trend of rose first and then fell with the increase of the treatment concentration. According to the order of principal component score, the ability of exogenous glucose and sucrose to alleviate the iron deficiency stress of malus tuber from high to low was: CK > -Fe+2 > -Fe+4 > -Fe+1 > -Fe+8 > -Fe. It was found that both 2 mM exogenous glucose and sucrose could alleviate the effect of iron deficiency stress by regulating osmotic substances, enhancing the activity of antioxidant enzymes, increasing the stability of biofilm, and improving the content of ferrous ions, and sucrose treatment had better mitigative effect.
Key words:Malusa halliana;Iron deficiency stress;Glucose treatment;Sugar treatment;Principal component analysis
铁作为植物生长发育过程中的一种必需微量元素,在光合作用、呼吸作用、氮的固定等生理代谢过程中发挥着极为重要的作用[1 ]。缺铁会对植物产生不同程度的伤害,轻则叶绿素合成受阻,光合速率下降,严重时甚至会使整个植株衰亡[2 ]。苹果是我国果产区的支柱性产业[3 ],西北黄土高原是我国苹果的优势产区,但该区域70%以上的土壤为含NaCl与NaHCO3的高pH石灰质土壤,铁易形成难溶性化合物,致使苹果易发生缺铁黄化症状[4 - 5 ],严重制约着苹果产业的发展。
葡萄糖和蔗糖不仅在植物生长发育中起着重要作用,还作为信号物质参与调控植物对生物和非生物胁迫的响应[6 - 7 ]。胡梦芸 等[8 ]在小麦的研究中发现,低浓度的外源葡萄糖可以诱导小麦叶片气孔关闭,降低蒸腾速率,使其适应干旱的能力增强。Siringam等[9 ]在水稻上的研究表明,外源蔗糖能够提高感盐水稻中果糖和葡萄糖的含量,增加高感盐水稻的耐盐性。外源葡萄糖处理还能有效提高青花菜中SOD、POD及CAT的活性,维持离体青花菜较高水平的总糖含量,提高自由基清除率,提升青花菜品质[10 ]。Boriboonkaset等[11 ]研究发现,经蔗糖处理过的水稻种子在高盐胁迫(342 mmol/L NaCl)下,表现出较强的抗性。目前,关于外源施糖处理的研究多集中在干旱及盐碱等非生物胁迫方面,而对缺铁胁迫下外源糖处理对木本植物尤其苹果砧木的研究则鲜见。
垂丝海棠(Malus halliana Koehen)原产于甘肃河西走廊,生长于干旱、盐碱生境,是一种抗寒、耐旱、耐盐碱的苹果砧木资源[12 - 13 ],又名倒挂珍珠,其在西北黄土高原地区,长势良好 ,具有较强抗性[14 - 15 ]。本试验以8叶龄垂丝海棠为试材,旨在研究外源葡萄糖和蔗糖在不同浓度下对缺铁胁迫是否具有缓解作用,为西北石灰性土壤苹果栽培提供理论参考。
