西双版纳热带雨林和热带山地常绿阔叶林附生苔藓的组成与多样性格局
2021-06-10全东丽宋亮沈婷吴毅李苏卢华正
全东丽 宋亮 沈婷 吴毅 李苏 卢华正
摘 要:為探讨西双版纳热带雨林和热带山地常绿阔叶林附生苔藓的组成、多样性格局及其维持机制,该文对两种植被的树干附生苔藓植物进行了调查与分析。所选择的样地位于西双版纳勐腊县,每种植被类型选择10个20 m × 20 m的样方,每个样方选择约10棵样树。结果表明:(1)本调查共记录到附生苔藓20科39属60种,其中热带雨林附生苔藓有19科33属48种,热带山地常绿阔叶林附生苔藓有9科14属19种,共有种6种。(2)热带雨林最优势科为平藓科,热带山地常绿阔叶林最优势科为锦藓科。(3)热带雨林比热带山地常绿阔叶林具有更高的物种丰富度、β多样性和γ多样性。(4)不同径级和不同树皮粗糙度宿主上附生苔藓植物的盖度差异显著,而物种丰富度差异不显著。(5)苔藓植物的生活型对植被类型有所偏好,表现为悬垂型、扇型、交织型、粗平铺型集聚于热带雨林,丛集型、垫状型、细平铺型集聚于热带山地常绿阔叶林。(6)直接排序结果表明,宿主特性特别是树皮粗糙度显著影响苔藓植物生活型的组成和分布。热带雨林比热带山地常绿阔叶林能提供更多样的微生境,因而孕育了物种更丰富的附生苔藓植物。考虑到不同的植被类型或宿主特性条件下孕育着不同生活型组成的苔藓植物,因此可将苔藓生活型作为今后森林监测的一项重要指标。
关键词: 西双版纳, 生物多样性, 附生植物, 生活型, 分布格局
中图分类号:Q948
文献标识码:A
文章编号:1000-3142(2021)03-0404-14
收稿日期:2020-03-22
基金项目:中国科学院‘一三五战略规划专项项目(2017XTBG-F03, 2017XTBG-F01);国家自然科学基金面上项目(31670452);澜沧江-湄公河合作专项基金项目; 中国科学院“西部之光”人才培养引进计划 [Supported by the 135 Program of CAS (2017XTBG-F03, 2017XTBG-F01); the National Natural Science Foundation of China (31670452); Lancang-Mekong Cooperation (LMC) Special Fund; CAS “Light of West China” Program]。
作者简介: 全东丽(1996-),硕士研究生,主要研究方向为生物多样性与保护,(E-mail)quandongli@xtbg.ac.cn。
通信作者:宋亮,博士,副研究员,研究方向为植物生态、恢复生态,(E-mail)songliang@xtbg.ac.cn。
Composition and diversity pattern of epiphytic bryophytes
in tropical rainforest and tropical montane evergreen broad-leaved forest in Xishuangbanna
QUAN Dongli1,2, SONG Liang1,3*, SHEN Ting4, WU Yi1, LI Su1,3, LU Huazheng1,3
( 1. CAS Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Menglun 666303, Yunnan, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Core Botanical Gardens, Chinese Academy of Sciences, Menglun 666303, Yunnan, China; 4. Institute of Botany, University of Liege, Sart Tilman 4000 Liege, Belgium )
Abstract:This study explored the bole epiphytic bryophytes in the tropical rainforest and the tropical montane evergreen broad-leaved forest in Xishuangbanna, aimed to reveal their composition, diversity patterns, and maintenance mechanisms. The study site is located in Mengla County. Ten sampling plots (20 m × 20 m) of each vegetation type were chosen, and about ten tree individuals were sampled in each plot to survey the bole epiphytic bryophytes. The results were as follows: (1) In total, 60 species belonging to 39 genera and 20 families were recorded. Among them, 48 species belonging to 33 genera and 19 families were recorded in the tropical rainforest, and 19 species belonging to 14 genera and 9 families were recorded in the tropical montane evergreen broad-leaved forest. (2) The dominant family of the tropical rainforest is Neckeraceae, while the dominant family of the tropical montane evergreen broad-leaved forest is Sematophyllaceae. (3) The species richness, β and γ diversities of the tropical rainforest were higher than the tropical montane evergreen broad-leaved forest. (4) The coverage of epiphytic bryophytes was significantly different among different diameter classes and bark roughness. (5) Pendants, fans, wefts, and rough mats prefer the tropical rainforest, while turfs, cushions, smooth mats gather in the tropical montane evergreen broad-leaved forest. (6) The results of redundancy analysis showed that host characteristics, especially bark roughness, significantly affected the composition and distribution of life forms. Tropical rainforest provide a greater diversity of microhabitats than tropical montane evergreen broad-leaved forest, and thus harbor more epiphytic bryophyte species than the latter. Considering different bryophyte life form were recorded in different vegetation types or host characteristics, life forms can be considered as an important indicator for forest monitoring and management.
