钙营养胁迫对加工番茄幼苗营养分配及其生理指标的影响
2021-06-17李秀霞努尔麦麦提·艾麦提齐曼·尤努斯艾克拜尔·毛拉陈远良
李秀霞 努尔麦麦提·艾麦提 齐曼·尤努斯 艾克拜尔·毛拉 陈远良
摘 要:以加工番茄87-5的幼苗为试验材料,设置Hoagland营养液钙浓度分别为0mg/L(不加钙)、160mg/L、320mg/L、640mg/L不同的钙浓度梯度胁迫处理,处理21d后测定其生长状况指标和生理指标。结果表明:不同钙浓度处理下,87-5叶片的氮磷钙含量均高于茎和根,320、640mg/L钙浓度处理下镁含量均降低,640mg/L钙浓度处理下根和茎镁浓度与对照差异显著;320mg/L钙浓度显著提高了加工番茄幼苗的根系活力,640mg/L钙浓度时叶绿素含量比对照显著降低。由此可知,钙浓度亏缺或过剩,均会影响加工番茄根系活力和各器官的钙镁元素分配,导致叶绿素合成减少。
关键词:加工番茄;钙营养胁迫;叶绿素含量
中图分类号 Q945文献标识码 A文章编号 1007-7731(2021)10-0013-04
Effects of Calcium Stress on Nutrient Allocation and Physiological Indexes of Processing Tomato Seedlings
LI Xiuxia1 et al.
(1Xinjiang Agricultural Vocational and Technical College, Changji 831100, China)
Abstract: In this study, the seedlings of processing tomato 87-5 (variety) were used as materials, the calcium concentration of Hoagland nutrient solution was 0mg/L (without calcium), 160mg/L, 320mg/L, 640mg/L. different calcium concentration gradient stress treatments were set, and the growth status and physiological indexes were measured after 21 days. The results showed that the contents of N, P, CA in leaves were higher than those in stems and roots under different Ca concentrations, the contents of Mg in roots and stems were lower under 320mg/L and 640mg/L Ca concentrations, and there was significant difference between roots and stems under 640mg/L Ca concentrations and the control; the root vitality of processing tomato seedlings was significantly