1 材料与方法
1.1 材料
选取饱满一致的垂丝海棠种子,经2 g/kg高锰酸钾消毒30 min后用流水冲洗12 h,置于4 ℃下沙藏30 d。发芽后,选取发芽一致的种子种于塑料钵(12.0 cm×18.0 cm)中,每个塑料钵含2.5 kg基质(草炭、珍珠岩、蛭石质量比为3∶1∶1),每钵5株,置甘肃农业大学玻璃温室统一管理,定期观察。
1.2 试验方法
采用水培法。设置正常供铁(CK)和缺铁胁迫(-Fe),外加4种浓度(1 mM、2 mM、4 mM、8 mM)的蔗糖(Suc)和葡萄糖(Glc)共10个处理,即CK(+Fe 40 μmol/L)、-Fe(+Fe 4 μmol/L)、-Fe+Glc 1 mM、-Fe+Glc 2 mM、-Fe+Glc 4 mM、-Fe+Glc 8 mM、-Fe+Suc 1 mM、-Fe+Suc 2 mM、-Fe+Suc 4 mM、-Fe+Suc 8 mM,3次重复。测定各处理下相对叶绿素含量(SPAD)、相对含水量、脯氨酸含量(Pro)、丙二醛含量(MDA)及抗氧化酶活性,并通过主成分分析对其进行综合评价。
幼苗长到8片真叶时选取长势一致、无病虫害幼苗转移到1/2 Han's[16 ]营养液中进行15 d的预培养。将预培养后的苗转移到含上述处理的500 mL塑料钵中,用电动氧气泵持续通氧。培养条件为温度26 ℃、光照16 h/黑暗8 h、光照强度为6 000 lx、相对湿度光照时为85%,黑暗时为80%。铁由Fe(III)-EDTA提供,处理7 d后进行各项指标测定。
1.3 测定指标及测定方法
胁迫7 d后,取植株中上部大叶节以上的功能性叶片,洗净并剪掉叶脉磨碎用于相关指标测定。叶绿素含量的测定参照朱祖雷等[17 ]的方法, 采集垂丝海棠叶片,去除叶脉并洗净,称取鲜样品0.2 g于玻璃试管中,加入10 mL 80%的丙酮进行24 h提取,在波长645、652、663 nm下测定吸光度,计算叶绿素a(Chl a)、叶绿素b(Chl b)含量;MDA 含量的测定参照Wang等[18 ]的方法,采用硫代巴比妥酸法測定;Fe2+的测定参考刘伟[19 ]的方法,取处理后的垂丝海棠叶片,用去离子水洗净,擦干表面水分,组织放置于烘箱中烘干,盐酸处理后上清液用原子吸收法测定Fe2+含量;游离脯氨酸含量采用酸性茚三酮法[20 ];叶片含水量的测定参照邬燕等[21 ]的方法,采用烘干法测定,将叶片剪下来,称得鲜重Wr,之后加入蒸馏水使叶片吸水饱和,称取饱和时的重量Wt,再称得烘干后恒重Wd。叶片相对含水量=[(Wr-Wd)/(Wt- Wd)]×100%。超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化酶(CAT)活性的测定均采用试剂盒法(购自苏州科铭)。
1.4 数据处理与分析
用 Excel 2013 及 Origin 8.0 进行数据处理及作图,用 SPSS 22.0 进行方差分析和主成分分析。采用单因素 ANOVA 的 LSD 比较处理间的差异显著性(α=0.05)。
2 结果与分析
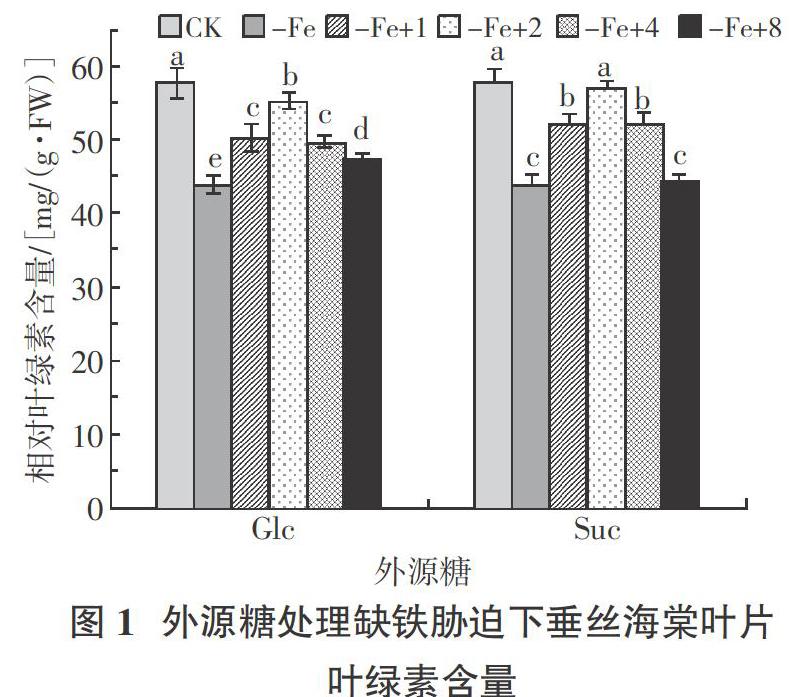
2.1 外源糖处理对缺铁胁迫下垂丝海棠叶绿素含量