Key words: Xishuangbanna, diversity, epiphyte, life form, distribution pattern
苔藓植物是4.7亿至5.5亿年前从水生藻类祖先绿藻成功登陆的首批绿色植物之一(Morris et al., 2018)。就物种多样性而言,苔藓植物是仅次于被子植物的第二大高等植物类群,是生物多样性的重要组成部分(Crosby et al., 2000)。虽然与绿藻相比,苔藓植物在形态结构和繁殖特性方面有了显著的进化。然而,由于没有根的分化(Ligrone et al., 2008)、主要通过配子体表面吸收水分和养分、有性生殖过程必须依赖于水(Steinhorst & Kudla, 2017)、变水(poikilohydry)等特性,苔藓植物对气候条件和环境变化非常敏感(Azuelo et al., 2011; Song et al., 2012; He et al., 2016; Shi et al., 2017)。而生物应对环境变化的能力因物种而异(Willis & MacDonald, 2011; Jaeschke et al., 2013; Bocedi et al., 2014)。栖息地破碎化和气候变化的综合影响可能导致群落中物种组成和多样性的实质性变化(Mantyka-Pringle et al., 2015)。在这样的背景下,苔藓植物的群落组成可能发生明显变化(Frego, 2007; Sporn et al., 2010; Lbel et al., 2018)。
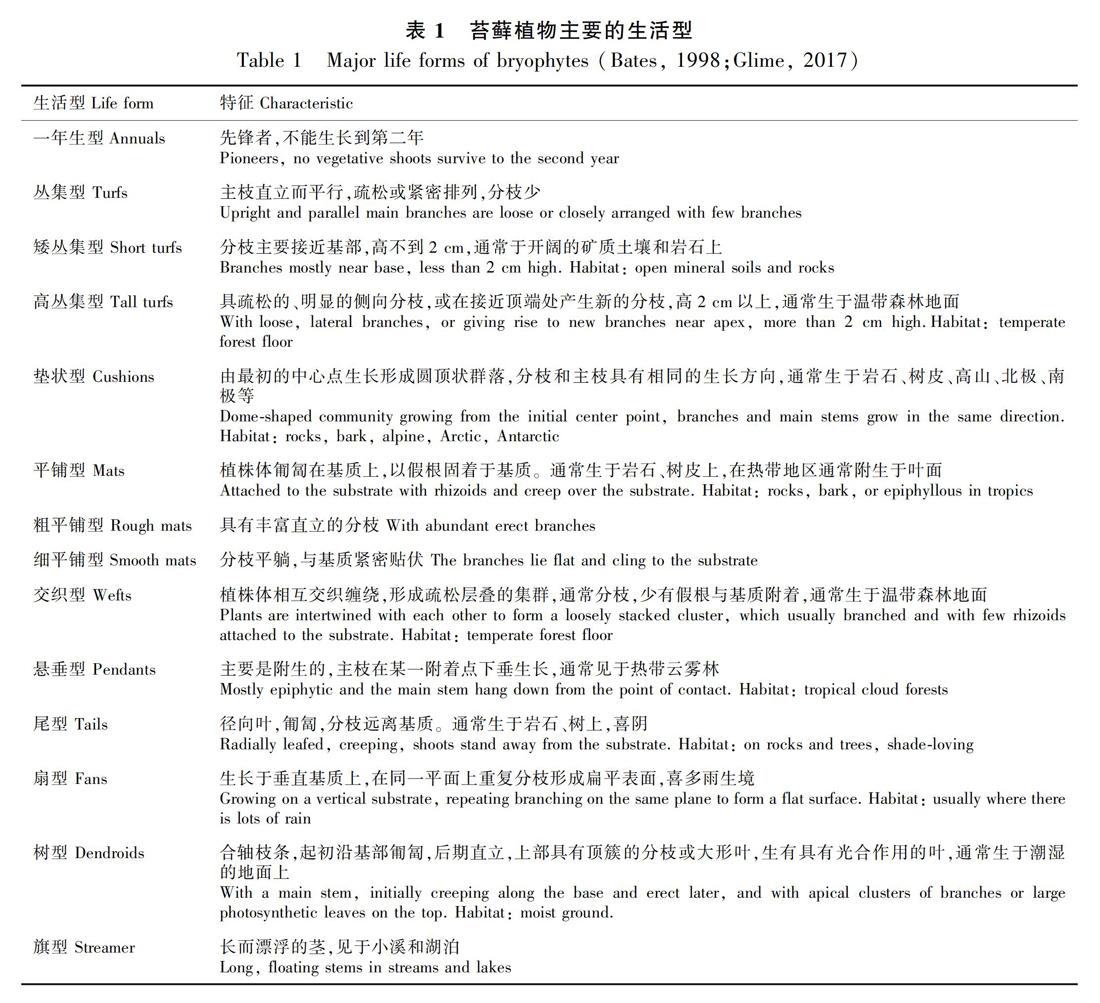
苔藓植物生活型反映了其对特定生境长期适应而表现出的形态和外貌特征(表1)(Mgdefrau, 1982; Bates, 1998; Glime, 2017)。苔藓植物生活型作为苔藓植物生存策略之一,与环境中的水分和光照条件密切相关,可作为环境条件的重要指示指标(Bates, 1998; Oishi, 2009; Glime, 2017)。垫状型和丛集型通常在具有充足光照、干旱的环境中占优势,树型、交织型、悬垂型和扇型则通常占据荫蔽、湿润的环境,平铺型多见于相对干燥的环境,其下细分的细平铺型偏好荫蔽而粗平铺型多分布于有充足光照的环境(Bates, 1998; Glime, 2017; Lbel et al., 2018)。因此,苔藓植物的生活型与其栖息地的环境条件紧密相关,体现了苔藓物种所承受的选择压力,具有适应性意义(Glime, 2017)。
