入侵植物印加孔雀草在不同生境的种群构件生物量及其分配特征
2021-06-10仇晓玉徐知远土艳丽罗建
仇晓玉 徐知远 土艳丽 罗建
摘 要:印加孔雀草是中國西藏新近入侵种,其危害已初见端倪,但我国鲜有对其入侵机理的研究。为探究印加孔雀草在异质环境下的种群构件生物量及其分配特征,进一步深入了解其生存策略和易入侵生境。该研究在菜园、果园、路边、荒地和河滩等五种典型入侵生境内对其花果期的种群构件生物量进行了测定和分析,计算了表型可塑性指标值。结果表明:(1)印加孔雀草种群各构件生物量的基本规律为茎>花果>叶>根,在路边最大,菜园最小,两地间各值均存在显著差异(P<0.05)。(2)印加孔雀草各构件生物量的总变异系数(CV)和可塑性指标(PI)的均值分别为46.93%和61.44%。(3)印加孔雀草繁殖与营养构件生物量比的关系为荒地>路边>果园>河滩>菜园,根冠比表现为菜园>荒地>河滩>路边>果园。(4)印加孔雀草各构件之间、各构件与总生物量间都存在显著的正相关关系,体现了其整体协调的生存策略。上述结果表明,印加孔雀草能通过各构件生物量的调整来适应异质生境,具有较高的可塑性;高繁殖输出和对异质环境的适应性可能是其成功入侵的重要原因。
关键词: 入侵植物, 印加孔雀草, 不同生境, 种群构件, 生物量分配, 中国西藏
中图分类号:Q944.3
文献标识码:A
文章编号:1000-3142(2021)03-0447-09
收稿日期:2020-01-06
基金项目:西藏自治区科技重大专项(ZD20170021);西藏农牧学院西藏高原生态安全联合实验室开放基金 (STX2018-14) [Supported by Science and Technology Great Program of Tibet, China (ZD20170021); Open Fund of United Key Laboratories of Ecological Security of Tibet Autonomous Region, Tibet Agriculture & Animal Husbandry University (STX2018-14)]。
作者简介: 仇晓玉(1991-),硕士研究生,研究方向为高原植物种群与群落生态学,(E-mail)2538343348@qq.com。
通信作者:罗建,硕士,研究员,研究方向为植物分类和生物多样性,(E-mail)luojian-sh@sohu.com。
Module biomass and allocation characteristics of invasive plant Tagetes minuta populations in different habitats
QIU Xiaoyu1,2,3, XU Zhiyuan4, TU Yanli5, LUO Jian1,2,3*
( 1. Research Institute of Tibet Plateau Ecology, Tibet Agriculture & Animal Husbandry University, Nyingchi 860000, Tibet, China; 2. Tibet Key Laboratory of Forest Ecology in Plateau Area, Ministry of Education, Nyingchi 860000, Tibet, China; 3. National Key Station of Field Scientific Observation & Experiment in Nyingchi Tibet, Nyingchi 860000, Tibet, China; 4. Resources & Environment College, Nyingchi 860000,Tibet, China; 5. Tibet Plateau Institute of Biology, Lhasa 850001, China )
Abstract:
Tagetes minuta, as a newly invasive species in Tibet of China, its damage is beginning to appear. However, there are few studies on its invasion mechanism in China. In order to explore the biomass and its allocation characteristics of population modules of T. minuta in heterogeneous habitats, and to further understand its survival strategies and invasive habitats, we determined and analyzed the biomass characteristics and calculated the index values of phenotypic plasticity of the population modules in the flowering and fruiting stages of the T. minuta in five typical invasive habitats, including vegetable garden, orchard, roadside, wasteland and riverside. The results were as follows: (1) The basic law of biomass of module in T. minuta population was stem > flower and fruit > leaf > root. The biomass of each module was the largest in the roadside and the smallest in the vegetable garden, and there were significant differences between these two habitats (P<0.05). (2) The total coefficient of variation (CV) and phenotypic plasticity index (PI), based on maximum and minimum means of each module were 46.93% and 61.44%, respectively. (3) The relationship between biomass ratio of reproductive module and nutrient module of T. minuta was wasteland > roadside > orchard > riverside > vegetable garden, and the root-shoot ratio was vegetable garden > wasteland > riverside > roadside > orchard. (4) There were significantly positive correlation between biomass of different modules, between biomass of different modules and total biomass, which showed its ensemble and consistent strategy of survival. The above results indicate that T. minuta can adapt to heterogeneous habitats by adjusting the biomass of each module, and thus has a strong phenotypic plasticity. High reproductive output and adaptability to heterogeneous environments may be the important reasons for its successful invasion.