improved under 320mg/L Ca concentrations, and the chlorophyll content was significantly lower at 640mg/L Ca concentrations than that of the control. It was speculated that both deficiency and excess of calcium concentration affected root activity and distribution of calcium and magnesium in various organs of processing tomato, resulting in the decrease of chlorophyll synthesis.
Key words: Processing tomato; Calcium stress; Chlorophyll content
鈣是植物必需元素之一,是植物组织构造的重要组成成分,对植物的生理发育起着重要作用[1]。钙是细胞膜结构的组分,可稳定细胞膜、细胞壁,保持细胞的完整性,钙将细胞膜表面的磷酸盐、磷酸酯与蛋白质基桥链接起来,提高膜结构的稳定性和疏水性[2]。作为第二信使,钙还能结合在钙调蛋白(CMA)上,对植物体内多种关键酶起活化作用,对细胞代谢有调节作用[3-4]。钙的吸收、转运和储存对植物的生理功能有着重要影响[5],适合的钙浓度才能维持植物正常的生长,钙浓度低或高会均影响植物的生长状况及生理变化。而高浓度的Ca2+是一种细胞毒害剂,若细胞内Ca2+浓度过高,将会与磷酸反应形成沉淀,从而扰乱与磷代谢有关的生理过程或妨碍正常的信号转导,进而影响植物生长[5-6]。
钙元素过剩会抑制植株对镁、铁、锌等元素的吸收,从而引起其他营养元素的缺素现象,如过剩的钙元素抑制植株对铁元素的吸收而出现叶片缺绿[7]。缺钙会降低人参对P的吸收;低浓度的钙促进K吸收,高浓度的钙抑制K的吸收;Mg、Fe及Cu在一定程度与钙存在拮抗作用,高浓度的钙会抑制它们的吸收;高浓度的钙会抑制Zn向人参地上部分运输;钙的多少对人参吸收N、Mn及Al无显著影响[8]。钙营养元素的缺乏会对加工番茄幼苗造成伤害,钙营养胁迫影响加工番茄幼苗光合色素及光合特性[9]。适量的钙能促进植物的光合作用,进而促进植物体内糖分的合成,而缺钙会明显减少植物各器官可溶性糖的含量[10-11]。钙胁迫能够破坏茶树的光合膜结构,植物体中生产活性氧的主要部位是线粒体和叶绿体[12]。
本试验以番茄幼苗为试验材料,研究番茄对不同钙浓度胁迫下的响应,探讨钙胁迫处理下番茄幼苗各器官离子分配特征及生理指标变化之间的关系,以期为番茄科学栽培研究提供理论依据。
1 材料与方法
1.1 供试材料 试验材料为加工番茄品种特选87-5,对种子进行消毒和催芽,播种育苗(每隔4d浇灌营养液1次)。选取生长良好、大小一致的幼苗植入花盆(盆土采用珍珠岩与蛭石以2∶1混匀作为培育基质,离子含量为0,每盆定植10株)。