苔藓植物是附生植物的重要组成部分(Nadkarni, 1984)。附生植物群落的聚集受其自身的扩散能力、地理隔离、植被类型以及宿主特性等的影响(Campos et al., 2006; Klimes et al., 2012)。不同的植被类型具有不同的内部环境条件,直接影响着苔藓植物的种类与分布(汪岱华等, 2012; Gehrig-Downie et al., 2013)。另外,宿主在一棵树上提供不同的基质和各种不同的微生境(microhabitats)(Sanger & Kirkpatrick, 2016),附生植物群落的物种组成和多度随宿主大小、 高度以及树皮特征的变化而变化(Song et al., 2015a; Wang et al., 2016; Patio et al., 2018)。例如,宿主径级越大代表其接受附生植物建群的时间可能更长、能够提供更宽的定殖面积以及具有更高的微生境异质性,树皮粗糙度越高通常意味着具有更强的吸水性和更高的持水力,可能为附生植物提供更丰富的腐殖质和定殖空间(Sáyago et al., 2013; Wagner et al., 2015; Taylor & Burns, 2015)。宿主植物的这些特征直接影响着附生苔藓物种及其生活型组成。
西双版纳是东喜马拉雅和印-缅生物多样性热点和关键地区的一部分,是世界生物多样性保护的关键和热点地区(Myers et al., 2000; Tordoff et al., 2012; Corlett, 2014),分布有我国面积最大的热带森林植被(朱华等, 2015)。热带雨林是西双版纳主要的地带性植被。由于西双版纳的山原地貌,在热带雨林水平地带性植被之上的山地发育有一系列垂向地带性植被,热带山地(低山)常绿阔叶林亦称季风常绿阔叶林,是西双版纳主要的山地植被类型(朱华等, 2015)。然而,刀耕火种、橡胶种植、林下经济等人類活动正大面积破坏该地区的森林植被,导致该地区生物多样性的迅速丧失(朱华等, 2015)。但是,目前针对西双版纳地区不同植被类型苔藓植物多样性的调查研究还很少(Song et al., 2015a; 沈婷等, 2019),对未来开展该地区苔藓植物多样性及其保护工作而言非常不利。因此,本研究通过对西双版纳地区两种典型植被类型附生苔藓植物的物种和生活型组成及分布格局的调查,旨在回答:(1)两种植被的附生苔藓组成和分布格局有何特征;(2)两种植被附生苔藓组成和分布格局在多大程度上受宿主胸径和树皮粗糙度的影响。
1 研究区概况与研究方法
1.1 研究区域概况
西双版纳傣族自治州(99°56′-101°50′ E、21°08′-22°36′ N)位于云南省南部边缘,属于横断山系南部无量山山脉和怒山山脉的余脉,整体地势表现为周围高、中部低,以山地为主,其中分布着许多宽谷盆地、环状低丘和低地,地形错综复杂,地势起伏较大;气候上属于西部型热带季风气候,主要受印度洋季风控制,年均温15.1~21.7 ℃,年降雨量1 193~2 491 mm,降雨时空分配不均,有明显的干、湿季之分,一般2月降雨最少,7月、8月降雨最多,东部降雨量大而西部降雨量较少;冬、春季节多雾,尤其在11月至次年2月期间,雾日超70%,浓雾降水量为0.1~0.3 mm·d-1;森林植被包括32个较为典型的群系,分属于7个主要的植被型,即热带雨林、热带季节性湿润林、热带季雨林、热带山地(低山)常绿阔叶林、热带棕榈林、暖热性针叶林和竹林(朱华等, 2015)。