如图1所示,缺铁胁迫下,外源施加葡萄糖时,垂丝海棠叶片的叶绿素含量随处理浓度的增加呈先升高后降低的趋势。其中,-Fe+2处理下相对叶绿素含量显著低于 CK,为 55.48 mg/(g·FW),显著高于-Fe处理及其他处理。外源施蔗糖时,垂丝海棠叶片叶绿素含量变化和外施葡萄糖一致,且在-Fe+2处理下叶绿素含量达到峰值,为 57.04 mg/(g·FW),与CK无显著差异,但显著高于-Fe处理及其他处理。
2.2 缺铁胁迫下不同糖类对垂丝海棠Fe2+的影响
由图2可知,在缺铁胁迫下施加外源葡萄糖时,垂丝海棠叶片Fe2+含量随处理浓度的增加呈先升后降的趋势。各处理的Fe2+含量分别为20.78 mg/g(-Fe)、21.69 mg/g(-Fe+1)、24.96 mg/g(-Fe+2)、22.75 mg/g(-Fe+4)、17.96 mg/g(-Fe+8);相比于CK,分别降低了18.7%、15.2%、2.4%、11.06%、29.83%,-Fe+2处理下的降幅为最小,-Fe+8处理下的降幅最大,-Fe处理次之。当施加外源蔗糖时,其变化趋势与蔗糖相一致,呈先升高后下降的趋势。在-Fe+2处理下达到最大值,为25.94 mg/g,与CK无显著差异,显著高于-Fe处理,相比于CK提高了28.93%
2.3 外源糖处理对缺铁胁迫下垂丝海棠脯氨酸含量的影响
如图3所示,当施加外源葡萄糖时,垂丝海棠叶片脯氨酸(Pro)含量均较CK呈升高的趋势。但不同处理浓度下的升幅不同,在-Fe+8处理下达到峰值,为101.40 mg/g,较CK提高了17.3%;在-Fe处理下升幅最小,为88.76 mg/g,比CK提高了5.8%。当施加外源蔗糖时,垂丝海棠叶片的Pro含量也呈现出与施加外源葡萄糖一致的趋势,且在-Fe+8处理下达到峰值,为111.53 mg/g,显著高于其他处理,相比于CK,提升了27.68%。
2.4 外源糖处理对缺铁胁迫下垂丝海棠叶片相对含水量的影响
由图4可得,缺铁胁迫下施加外源葡萄糖时,垂丝海棠叶片含水量在-Fe+4处理下达到最大值,显著低于CK,显著高于-Fe、-Fe+1及-Fe+2处理。施加外源蔗糖时,垂丝海棠叶片含水量在-Fe+4处理下叶绿素含量达到峰值,为 74.82%,与CK(74.8)无显著差异,但显著高于其他处理。
2.5 外源糖处理对缺铁胁迫下垂丝海棠SOD、POD、CAT活性的影响
由图5可以看出,缺铁胁迫后施加外源葡萄糖时,垂丝海棠叶片SOD活性随处理浓度的增加呈先升后降的趋势,在-Fe+2处理下达到最大值,显著高于-Fe处理,相比-Fe升高了16.34%。施加外源蔗糖时,与外施葡萄糖变化趋势一致,SOD活性呈先升后降。在-Fe+2处理下SOD活性最高,为270.67 U/(g·FW),显著低于-Fe处理,相比-Fe升高了15.23%。
施加外源葡萄糖时,垂丝海棠叶片其他处理的POD活性相比于-Fe处理均升高,但不同处理升幅不同。在-Fe+2处理下POD活性最高,为111.69 U/(g·FW);在Fe+8处理下POD活性最低,为94.25 U/(g·FW),显著高于-Fe处理。施加外源蔗糖时,垂丝海棠叶片POD活性与外源施葡萄糖的变化趋势相一致。在-Fe+2处理下POD活性最高,为106.56 U/(g·FW),显著高于-Fe;在-Fe+8处理下POD活性最低,为94.25 U/(g·FW),显著高于-Fe处理。