Key words: invasive plant, Tagetes minuta, different habitats, population module, biomass allocation, Tibet of China
生物入侵是造成物种濒危及灭绝的重要原因之一(谭勇等, 2014),其中,植物入侵已成为世界上最难应对的三大环境问题之一(郑欣颖和薛立, 2018)。表型可塑性是植物拥有较大生态幅和更高耐受性的重要因素,对于生物的分布有重要意义(陆霞梅等, 2007; Dostál et al., 2016)。与本地植物相比,入侵植物常具有较大的表型可塑性(Thompson, 1991; Godoy et al., 2011),约有50%入侵植物的入侵能力与表型可塑性相关(郑欣颖等, 2018),且同一物种的不同种群之间,表型可塑性也会存在差异(Vilela et al., 2008)。生物量作为表型可塑性表现形式之一,直接影响着植物的生长和繁殖(Weine, 2004),可反映环境对植物的影响程度以及植物对环境的适应情况(周兵等, 2015),其分配方式的转变可造成外来植物入侵扩张(齐淑艳等, 2006)。近年来,许多学者通过植物种群构件生物量及其分配特征展开了对入侵植物表型可塑性的研究(朱慧和马瑞君, 2010; 谭勇等, 2014; 周兵等, 2015),这对于揭示入侵机理具有重要意义。
印加孔雀草(Tagetes minuta)是菊科(Asteraceae)万寿菊属一年生草本植物,原产于南美洲,现今已广泛分布于北美洲、欧洲、亚洲、非洲和大洋洲等20多个国家和地区(张瑞海等, 2019)。该植物在我国台湾、北京、河北、山东和江苏等地已成功定植分布(张瑞海等, 2019),目前是西藏危害大、入侵性较强的外来物种之一(杨春蕾等, 2018)。印加孔雀草具有较强的化感抑制作用(Arora et al., 2016)、抗逆能力强(张瑞海等, 2019)、生态幅宽泛(张劲林等, 2014)、繁殖和传播迅速(张瑞海等, 2019),可在短期内迅速占据生态位并形成其单一优势群落。该植物自2009年在西藏被发现以来(许敏和扎西次仁, 2015),种群数量及分布区域面积均已呈爆发增长的趋势(土艳丽等, 2018),开展印加孔雀草的入侵生物学研究显得尤为迫切。
目前,国内关于印加孔雀草的研究主要集中在归化分布(
董振国等, 2013;张劲林等, 2014;许敏和扎西次仁, 2015)、竞争能力(土艳丽等,2018)、传粉网络(土艳丽等,2019)和风险评估(张瑞海等, 2019)等方面。本研究拟对不同生境印加孔雀草各构件生物量特征进行定量分析,了解其表型可塑性,探讨入侵机理和生态学适应性,预测其最易成灾生境,为防控提供依据。
1 材料与方法
1.1 研究区概况
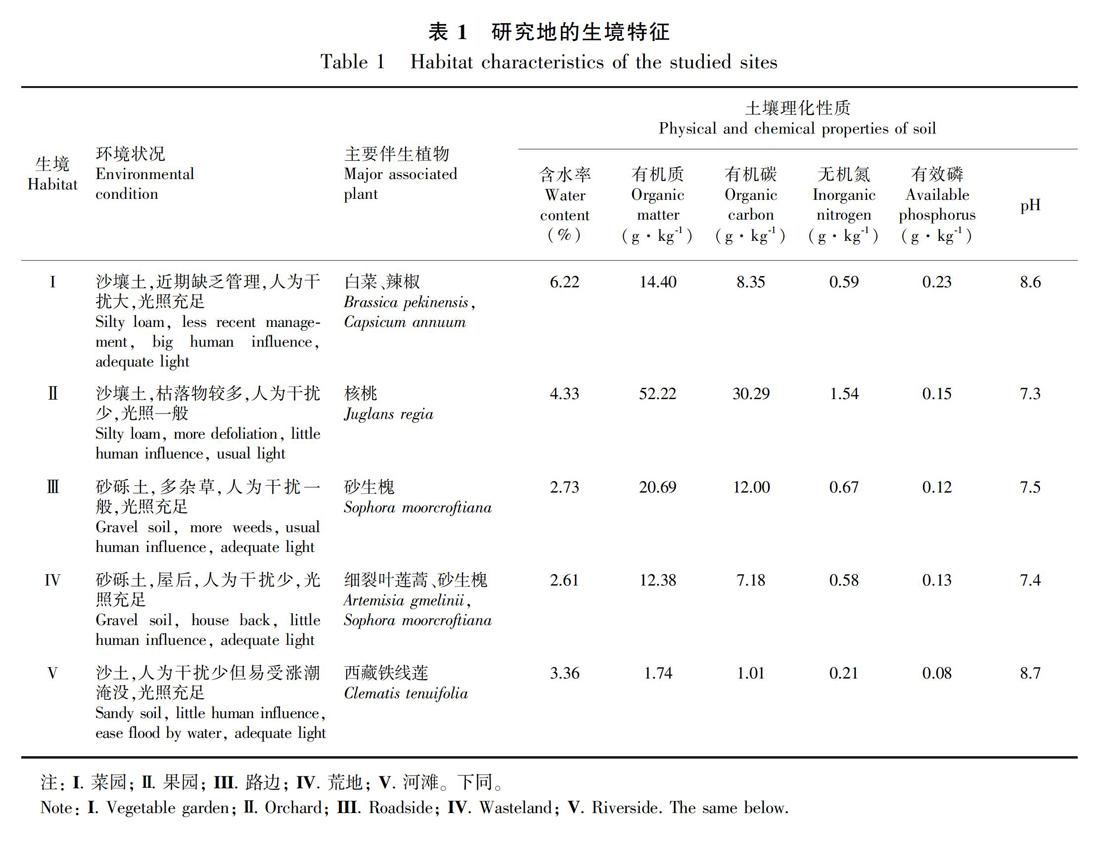
研究区域位于西藏东南部,在林芝市米林县至朗县地区(92°28′—95°12′ E、28°39′—29°50′ N,海拔3 200~3 700 m),印加孔雀草入侵较为严重。该地属高原温带半湿润季风气候,年降雨量为350~641 mm,年均温度为8.2~11.0 ℃,年均日照为2 000~2 500 h,无霜期为130~170 d。
1.2 研究方法
1.2.1 调查取样 2018年10月,于印加孔雀草花果期,选择菜园、果园、路边、荒地和河滩五种典型生境(表1)分别调查取样。印加孔雀草在各生境和样方内均为优势种,在各样方内盖度达80%以上,密度约为每平方米179株。在各生境内分别选取3个1 m × 1 m的样方,样方间距离大于5 m且小于10 m。调查每个样方中印加孔雀草的密度,随机选取10个完整植株测量株高,并将其所有根系及0~10 cm层土样一起挖出,同样方内的土壤混合在一起,同一样方内土样测定重复3次。植株洗净根部后,根、茎、叶和花果分开,分别置于信封内,在烘箱内105 ℃下杀青20 min,然后70 ℃烘至恒重,用1/10000电子天平称重。土壤有机质(有机C)、无机N和有效P的测定分别采用重铬酸钾浓硫酸氧化法、KCl浸提-连续流动分析仪法和NaHCO浸提-钼锑抗比色法,土壤pH值采用酸度计测定(浸提液水土质量比2.5∶1)。