西双版纳的热带雨林是在热带季风气候下发育的,是在水分、热量和海拔达到极限条件下的热带雨林。热带雨林又进一步划分为两种亚型,即在海拔800 m以下沟谷雨林两侧及低丘台地所分布的热带季节雨林,其与典型的热带雨林相比表现为偏干性且呈现季节性变化,以及紧接季节雨林上缘的热带山地雨林,其以热带种群为主并混杂一定数量的亚热带树种,显示出在生态特征与区系组成上热带季节雨林向南亚热带季风常绿阔叶林过渡的特点。热带山地常绿阔叶林则是指分布于热带非石灰岩山地的常绿阔叶林,其常绿乔木无论在种数还是个体数上均达乔木总数的90%以上,整個森林呈常绿季相(朱华等, 2015)。
1.2 研究方法
1.2.1 样地设置与调查方法 本研究选择西双版纳森林生态系统国家野外科学观测研究站位于勐腊县的样地进行调查,以样地内的热带雨林和热带山地常绿阔叶林两种植被类型作为研究对象,每种类型选择10个20 m × 20 m的样方,每个样方选择10棵样树。样树的选择以胸径20 cm以上的大树优先,数量不足再选取胸径较小的样树。记录样树的胸径和树皮粗糙度信息(表2)。在每株样树1.5 m处的西南面用20 cm × 20 cm的方格网(16格×16格)调查附生于树干上的苔藓,记录苔藓物种、苔藓生活型及所占方格数(Song et al., 2015a)。
样树胸径在树干离地面1.3 m处测量,根据胸径大小,将胸径划分为5个径级,即1=20 cm以下;2=20~30 cm;3=30~40 cm;
4=40~60 cm;5=60 cm及以上。树皮粗糙度参照Male & Roberts(2005)的方法进行目测评估,即将树皮粗糙度划分为9个等级,分别为1.非常平滑;3.平滑但有裂纹;5.有浅裂沟;7.有较深沟壑;9.布满起伏不平的深沟壑;2、4、6、8 为上述等级之间的过渡类型。根据苔藓生活型的划分标准(Glime, 2017),结合野外实际情况,本研究将西双版纳两种植被类型中的附生苔藓植物生活型划分为扇型(fans)、粗平铺型(rough mats)、细平铺型(smooth mats)、悬垂型(pendants)、交织型(wefts)、垫状型(cushions)和丛集型(turfs)7种类型。两种植被类型附生苔藓的物种组成、生活型划分与频度数据如表3所示。
1.2.2 标本鉴定与名录系统 苔鲜植物标本鉴定主要参考《云南植物志》第17卷-19卷(高谦和曹同, 2000; 黎兴江, 2002, 2005)。对未能鉴定的物种,请教有关专家协助鉴定。凭证标本存放于中国科学院西双版纳热带植物园恢复生态研究组实验室。苔藓植物名录采用Goffinet 系统(Goffinet, 2009)对科属一级的分类学单元进行编排。