施加外源葡萄糖时,垂丝海棠叶片其他处理CAT活性相比于-Fe处理均升高,但不同处理的升幅不同。在-Fe+2处理下CAT活性最高,相比CK提升了74.87%;在Fe+1处理下CAT活性较低,相比CK提升了47.81%。施加外源蔗糖时,垂丝海棠叶片其他处理CAT活性相比于-Fe处理均升高,但不同处理的升幅不同。在-Fe+4处理下活性最高,为44.21 nmol/(min·g) FW,显著高于-Fe;在Fe+8处理下活性较低,为37.99 nmol/(min·g) FW,显著高于-Fe。
2.6 外源糖处理对缺铁胁迫下垂丝海棠丙二醛含量的影响
如图6所示,当施加外源葡萄糖时,缺铁胁迫下,垂丝海棠叶片丙二醛含量随处理浓度的增加呈先降后升的趋势。MDA在各处理下均显著高于CK,其中-Fe处理下MDA含量最高,为6.12 μmol/(g·FW);而-Fe+1 处理下则较低,为 4.74 μmol/(g·FW),高于 CK[4.25 μmol/(g·FW)]。当施加外源蔗糖时,垂丝海棠叶片丙二醛含量随处理浓度的增加也呈先降后升的趋势。MDA在各处理下均显著高于CK。与 CK 相比,-Fe(6.13)升幅最高,显著高于其他处理;-Fe+2(4.60)升幅最小,顯著小于其他处理。
2.7 缺铁胁迫下不同浓度外源葡萄糖对垂丝海棠叶片生理效应的综合评价
2.7.1 相关性分析 将测得的处理后各项指标进行相关性分析,得到相关系数矩阵。对处理后垂丝海棠的8个相关指标进行相关性分析,结果见表1。表明垂丝海棠叶片的SPAD与SOD呈极显著正相关(P < 0.01),与POD、Fe2+、CAT呈显著正相关(P < 0.05),与 MDA 呈极显著负相关(P < 0.05)。
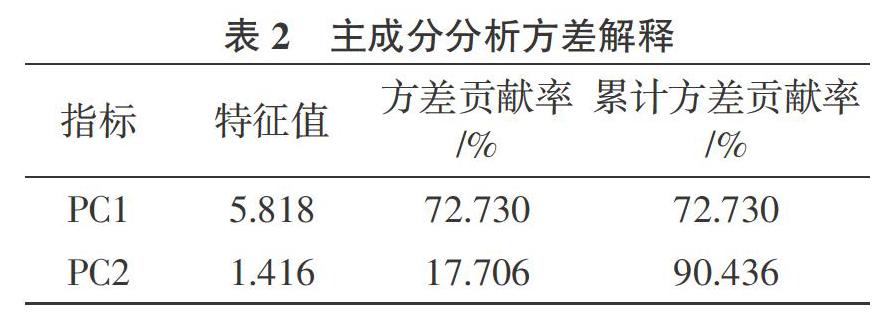
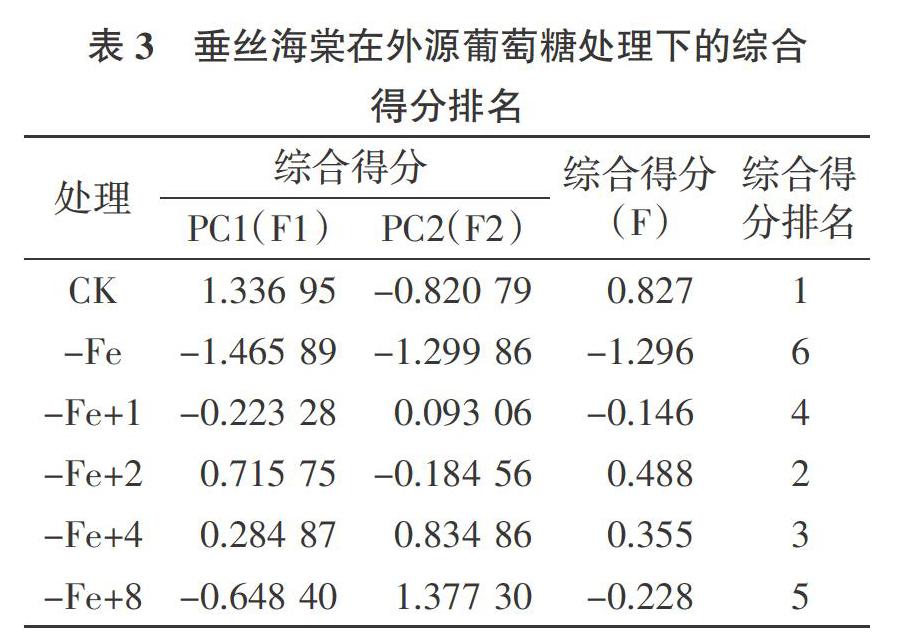
2.7.2 主成分分析 为综合评价缺铁胁迫下不同浓度外源葡萄糖对垂丝海棠的生理响应特性,将胁迫处理后的相关指标进行主成分分析。提取特征值>1 的 2 个主成分,其特征值分别为 5.818和1.416(表 2),方差贡献率分别为72.730%和17.706%,累计方差贡献率达到90.436%,符合分析要求。对上述2个主成分代表的指标进行总得分排名,F1、F2 分别代表第1、第2主成分。综合得分(F)是每个主成分得分与对应贡献率的乘积之和,即:F=F1×72.730%+F2×17.706%。由表 3 可知,垂丝海棠在不同处理下的综合得分分别为0.827(CK)、-1.296(-Fe)、-0.146(-Fe+1)、0.488(-Fe+2)、0.355(-Fe+4)、 -0.228(-Fe+8),即缺铁胁迫下不同浓度外源葡萄糖对垂丝海棠生理特性的缓解效应依次为CK、-Fe+2、-Fe+4、-Fe+1、-Fe+8、-Fe。
2.8 缺铁胁迫下不同浓度外源蔗糖对垂丝海棠叶片生理效应的综合评价
2.8.1 相关性分析 将处理后的各项指标进行相关性分析,得到相关系数矩阵。对处理后垂丝海棠的相关指标进行相关性分析,结果见表4。表明,垂丝海棠叶片SPAD与SOD、POD、Fe2+呈极显著正相关(P < 0.01),与RWC呈显著正相关(P < 0.05),與 MDA 呈显著负相关(P < 0.05)。