1.2.2 数据处理 采用Excel软件进行数据处理及回归分析,在SPSS 19.0软件中分别运用Pearson相关分析方法和单因素方差分析(one-way ANOVA)分析株高与各构件生物量等数据的相关性和差异显著性(P<0.05),采用LSD法进行多重比较,利用Origin 2018软件作图。把印加孔雀草所有构件称重后,将平均生物量值带入以下公式,求出表型可塑性表达公式,即总变异系数(CV)和基于最大平均值和最小平均值的可塑性指数(PI)(Valladares et al., 2006;王姝和周道玮, 2017)。本研究用CV和PI的平均值表示印加孔雀草的表型可塑性值。
CV= (标准差/平均值)×100%(1)
PI =(最大平均值-最小平均值)/最大平均值×100%(2)
式中:最大平均值和最小平均值是指植物生物量分别在不同生境的各重复样方间的平均值,数值较大的平均值即为最大平均值,较小的即为最小平均值。
2 结果与分析
2.1 不同生境印加孔雀草種群构件生物量及分配比特征
如表2所示,在各生境下,茎生物量的分布为路边>果园>荒地>河滩>菜园,其余构件均为路边>荒地>果园>河滩>菜园。植株总量及各构件生物量以路边最大,菜园最小,根、茎、叶、花果构件平均生物量及总量分别占路边值的2.71%、1.68%、4.14%、1.36%和2.14%。其中:路边与其他生境均存在显著差异;菜园与河滩无显著差异,而与其他生境存在显著差异;果园与荒地无显著差异,而与其他生境存在显著差异(P<0.05)。
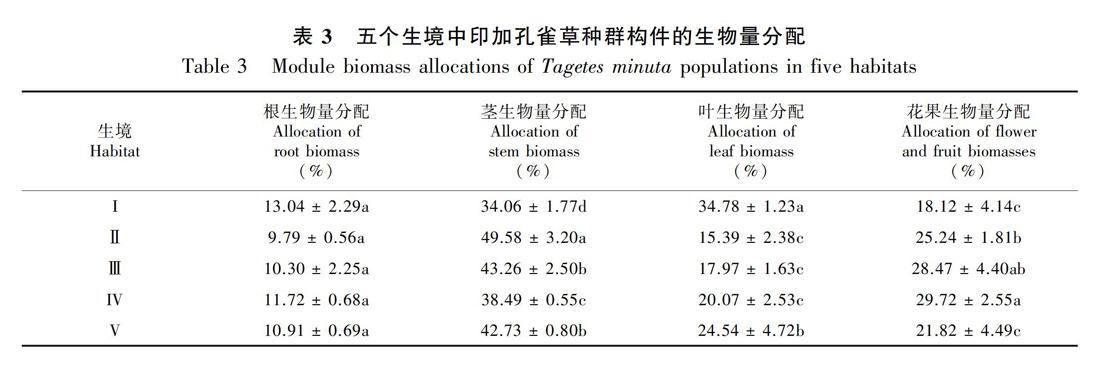
由表3可知,印加孔雀草构件生物量分配比在各生境有相似规律:茎最大,根最小。其中:在果园、路边和荒地生境下,各构件生物量分配比为茎>花果>叶>根;在菜园为叶>茎>花果>根;在河滩则为茎>叶>花果>根。其中:根在各生境无显著差异;茎在路边和河滩无显著差异,其他生境下呈显著差异;叶在菜园和河滩分别与其他生境存在显著差异;花果生物量除路边和荒地外,其余各生境下均存在显著差异(P<0.05)。
2.2 不同生境印加孔雀草种群构件表型可塑性特征
由表4可知,不同生境中,印加孔雀草各构件和全株的表型可塑性值存在较大差异,在河边生境最大,路边最小;各构件中花果的表型可塑性值最高(59.66%)。各生境中印加孔雀草构件生物量的CV平均值为46.93%,其中:根、茎、叶、花果和全株生物量依次为41.69%、45.25%、48.71%、52.09%和46.43%;PI平均值为61.44%,根、茎、叶、花果和全株生物量依次为55.93%、60.51%、61.60%、67.22%和62.28%。
2.3 不同生境印加孔雀草种群繁殖与营养构件的生物量比
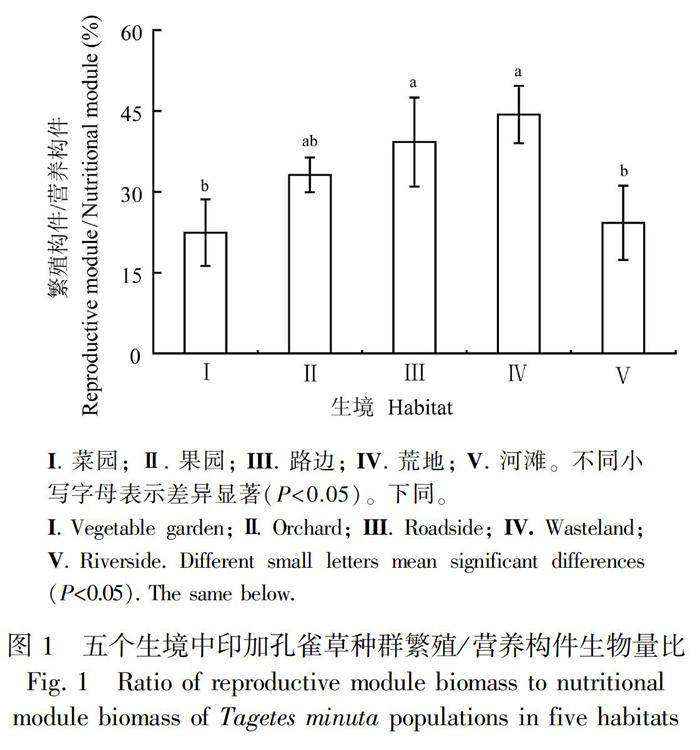
由图1可知,印加孔雀草种群繁殖构件(花果)与营养构件(根茎叶)生物量比值在不同生境下由大到小依次为荒地、路边、果园、河滩和菜园,其值分别为42.28%、39.80%、33.75%、27.91%和22.12%。果园、路边和荒地无显著差異,菜地、果园和河边无显著差异,而路边和荒地均与菜园和河边存在显著差异,且最大值是最小值的1.91倍,差异显著(P<0.05)。
2.4 不同生境印加孔雀草种群的根冠比
如图2所示,印加孔雀草种群根冠比在五种生境下分布规律为菜园>荒地>河滩>路边>果园,其值分别为15.49%、13.60%、11.88%、11.85%和11.01%。除荒地外,菜园与其他生境值均存在显著差异(P<0.05),果园、路边、荒地和河滩无显著差异(P<0.05)。