1.2 试验设计 不同浓度的钙营养液处理加工番茄幼苗,Hoagland完全营养液中钙浓度梯度分别为0(缺素)、160mg/L(ck)、320mg/L、640mg/L。以上处理分别是在Hoagland完全营养液中加入CO(NH2)2、KHCO3和CaCl2 。10月2日开始营养液处理,将水培盆和基质栽培花盆放在光照强度3000lx的栽培架,每天观察1次,用pH试纸和pH值仪检测水培养液的pH值,用1%盐酸或氯化钠调整至pH5~6。每隔4d浇灌营养液1次,浇灌量2L,营养液浓度一样。处理21d后取样,测定钾、氮、钙、镁和磷离子的含量。
1.3 测定方法 根、茎、叶样品测定:钙、镁离子采用混合消煮原子吸收分光光度法[13],氮的测定采用奈氏比色法[14-15],磷的测定使用矾钼黄比色法[16-17]。叶绿素含量的测定采用丙酮法[18];根系活力测定采用氯化三苯基四氮唑(TTC)染色法[18]。
1.4 数据处理 试验所有数据作3个重复,用DPS统计软件处理数据,用LSD方法进行方差分析。用Excel 2010软件作图(显著性关系:a,b,c,d;i,j,k,l;q,r,s,t)。
2 结果与分析
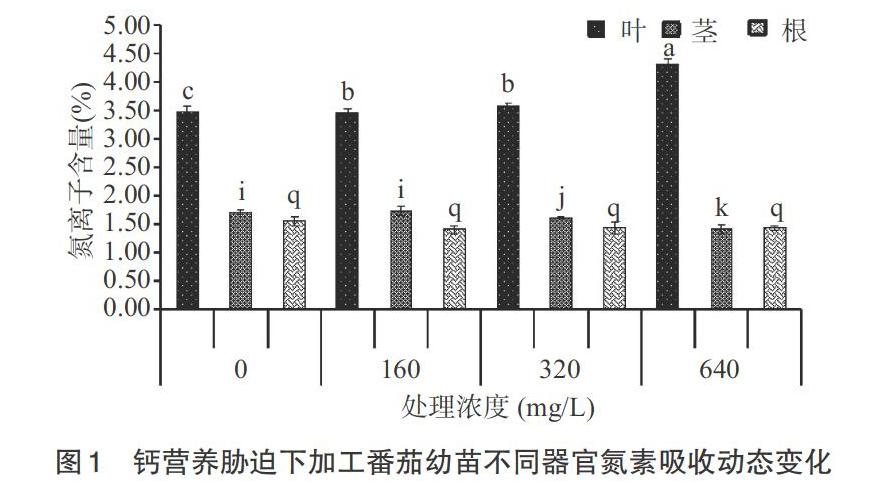
2.1 钙营养元素胁迫下加工番茄幼苗不同器官离子吸收动态
2.1.1 氮素 从图1可以看出,钙营养胁迫下叶片中氮素含量逐渐增高,640mg/L处理时最高,无钙处理最低,对照和320mg/L处理之间存在显著性差异(P<0.05)。茎中氮素含量随着钙营养浓度的提高而减少,0(无钙)和对照处理之间无显著性差异(P>0.05)。根中氮素含量不同浓度处理之间无显著性差异(P>0.05)。整体上,叶片氮素含量最高,根最少。
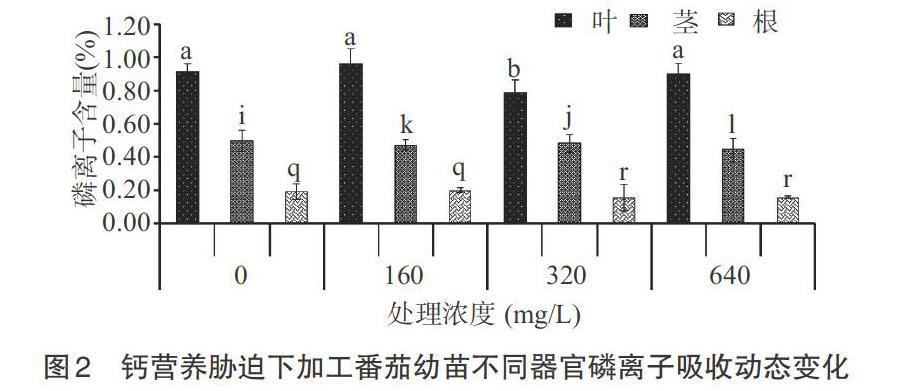
2.1.2 磷离子 从图2可以看出,钙营养胁迫对不同器官磷离子含量的影响比较明显,呈先增高后减少的变化趋势;320mg/L处理各器官磷离子含量最高,0(无钙)和640mg/L处理的最低。叶片中磷离子含量320mg/L处理时最高,640mg/L处理的最低。0(无钙),320mg/L和160mg/L之间无显著性差异(P>0.05)。茎中磷离子含量320mg/L处理时最高,0(无钙),160mg/L和320mg/L处理之间无显著性差异(P>0.05),对照和其他处理之间差异明显。根中磷离子含量变化,与320mg/L相比320mg/L与640mg/L和160mg/L无显著性差异(P>0.05),320与0mg/L处理之间存在显著性差异(P<0.05)。
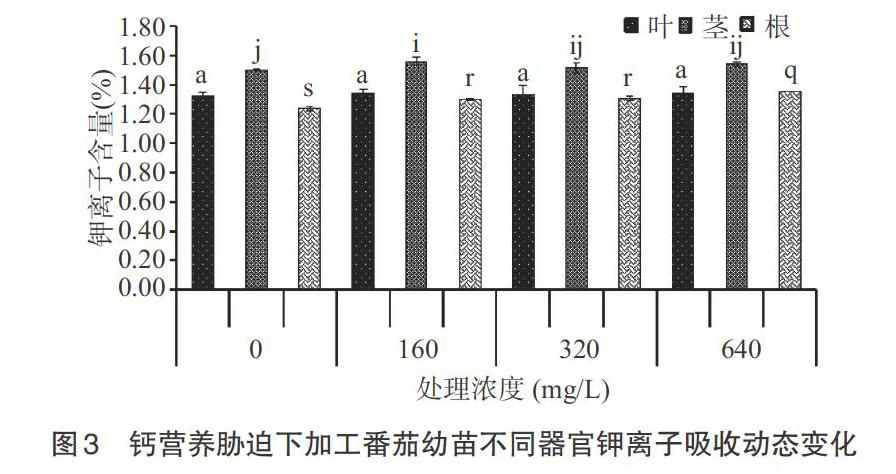
2.1.1 钾离子 从图3可以看出,钙营养胁迫下加工番茄不同器官钾离子含量变化不同。叶片中钾离子含量在不同浓度处理之间无显著性差异(P>0.05)。茎中对照与相比320mg/L和640mg/L处理之间无显著性差异(P>0.05)。随着钙离子浓度的增加,根中钾离子含量呈先上升后下降的趋势。与160mg/L相比320mg/L处理无显著性差异(P>0.05),0mg/L和640mg/L处理存在显著性差异(P<0.05)。整体上,茎中钾离子含量均高于叶片和根中的,根中最低。
2.1.4 钙离子 从图4可以看出,各处理间,叶片中钙离子含量最高,茎次之,根中钙离子含量最低。160mg/L对照处理的叶片和根中钙离子含量显著性高于其他3个处理,其中,对照根中的钙离子含量比其他3个处理根的高于2倍左右。0mg/L浓度钙处理的各个器官的钙离子含量均低于其他3个处理,根中钙离子含量显著性低于其他处理。