2.8.2 主成分分析 将胁迫处理后的8个相关指标进行主成分分析,提取特征值>1 的 2 个主成分,其特征值分别为 6.018和1.289(表2),方差贡献率分别为75.225%和16.116%,累计方差贡献率达到91.341%,符合分析要求。对上述2个主成分代表的指标进行总得分排名,F1、F2 分别代表第一、第二主成分。综合得分(F)是每个主成分得分与对应贡献率的乘积之和,即:F=F1×75.225%+F2×16.116%。由表6可知,垂丝海棠在不同处理下的综合得分分别为0.831(CK)、-1.292(-Fe)、0.077(-Fe+1)、0.433(-Fe+2)、0.426(-Fe+4)、-0.474(-Fe+8),即缺铁胁迫下不同浓度外源蔗糖对垂丝海棠生理特性的缓解效应依次为CK、-Fe+2、-Fe+4、-Fe+1、-Fe+8、-Fe。
3 结论与讨论
铁是植物生长必需的营养元素。植物遭受到缺铁胁迫时生长会受到限制,致使作物减产,品质变劣[22 ]。糖在植物的生命周期中扮演着重要作用,既是能量来源和结构物质,又是一类可响应各种逆境的信号分 子[23 - 25 ]。叶绿素是一种重要的光合色素,对光能的吸收、传递及转化起着重要作用,其含量的高低是衡量植物耐缺铁性的一个重要指标[26 ]。本试验中,在-Fe处理下,叶绿素含量相比于对照显著降低,在施加不同浓度的葡萄糖和蔗糖后,叶绿素含量相比于-Fe处理显著上升。这可能是由于葡萄糖与蔗糖的加入激活了差异表达基因DEG,使得DEG与铁离子结合,作用于光合作用天线蛋白,促进了卟啉和叶绿素代谢引起的,这与Chen等[27 ]研究相一致。表明外施蔗糖与葡萄糖可以缓解轻度缺铁胁迫。此外,植物遭受缺铁胁迫时,常通过多种机制来活化土壤中的难溶性铁,提高铁的有效性,提升植物对铁的吸收利用,从而维持植物体内铁的稳态平衡。Fe2+作为植物吸收利用的有效形式,是研究植物抗缺铁性的关键指标。在本试验中,缺铁处理下Fe2+相比于对照显著降低,在施加不同浓度的葡萄糖和蔗糖后,除处理-Fe+8外,其余处理均显著高于缺铁处理,这表明外施一定浓度的葡萄糖和蔗糖,可促进植物对铁的吸收利用,能显著提升Fe2+的含量。
SOD、CAT和 POD是重要的植物抗氧化酶,具有提高抵抗活性氧的毒害和环境胁迫的能力。本试验中,缺铁处理下,SOD、CAT和 POD活性相比于对照显著降低,这可能是胁迫超出其忍耐范围时,体内活性氧的形成和清除系统之间的平衡被打破,导致酶活性下降[28 ]。在外施不同浓度的葡萄糖和蔗糖后,各处理的SOD、CAT和 POD活性相比于-Fe处理有着显著提高,表明外施蔗糖和葡萄糖处理能够增强 SOD、POD、CAT 活性,从而增强植物抗氧化系统的防御能力,维持膜系统的稳定,进而在一定程度上缓解缺铁胁迫引起的氧化损伤。MDA是膜脂过氧化作用的产物之一,其不断积累会引起酶和细胞膜的破坏,进而造成一系列生理生化代谢紊乱[29 ]。在本试验中,-Fe处理下MDA相比于对照(CK)显著升高,可能此时植株细胞膜已受到伤害,植株体内活性氧产生和清除系统间的平衡遭到破坏,细胞质膜在活性氧的攻击下发生了过氧化,细胞膜的完整性受到损伤,细胞的生理机能降低,植株生长发育水平下降。然而,外施不同浓度的葡萄糖和蔗糖后,丙二醛含量相比于-Fe处理显著降低。表明外源糖处理能有效缓解MDA含量升高,这与王丽燕等[30 ]在番茄上的研究相同,可能是外源糖增加了垂丝海棠植株保护酶活性及生物膜的稳定性,在一定程度上缓解了缺铁胁迫造成的损伤。脯氨酸(Pro)是植物体内重要的一种渗透调节物质,当植物在遭受逆境条件时其体内的Pro含量会大量积累,提高植物的抗逆 性[31 ]。本文研究表明,缺铁胁迫下垂丝海棠Pro含量呈上升趋势,而通过施不同浓度葡萄糖处理、蔗糖处理与-Fe处理相比,升幅显著高于-Fe处理,表明通过外源葡萄糖与蔗糖处理能促进渗透调节物质的积累来抵抗缺铁胁迫。