2.5 不同生境土壤理化性状对印加孔雀草生物量分配的影响
对各生境印加孔雀草构件生物量与对应群落内土壤含水率、C、N、P含量以及pH值进行Pearson相关性分析后发现,印加孔雀草各构件生物量仅与含水率和pH值间呈显著负相关(表5),这表明印加孔雀草各构件生物量与土壤元素间的相关性较弱,含水率与pH值可能是制约印加孔雀草生物量分配的主要因素。
2.6 印加孔雀草种群各构件生物量之间关系
五种生境中印加孔雀草的根、茎、叶、花果的生物量间以及与总生物量间都存在显著的正相关关系(P<0.01),对各构件生物量及总生物量的数据分别进行汇总后发现结果仍为显著正相关关系(P<0.01)(表6)。各构件生物量均与总生物量间的相关性最强,R在0.977~0.995间;茎与总生物量间的相关系数最大(0.995);根与花果生物量的相关性最小(0.951),但仍大于0.950。
3 讨论与结论
3.1 印加孔雀草种群各构件生物量及分配特征
印加孔雀草种群各构件生物量因不同生境而存在差异,表明生境综合条件是其影响因素,也表明该物种的生物量积累具较大的可调节性和表型可塑性。印加孔雀草各构件生物量在路边最大、菜园最小,这可能与生境内土壤肥力、光照以及人为干扰等因素有关。路边无高大乔木遮阴,光照条件好,空气流通性好,土壤养分相对较高,故其生物量最大。而菜园为半自然状态,长期种植蔬菜却施肥较少,有机碳含量仅为8.35 g·kg-1,养分不足限制了印加孔雀草的物质积累;此外,菜地内相对采取栽培措施,人工物理化学除草较多,印加孔雀草高大,植株易被拔除。
不同生境印加孔雀草种群构件生物量分配的异同,体现了其具有重要的物质分配策略和相对稳定的分配模式。在菜地,人为清除抑制了印加孔雀草株高的优势,促使其把更多能量倾向于光合构件叶的建造上,以保证足够的物质生产;在其他生境则均表现为优先供应茎生长,茎生物量较大有利于其获得空间和光照等资源,提高竞争力和入侵能力。入侵植物在入侵地常具更强繁殖力(Marcia et al., 2017),这也是其快速定殖和扩散的重要因素(闫小红等, 2017)。印加孔雀草在各生境花果生物量分配比为18.12%~29.72%,显著高于同为菊科的较强入侵性植物胜红蓟(Ageratum conyzoides)(3.19%~6.20%)(周兵等, 2015),这有利于其种群稳定发展,大狼把草(Bidens frondosa)(周兵等, 2012)、薇甘菊(Mikania micrantha)(徐高峰等, 2014)和3种苋科(Amaranthaceae)植物(项剑桥等, 2017)等也存在相似的分配规律。印加孔雀草种群各构件生物量根据环境做出了不同分配响应并成功定植繁殖,该适应机制对于拓宽其生态幅、增强入侵性具有重要意义。
3.2 印加孔雀草表型可塑性特征
入侵种群间表型可塑性的差异表明,入侵种群具有可塑性进化的潜力,这可能是其优势的原因(Hiatt & Flory, 2020)。目前,人们对可塑性量化方面的问题还缺少共识(王姝和周道玮, 2017),但本研究中,印加孔雀草大多数构件生物量在不同生境存在显著差异,且其CV与同为菊科的具有较大表型可塑性的蒲公英(Taraxacum mongolicum)(张丽辉等, 2017)和入侵植物钻形紫菀相近(潘玉梅等, 2010),表明印加孔雀草具较强的表型可塑性。在定殖、种群建立、潜伏与扩散传播的过程中,外来入侵植物的表型可塑性常通过适应机制拓宽生态幅,增强其入侵性(宫伟娜等, 2009), 所以, 印加孔雀草的表型可塑性可加强其对异质环境的适应性和耐受性,在一定程度上增加了其入侵能力。此外,印加孔雀草各构件中花果的表型可塑性最高,这更加有利于其繁殖,为其扩散入侵至新生境提供了条件。因此,表型可塑性强可能是其成功入侵的重要因素之一。
3.3 印加孔雀草种群繁殖与营养构件的生物量比、根冠比特征