1.2.3 数据分析 由于苔藓植物的个体数目不易测量,因此附生苔藓植物的多度用盖度代替。某种附生苔藓植物的盖度用其所占方格网面积计算。
数据分析使用R 3.6.1(R Development Core Team, 2019)完成。
1.2.3.1 重要值 由于苔藓植物特殊的性质,因此苔藓植物优势度的度量需要同时考虑盖度和频度指标。附生苔藓植物优势科、属、种的确定采用重要值(IV)公式计算:
IV=F+C2×100。
式中:相对频度F为某种附生苔藓出现的频度/所有附生苔藓出现的频度之和;相对盖度C为某种附生苔藓植物的盖度/所有附生苔藓物种出现的盖度之和。
1.2.3.2 物种累积曲线 通过R 3.6.1中vegan包的specaccum函数作出物种累积曲线,检验样本数量是否足够代表森林中附生苔藓植物实际的物种丰富度。
1.2.3.3 多样性指数 两种森林附生苔藓植物多样性的度量使用α、β和γ多样性指数进行。
α多样性指数,包括每个样方的平均物种丰富度(S)、Marglef指数(d)、Shannon-Weiner多样性指数(H′)和Simpson指数(D):
d=(S-1)/lnN; H′=-∑Si=1PilnPi; D=1-∑Si=1P2i
式中:S为所有附生苔藓植物种数;N为所有附生苔藓物种出现的盖度之和;Pi为第i种附生苔藓植物的相对盖度。
β多样性指数,即Whittaker指数: βw=γα-1。
γ多样性指数,即该森林类型附生苔藓植物的总物种数(Li et al., 2013)。
多样性指数通过R语言psych软件包(Revelle et al., 2018)的alpha函数得到。
1.2.3.4 组间差异检验 检验附生苔藓植物物种和生活型在两种植被的分布格局以及不同径级、不同粗糙度宿主上附生苔藓植物物种丰富度和盖度。在进行分析前对各数据进行Shapiro-Wilk正态性检验和Bartlett方差齐性检验,对于均满足条件的数据使用t检验,对于不满足正态性检验的数据使用非参数的Pairwise Wilcoxon秩和检验和Kruskal-Wallis秩和检验,对于满足正态性但方差不齐的数据使用近似性t检验。
1.2.3.5 直接排序分析 分析两种植被附生苔藓植物物种和生活型的分布格局受宿主径级和树皮粗糙度特性影响的程度。物种数据矩阵每行为一个样方,每列为对应样方的一种苔藓或生活型的盖度,环境因子矩阵每行为一个样方,每列为一个宿主特性因子。使用R语言psych包的corr函数对宿主特性因子进行Spearman相关性分析,对相关的两个因子只保留其中关系最密切的一个。Spearman相关分析结果表明,本研究的两个宿主特性因子之间无显著共线性。