本试验通过主成分分析得出,缺铁胁迫下不同浓度外源葡萄糖与蔗糖对垂丝海棠生理特性的影响排名均为CK>-Fe+2>-Fe+4>-Fe+1>-Fe+8>-Fe。外源施葡萄糖与蔗糖能够缓解缺铁胁迫,均以2 mM的作用效果最为显著,其效果蔗糖优于葡萄糖。
参考文献:
[1] BRIAT J F,LOBREAUX S. Iron transport and storage in plants[J]. Trends in Plant Science,1997. 2(5):187-193.
[2] FAN W J,WANG H X,WU Y L,YANG N,ZHANG P. H+-pyrophosphatase IbVP 1 promotes efficient iron use in sweet potato [Ipomoea batatas(l.) lam.][J]. Plant Biotechnology Journal,2017. 15 (6):698-712.
[3] 陈学森,韩明玉,苏桂林,等. 当今世界苹果产业发展趋势及我国苹果产业优质高效发展意见[J]. 果树学报,2010,27(4):598-604.
[4] LI Q,YANG A,ZHANG W H. Efficient acquisition of iron confers greater tolerance to saline-alkaline stress in rice(Oryza sativa L.)[J]. Journal of Experimental Botany,2016, 67(22):6431-6444.
[5] BAUER P. Fitting into the harsh reality:Regulation of iron-deficiency responses in dicotyledonous plants[J]. Molecular Plant,2012,
5(1):27-42.
[6] KEUNEN E,PESHEV D,VANGRONSVELD J,et al. Plant sugars are crucial players in the oxidative challenge during abiotic stress:extending the traditional concept[J]. Plant Cell & Environment,2013,36(7):1242-1255.
[7] JACKSON E,DAVID P. Sugars,signalling,and plant development[J]. Journal of Experimental Botany,2012, 63(9):3367-3377.
[8] 胡夢芸,李 辉,张颖君,等. 水分胁迫下葡萄糖对小麦幼苗光合作用和相关生理特性的影响[J]. 作物学报,2009,35(4):724-732.
[9] SIRINGAM K,JUNTAWONG N,ChAUM S,et al. Salt tolerance enhancement in indica rice (Oryza sativa L. Spp. indica)seedlings using exogenous sucrose supplementation[J]. Plant Omics,2012,5(1):52-59.