印加孔雀草繁殖与营养构件生物量比表现为荒地>路边>果园>河滩>菜园,表明在各生境下该比值具可调节性,在土壤C、N和P等养分含量相对较低时(荒地),繁殖器官分配比(29.72%)和繁殖/营养构件比值(42.28%)最高,表明在荒地繁殖能力最强。这可能是受资源限制,印加孔雀草在对营养器官具足够的投入以保证其存活后,将大部分资源投入繁殖器官以促進其繁殖扩散。
根冠比可直接体现植物生物量在地下与地上间分配的协调状况,对增加植株总产量以及个体生长具有重要意义(Enquist & Niklas, 2002;Hui & Jackson, 2006)。印加孔雀草的根冠比在菜园最大,这可能与含水率、氮以及土壤透气性有关。菜园含水率值为6.22%,处于干旱胁迫状态,且氮素含量较少,仅为0.59g·kg-1,植物根系会先满足自身的水分和氮素需要,则冠部供应量会相对减少,地上部分生长受到抑制,所以根冠比会增加(李清, 2019;邹升等, 2019);这也与菜地常被翻耕,土壤较疏松且透气性好而有利于根系生长有关。根冠比的可调节性在狗尾草等(贾风勤等, 2014)植物研究中也得到了验证。印加孔雀草可以合理地分配水分和养分元素等生存资源,对根系和地上生物量进行适时调节的能力,可有效促进其生态适应性,是其生存策略之一。
3.4 印加孔雀草各构件间关系特征
植物生长与物质分配策略会根据自身需求而调整,但也遵循相协调原则(杨允菲和李建东, 2003)。尽管不同生境中印加孔雀草各构件及分配比都存在差异,但其各构件生物量间以及与总生物量间均存在显著的正相关关系(P<0.01),相关系数均大于0.951。入侵植物牛膝菊(Galinsogaparviflora)(齐淑艳等, 2006)、小飞蓬(Conyza canadensis)(潘玉梅等, 2009)和小花山桃草(Gaura parviflora)(刘龙昌等, 2012)等各构件生物量间也表现出相似的相关性,但相关系数均小于印加孔雀草,这表明印加孔雀草各构件间具更紧密的联系,在结构和功能上相互依存和制约,整体协调的生存策略有利于提高其对环境的适应能力,增强其入侵性。
综上所述,五种生境中印加孔雀草各构件生物量的分配规律为茎>花果>叶>根,且其各构件间及与总生物量间存在极显著正相关(P<0.01),表明了其具有相对稳定性和整体协调的生存策略;印加孔雀草在异质环境下通过可塑性调节,在路边生物量积累最好,在荒地生境产种量最高,这两种生境是印加孔雀草最易入侵成灾之地,应加强对两地的监控与管理;较高的总变异系数值(CV)和可塑性指标(PI)均体现了其具有较高的表型可塑性。总之,印加孔雀草种群为适应异质环境而对其各构件生物量做出了优化配置,且其花果的生物量较高以及较强表型可塑性增加了其环境适应性,这可能是其成功入侵的原因。
参考文献:
ARORA K, BATISH D, SINGH HP, et al., 2016. Comparative account of allelopathic potential of essential oil of Tagetes minuta L. and its major component cis-β-Ocimene [J]. Ann Plant Sci, 5(9): 1428-1431.
DOSTL P, FISCHER M, PRATI D, 2016. Phenotypic plasticity is a negative, though weak, predictor of the commonness of 105 grassland species [J]. Global Ecol Biogeogr, 25 : 464-474.
DONG ZG, LIU QX, HU J, et al., 2013. New records of naturalized plants from the Chinese Mainland [J]. Guihaia, 33(3): 432-434. [董振国, 刘启新, 胡君, 等, 2013. 中国大陆归化植物新记录 [J]. 广西植物, 33(3): 432-434.]