2.1.3 镁离子 从图5可以看出,钙营养胁迫对不同器官镁离子含量的影响比较明显,呈先增高后减少的变化趋势;对照各器官镁离子含量最高,640mg/L处理的最低。叶片中镁离子含量对照处理时最高,640mg/L处理的最低。320mg/L和640mg/L处理之间无显著性差异(P>0.05)。茎中镁离子含量对照处理时最高,0(无钙),320mg/L和640mg/L处理之间无显著性差异(P>0.05),对照和其他处理之间差异明显。根中镁离子含量变化和與叶片相同。
2.2 钙营养胁迫对加工番茄幼苗叶绿素含量的影响 从图6可以看出,随着钙营养胁迫的加重,番茄幼苗叶绿素含量呈先逐渐增强后缓慢下降的变化趋势。钙营养浓度从0(无钙)处理增到320mg/L时,幼苗叶绿素含量显著上升,到320mg/L时其叶绿素含量表现最高。160mg/L和320mg/L之间无显著性差异(P>0.05)。0(无钙)处理、640mg/L与160mg/L处理相比存在显著性差异(P<0.05)。
2.3 钙营养胁迫对加工番茄幼苗根系活力的影响 从图7可以看出,随着钙浓度的增加,加工番茄幼苗根系活力呈先逐渐增高后下降的变化趋势。钙营养浓度从0(无钙)处理增到320mg/L时,幼苗根系活力显著上升,到320mg/L时其根系活力表现最强。各钙浓度处理与160mg/L对照相比有显著性差异(P<0.05)。
3 讨论
钙是植物生长必需的营养元素,是构成细胞壁和细胞膜的重要组分,对于维持细胞结构的稳定性,调节无机离子运输等方面起着至关重要的作用[19]。钙元素对植物吸收矿质元素起促进和抑制作用,与植物品种、栽培环境及钙水平有关,机理较复杂且尚不明确。Ca对叶用莴苣吸收N、P的影响较小,而对K的吸收具有双重作用。一方面,适量的Ca可激活质膜上与离子运输有关的酶(ATP酶等),增强膜上K的渗透,进而促进K的吸收;另一方面,由于植物对K的吸收与根系的生长状况有关系,Ca浓度过高抑制了根系的生长,从而减少了K的吸收[20]。李娟等研究发现,K、Ca、Mg之间存在交互作用,但不是单一的拮抗作用,K对烟草吸收Ca、Mg产生抑制作用,Ca对Mg的吸收产生抑制作用,而Ca对K的吸收具有促进作用[21]。本试验结果显示,钙水平过高或过低均会影响其他元素的吸收。不同钙浓度处理下叶片氮磷钙含量均高于茎和根,320、640mg/L钙浓度处理下镁含量均降低,640mg/L钙浓度处理下根和茎镁浓度与对照差异显著。
付嵘等[22]研究发现,钙浓度过高或过低均会对叶绿体的结构和叶绿素a、叶绿素b等含量有不利影响,从而影响光合作用。本试验推测钙浓度亏缺或过剩均影响加工番茄根系活力和各器官的钙镁元素分配,导致叶绿素合成减少。320mg/L钙浓度处理下显著提高了加工番茄幼苗的根系活力,640mg/L鈣浓度处理下叶绿素含量比对照显著降低。在实际生产中,番茄脐腐病的发病率较高,其发病率与果实钙含量有关[23]。为了防治脐腐病,可使用钙肥减轻病害的发生程度。总之,在农业生产中合理施肥不仅能提高产量,还会降低成本和环境污染,达到绿色生产的效果。
参考文献
[1]Hepler P K. Calcium, central regulator of plant growth and development a [J]. Plant cell,2005,17:2152-2155.