在进行排序分析之前,先使用vegan包(Jari et al., 2019)的decorana函数进行除趋势对应分析(detrended correspondence analysis,DCA),判别选择冗余分析(redundancy analysis,RDA)还是典范对应分析(canonical correspondence analysis,CCA)。如果DCA排序 “Axis lengths”前4个轴中最大值小于3,则选择RDA分析;超过4,则选择CCA分析;在3~4之间,则RDA或CCA分析都适用。DCA结果表明,本研究对附生苔藓物种与宿主特性因子进行CCA分析,对附生苔藓生活型与宿主特性因子进行RDA分析。使用vegan包的permutest函数对排序结果进行蒙特卡罗置换检验(Monte Carlo permutation test),检验宿主特性因子对苔藓物种或生活型分布的解释量是否显著。使用vegan包的envfit函数检验每个宿主特性因子的显著性。
2 结果与分析
2.1 两种植被类型附生苔藓植物物种组成
本次调查共记录附生苔藓植物20科39属60种,其中热带雨林附生苔藓植物共19科33属48种,热带山地常绿阔叶林附生苔藓植物共9科14属19种,共有种6种。从物种累积曲线来看(图1),热带雨林附生苔藓植物的物种丰富度还略有上升的趋势,而热带山地常绿阔叶林的物种累积曲线已基本接近渐近线,说明本次调查样本量对于热带山地常绿阔叶林的附生苔藓物种丰富度基本足够,热带雨林则还具有更高的苔藓物种丰富度有待调查,后续研究需要增加调查样树的数量。
根據重要值,排列前3的优势科、属、种如表4所示。优势科方面,3个优势科代表了超过66%的附生苔藓出现频度和盖度,其中对于热带山地常绿阔叶林,优势科覆盖了超过86%的附生苔藓出现频度和盖度。此外,热带雨林附生苔藓优势科所含种数(17种)占其总种数的35.42%,热带山地常绿阔叶林附生苔藓优势科所含种数(10种)占其总种数的52.63%,代表了所在植被类型附生苔藓植物的主要群体。优势属方面,热带雨林附生苔藓的前3个优势属均属于最优势的平藓科,所含种数(5种)占其总种数的10.42%;热带山地常绿阔叶林附生苔藓排列前3的优势属分别对应其优势科的顺序,所含种数(7种)占其总种数的36.84%。优势种方面,两种植被类型的优势种均在其前3优势科和优势属内。从表4可以看出,两种植被类型的附生苔藓优势科、属、种的代表性均较高。
2.2 两种植被类型附生苔藓植物生活型组成
热带雨林附生苔藓植物具有全部7种生活型,最主要的2种生活型为扇型(15种,占其全部种的31.25%)和细平铺型(10种,占其全部种的20.83%);热带山地常绿阔叶林附生苔藓植物只出现5种生活型,没有出现扇型和粗平铺型这两种生活型,最主要的2种生活型为细平铺型(9种,占其全部种的47.37%)和丛集型(4种,占其全部种的21.05%)。每一径级(图2)及每一树皮粗糙度等级(图3),热带雨林附生苔藓生活型都以扇型为最优,热带山地常绿阔叶林则以细平铺型为最优。