ENQUIST BJ, NIKLAS KJ, 2002. Global allocation rules for patterns of biomass partitioning in seed plants [J]. Science, 295(5559): 1517-1520.
GODOY O, VALLADARES F, CASTRO DEZ P, 2011. Multispecies comparison reveals that invasive and native plants differ in their traits but not in their plasticity [J]. Funct Ecol, 25: 1248-1259.
GONG WN, WAN FH, XIE BY, et al., 2009. Phenotypic plasticity and adaptability of the invasive alien species [J]. Plant Protect, 35(4): 1-7. [宫伟娜, 万方浩, 谢丙炎, 等, 2009. 表型可塑性与外来入侵植物的适应性 [J]. 植物保护, 35(4): 1-7.]
HULME PE, 2008. Phenotypic plasticity and plant invasions: Is it all jack? [J]. Funct Ecol, 22: 3-7.
HUI DF, JACKSON RB, 2006. Geographical and interannual variability in biomass partitioning in grassland ecosystems: A synthesis of field data [J]. New Phytol, 169(1): 85-93.
HIATT D, FLORY SL, 2020. Populations of a widespread invader and co-occurring native species vary in phenotypic plasticity [J]. New Phytol, 225(1):584-594.
JIA FQ, ZHANG N, NUERMAIMAITI, et al., 2014. Biomass structure and growth of reproductive ramets of Setaria viridis (L.) Beauv and S. glauca (L.) Beauv [J]. Chin J Ecol, 33(1): 48-52. [贾风勤, 张娜, 努尔买买提, 等, 2014. 狗尾草和金色狗尾草种群生殖分株生物量结构及生长 [J]. 生态学杂志, 33(1): 48-52.]
LU XM, ZHOU CF, AN SQ, et al., 2007. Phenotypic plasticity, allometry and invasiveness of plants [J]. Chin J Ecol, 26(9): 1438-1444. [陆霞梅, 周长芳, 安树青, 等, 2007. 植物的表型可塑性、异速生长及其入侵能力 [J]. 生态学杂志, 26(9): 1438-1444.]
LIU LC, FAN WJ, DONG LM, et al., 2012. Module biomass structure and seed germination traits of the exotic invasive plant Gaura parviflora [J]. Guihaia, 32(1): 69-76. [刘龙昌, 范伟杰, 董雷鸣, 等, 2012. 入侵植物小花山桃草种群构件生物量结构及种子萌发特征 [J]. 广西植物,32(1): 69-76.]