[2]曹一平,陆景陵.高等植物的矿质营养[M].北京:北京农业大学出版社,1991:213-218.
[3][5]Dias AS,Lidon FC,Ramalho JC. I. Heat stress in triticum:kinetics of and accumulation [J].Brazilian society of Plant Physiology,2009,21[2]:123-124.
[4]Dodd AN,Kudla J,Sanders D.The language of Calcium signaling [J].Annull Review of Plant Biology,2010,61:620.
[5]White P J ,Broadley M R . Calcium in Plants[J]. Annals of Botany,2003(4):487-511.
[6]HIRSCHI ILD. The calcium conundrum,both versatile nutrient and specific signal[J]. Plant Physiol,2004,136:2438-2442
[7]杨进,靳杏子.切花月季营养元素缺素或过剩现象及其生理意义[J].北方园艺,2014(2):191-195.
[8]杨振.钙对人参生长发育影响的研究[D].北京:中国农业科学院,2015:15-20.
[9]阿不都外力·卡力阿不都,夏依买尔旦·艾白都力,李烨,等.氮、磷、钾、钙营养胁迫对加工番茄幼苗光合色素及光合特性的影响[J].新疆农业科学,2013,50(1):71-76.
[10]张海平.钙调控花生(Arachis hypogaea L.)生长发育的细胞生理机制研究[D].福州:福建农林大学,2003:15-25.
[11]刘卫萍,陈日远,孙光闻,等.不同钙素水平对红葱生长和品质的影响[J].中国蔬菜,2008,1(2):25-27.
[12]Singh K K,Goswami T K.Mechanical properties of cumin seed (Cuminum cyminum Linn.) under compressive loading.[J].Journal of Food Engineering,1998,36(3):311-321.
[13]中国科学院南京土壤研究所.土壤理化分析[M].上海:科学技术出版社,1977(10):73-78.
[14]张行峰.使用农化分析[M].北京:化学工业出版社,2005:193-202.
[15]杜森,高祥.土壤分析技术规范[M].2版.中国农业出版社,2006.
[16]张会民,刘红霞.土壤与植物营养试验实习教程[M].杨凌:西北农业科技大学出版社,2004.
[17]鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,1999:194.
[18]李合生.植物生理生化试验原理与技术[M].北京:高等教育出版社,2000.
[19]ILINZEL H Calcium in the vacuoles and cell walls of plant tissue[J]. Flora,1989.182:99-125
[20]范双喜,伊东正.钙素对叶用莴苣营养吸收和生长发育的影响[J].园艺学报,2002,29(2):149-152.
[21]李娟,章明清,林琼,等.钾、钙、镁交互作用对烤烟生长和养分吸收的影响[J].安徽农业大学学报,2005,32(4):529-533.
[22]付嵘,孟小暇,柴胜丰.植物与钙环境关系的研究进展[J].北方园艺,2019,426(03):167-172.
[23]罗绪强,王世杰,张桂玲,等.钙离子浓度对两种蕨类植物光合作用的影响[J].生态环境学报,2013,22(02):258-262.
(责编:张宏民)