[10] 董栓泉,许 凤,王春幸,等. 不同糖处理对离体青花菜抗氧化酶以及品质的影响[J]. 现代食品科技,2016,32(5):199-204.
[11] BORIBOONKASET T,BUNYAJINDA V, CHAUM S,et al. Effect of exogenous sugar classes and concentrations on salt-tolerant ability of indica rice(Oryza sativa L.)[J]. Acta Horticulturae,2007,746:155-163.
[12] 刘家尧,赵可夫. 维管植物抗盐生理的某些问题[J]. 曲阜师范大学学报(自然科学版),1991(2):73-80.
[13] 石晓昀. 3个苹果砧木对长期水分胁迫的响应特性[D]. 兰州:甘肃农业大学,2017.
[14] 贾旭梅,朱燕芳,王 海,等. 垂丝海棠应对盐碱复合胁迫的生理响应[J]. 生态学报,2019,39(17):6349-6361.
[15] 郭爱霞. 苹果砧木垂丝海棠响应缺铁胁迫的叶绿素合成关键基因的筛选、克隆及功能鉴定[D]. 兰州:甘肃农业大学,2019.
[16] HAN Z H,WANG Q,SHEN T. Comparison of some physiological and biochemical characteristics between iron efficient and iron efficient species in the genus malus[J]. Journal of Plant Nutrition,1994,17(7):1257-1264.
[17] 朱祖雷,黄华梨,张露荷,等. 不同施钾量对‘骏枣产量,品质及光合特性的影响[J]. 果树学报,2019,36(12):87-97.
[18] WANG S,LIANG D,LI C,et al. Influence of drought stress on the cellular ultrastructure and antioxidant system in leaves of drought-tolerant and drought-sensitive apple rootstocks[J]. Plant Physiology & Biochemistry,2012,51(none):81-89.
[19] 刘 伟. 乙烯响应因子ERF4/ERF72参与苹果砧木缺铁应答的功能研究[D]. 北京:中国农业大学,2017.
[20] REDILLAS M C F R,PARK S H,LEE J W,et al. Accumulation of trehalose increases soluble sugar contents in rice plants con-ferring tolerance to drought and salt stress[J]. Plant Biotechnology Reports,2012,6(1):89-96.
[21] 邬 燕. 模拟干旱胁迫下葡萄的抗旱生理生化机理研究[D]. 呼和浩特:内蒙古农业大学,2019.
[22] KOBAYASHI T,NISHIZAWA N K. Iron uptake,translocation,and regulation in higher plants[J]. Annual Review of Plant Biology,2012,63(1):131-152.
[23] ELS K,DARIN P,JACO VANGRONSVELD. Plant sugars are crucial players in the oxidative challenge during abiotic stress:extending the traditional concept[J]. Plant,Cell & Environment,2013.
[24] BAENA G L,JEN S. Convergent energy and stress signaling[J]. Plant Cell,2008,12(9):1360-1385.
[25] JAKSO,EVELAND D P. Sugars,signalling,and plant development[J]. Journal of Experimental Botany,2012,63(9):3367-3377.
[26] YANG J Y,ZHENG W,TIAN Y,et al. Effects of various mixed salt-alkaline stresses on growth,photosynthesis,and photosynthetic pigment concentrations of Medicago ruthenica seedlings[J]. Photosynthetica,2011,49(2):275-284.
[27] CHEN P F,CHEN L,JIANG Z R,et al. 2018. Sucrose is involved in the regulation of iron deficiency responses in rice(Oryza sativa L.)[J]. Plant Cell Reports,37:1-10
[28] 黄尧瑶,邓明华,彭春秀,等. 百合花瓣抗氧化酶系统对干旱胁迫响应的研究[J]. 园艺学报,2020,47(4):788-796.
[29] 李天来,李益清. 钙对弱光胁迫下番茄叶片保护酶活性及可溶性蛋白含量的影响[J]. 园艺学报,2008(11):1601-1606.
[30] 王丽燕,薄存娇,徐琳娜,等. 外源葡萄糖对番茄盐胁迫耐受性的影响[J]. 北方园艺,2018(7):9-14.
[31] 任文伟,钱 吉,馬 骏,等. 不同地理种群羊草在聚乙二醇胁迫下含水量和游离脯氨酸含量的比较[J]. 生态学报,2000(2):349-352.
(本文责编:陈 珩)