LI Q, 2019. Preliminary study on root-crown ratio of main green manure [J]. Forum S Chin, 50(13): 23. [李清, 2019. 主要綠肥根冠比的初步研究 [J]. 南方农机, 50(13): 23.]
MARCIA GT, QUIROZ CL,BlOOMFIELD IC, et al., 2017. Enhanced fitness and greater herbivore resistance: Implications for dandelion invasion in an alpine habitat [J]. Biol Invasions, 19: 647-653.
PAN YM, TANG SC, CEN YX, et al., 2010. Biomass allocation on the modules of Aster subulatus Michx. population at flowering stage [J]. J Trop Subtrop Bot, 18(2):176-181. [潘玉梅, 唐赛春, 岑艳喜, 等, 2010. 钻形紫菀开花期种群构件的生物量分配 [J]. 热带亚热带植物学报, 18(2): 176-181.]
PAN YM, TANG SC, PU GZ, et al., 2009. Module biomass structure of the alien invasive plant Conyza canadensis [J]. Guihaia, 29(4): 476-480. [潘玉梅, 唐赛春, 蒲高忠, 等, 2009. 外来入侵植物小飞蓬种群构件生物量结构特征 [J]. 广西植物, 29(4): 476-480.]
PETR D, MARKUS F, DANIEL P, 2016. Phenotypic plasticity is a negative, though weak, predictor of the commonness of 105 grassland species [J]. Global Ecol Biogeogr, 25: 464-474.
QI SY, XU WD, WEN Y, 2006. Biomass structure of exotic invasive plant Galinsona parviflora [J]. Chin J Appl Ecol, (12): 2283-2286. [齐淑艳, 徐文铎, 文言, 2006. 外来入侵植物牛膝菊种群构件生物量结构 [J]. 应用生态学报, (12): 2283-2286.]
TAN Y, HE JD, YOU WB, et al., 2014. Spartina alterniflora morphological plasticity and biomass allocation in response to tidal gradient change in Mindong coastal wetlands [J]. Chin J Appl Environ Biol, 20(5): 863-868. [谭勇, 何东进, 游巍斌, 等, 2014. 闽东滨海湿地入侵种互花米草表型可塑性及生物量分配对潮位梯度的响应 [J]. 应用与环境生物学报, 20(5): 863-868.]
TU YL, QIU XY, LUO J, et al., 2018. Competitive effect between invasive plant Tagetes minuta L. and crop Hordeum vulgare L. var. trifurcatum in Tibet Nyingchi [J]. Tib Sci Technol, (11): 62-65. [土艳丽, 仇晓玉, 罗建, 等, 2018. 西藏林芝入侵植物印加孔雀草与农作物藏青稞的竞争研究 [J]. 西藏科技, (11): 62-65.]
TU YL, WANG LP, WANG XL, et al., 2019. Status of invasive plants on local pollination networks: A case study of Tagetes minuta in Tibet based on pollen grains from pollinators [J]. Biodivers Sci, 27(3): 306-313. [土艳丽, 王力平, 王喜龙, 等, 2019. 利用昆虫携带的花粉初探西藏入侵植物印加孔雀草在当地传粉网络中的地位 [J]. 生物多样性, 27(3): 306-313.]
THOMPSON JD, 1991. The biology of an invasive plant: what makes Spartina anglica so successful ? [J]. Bioscience, 41: 393-401.
VILELA A, GONZLEZ-PALEO L, RONDANINI D, et al., 2008. Biomass allocation patterns and reproductive output of four Oenothera L. accessions native to Argentina [J]. Ind Crops Products, 27(3): 249-256.
VALLADARES F, SANCHEZGOMEZ D, ZAVALA MA, 2006. Quantitative estimation of phenotypic plasticity: Bridging the gap between the evolutionary concept and its ecological applications [J]. J Ecol, 94: 1103-1116.
WANG S, ZHOU DW, 2017. Research on phenotypic plasticity in plants: An overview of history, current status, and development trends [J]. Acta Ecol Sin, 37(24): 8161-8169. [王姝,周道瑋, 2017. 植物表型可塑性研究进展 [J]. 生态学报, 37(24): 8161-8169.]
WEINER J, 2004. Allocation, plasticity and allometry in plants [J]. Perspect Plant Ecol Evol Syst, 6(4): 207-215.
XU M, TASHI T, 2015. A newly naturalized plant in Qinghai-Tibetan Plateau [J]. Guihaia, 35(4): 554-555. [许敏, 扎西次仁, 2015. 青藏高原一新归化种 [J]. 广西植物,35(4): 554-555.]
XU GF, SHEN SC, ZHANG FD, 2014. Adaptability andreproductive characteristics of Mikania micrantha H.B.K under different habitats [J]. Ecol Environ Sci, 23(8):1258-1264. [徐高峰, 申时才, 张付斗, 2014. 异质环境下入侵植物薇甘菊的适应性与繁殖特性 [J]. 生态环境学报,23(8): 1258-1264.]
XIANG JQ, DAI GZ, WU DM, et al., 2017. Relationship between soil total soil microelement with soil organic carbon and pH in Xuanen county of Hubei Province: Taking example of Xiao Guan Dong-minority Township [J]. SW Chin J Agric Sci, 30(8): 1849-1853. [项剑桥, 戴光忠, 吴冬妹, 等, 2017. 湖北宣恩县土壤全量微量元素与有机碳和pH的关系分析—以晓关侗族乡为例 [J]. 西南农业学报, 30(8): 1849-1853.]
YAN XH, HE CL, ZHOU B, et al., 2017. Biomass distribution and allometric analysis of Bidens frondosa relative to growth stage [J]. J Ecol Rural Env, 33(2): 150-158. [闫小红, 何春兰, 周兵, 等, 2017. 不同生育期入侵植物大狼把草的生物量分配格局及异速生长分析 [J]. 生态与农村环境学报, 33(2): 150-158.]
YANG CL, LUO J, LAN JJ, 2018. Index system for risk assessment of alien plant invasion in Nyingchi, Tibet autonomous region, China [J]. J Plateau Agric, 2(2): 167-175. [杨春蕾, 罗建, 拦继酒, 2018. 西藏林芝市外来植物入侵风险评估指标体系 [J]. 高原农业, 2(2): 167-175.]
YANG YF, LI JD, 2003. Biomass allocation and growth analysis on the ramets of Phragmites communis populations in different habitats in the Songnen Plains of China [J]. Chin J Appl Ecol, (1): 30-34. [杨允菲, 李建东, 2003. 松嫩平原不同生境芦苇种群分株的生物量分配与生长分析 [J]. 应用生态学报, (1): 30-34.]
ZHU H, MA RJ, 2010. Modulebiomass structures of Amaran thaceous invasive plant populations in East Guangdong [J]. SW Chin J Agric Sci, 23(3): 876-880. [朱慧, 马瑞君, 2010. 粤东地區苋科入侵植物种群的构件生物量结构 [J]. 西南农业学报, 23(3): 876-880.]
ZHOU B, YAN XH, XIAO YA, et al., 2015. Module biomass of Ageratum conyzoides populations in different habitats [J]. Acta Ecol Sin, 35(8): 2602-2608. [周兵, 闫小红, 肖宜安, 等, 2015. 不同生境下入侵植物胜红蓟种群构件生物量分配特性 [J]. 生态学报, 35(8): 2602-2608.]
ZHENG XY, XUE L, 2018. Research progress about phenotypic plasticity of exotic invasive species Bidens pilosa and a congeneic native species B. biternate [J]. Chin J Ecol, 37(2): 580-587. [郑欣颖, 薛立, 2018. 入侵植物三叶鬼针草与近缘本地种金盏银盘的可塑性研究进展 [J]. 生态学杂志, 37(2): 580-587.]
ZHANG RH, ZANG GL, SONG Z, et al., 2019. The invasive assessment and management measure of Tagetes minuta L. [J]. J Biosafety, 28(1): 71-75. [张瑞海, 张国良, 宋振, 等, 2019. 印加孔雀草入侵生态风险评估及其管理措施 [J]. 生物安全学报, 28(1): 71-75.]
ZHANG JL, L YF, BIAN Y, et al., 2014. A new kind of invasive plant from mainland China-Tagetes minuta L. [J]. Plant Quar, 28(2): 65-67. [张劲林, 吕玉峰, 边勇, 等, 2014. 中国境内(内地)一种新的入侵植物—印加孔雀草 [J]. 植物检疫, 28(2): 65-67.]
ZOU S, WANG JC, CHEN H, et al., 2019. Effects of water and nitrogen management under drip irrigation on root crown growth and yield of spring wheat [J]. Jiangsu Agric Sci, 47(12): 129-133. [邹升, 王冀川, 陈 慧, 等, 2019. 滴灌水氮运筹对春小麦根冠生长及产量的影响 [J]. 江苏农业科学, 47(12): 129-133.]
ZHOU B, YAN XH, XIAO YA, et al., 2012. Module biomass structure traits of the alien invasive Bidens frondosa population [J]. Guihaia, 32(5): 650-655. [周兵, 闫小红, 肖宜安, 等, 2012. 外来入侵植物大狼把草种群构件生物量结构研究 [J]. 广西植物, 32(5): 650-655.]
ZHANG LH, NI XZ, TANG QL, 2017. Biomass structure and allometric analysis on the modules of Taraxacum mongolicum population at flowering stage [J]. J Weed Sci, 35(1): 20-24. [张丽辉, 倪秀珍, 汤庆莲, 2017. 蒲公英花期种群构件的生物量结构与异速生长分析 [J]. 杂草学报,35(1): 20-24.]
(责任编辑 周翠鸣)