2.3 两种植被类型附生苔藓植物物种、生活型、宿主特性的比较
热带雨林的附生苔藓植物多样性指数为1.538,热带山地常绿阔叶林的附生苔藓植物多样性指数为1.357。比较分析结果(表5)表明,两种植被附生苔藓植物的物种丰富度(P=0.017)、Marglef指数(P=0.003)、不同径级(P=0.007)、不同树皮粗糙度(P=0.005)宿主附生苔藓植物的盖度存在显著差异。
2.4 两种植被类型附生苔藓植物物种和生活型分别与宿主特性的排序分析结果
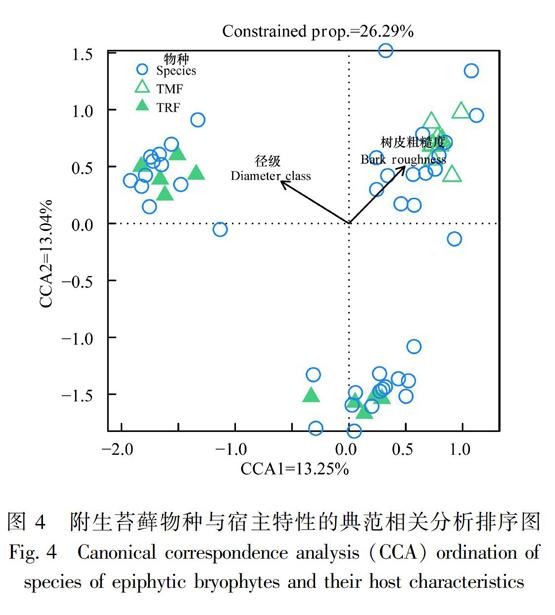
为了解宿主的胸径和树皮粗糙度对苔藓物种及其生活型分布格局的影响,本研究进行了直接排序分析。苔藓物种与宿主特性的CCA排序结果显示(图4),CCA前两轴对物种分布方差变化的总解释量为26.29%(P=0.001),表明宿主径级和树皮粗糙度与苔藓物种的分布是显著相关的,且后者的影响更大。对径级和树皮粗糙度进行偏分析,结果显示径级因子和树皮粗糙度因子各自单独所能解释的变量分别为13.15%(r2=0.950,P=0.001)和13.10%(r2=0.855,P=0.001),表明径级和树皮粗糙度对附生苔藓物种分布的影响大致相等。
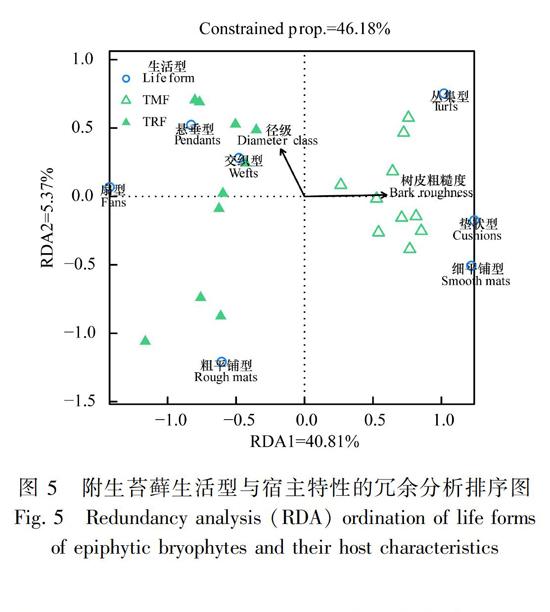
附生苔藓生活型与宿主特性的RDA排序结果显示(图5),RDA前两轴对生活型分布方差变化的总解释量为46.18%(P=0.001),第一轴的解释量(40.81%)为第二轴解释率(5.37%)的五倍,表明附生苔藓生活型的分布主要与第一轴相关。径级和树皮粗糙度的偏分析结果表明,径级对附生苔藓生活型分布的解释量为2.78%(r2=0.612,P=0.001),树皮粗糙度因子单独的解释量为47.65%(r2=0.782,P=0.001)。表明生活型的分布主要受宿主树皮粗糙度的影响。从图5中可看出,热带雨林和热带山地常绿阔叶林的样方点明显分离聚集于各自植被,生活型也明显对植被类型有所偏好而向两边集聚,表现为悬垂型、扇型、交织型、粗平铺型集聚于热带雨林,丛集型、垫状型、细平铺型集聚于热带山地常绿阔叶林。
3 讨论与结论
3.1 两种植被类型附生苔藓植物多样性、分布格局及其驱动因子
不同森林类型中苔藓植物的物种丰富度通常与空气湿度和林冠郁蔽度有密切关系(Gradstein & Sporn, 2010)。热带雨林的林冠较密、林下湿度较高、光照较弱,有利于依靠体表吸收水分和养分的附生苔藓植物生长(Song et al., 2015b)。森林苔藓植物多样性和组成的最主要影响因素是不同微生境(microsites)的可获得性和异质性(Tina et al., 2009)。dor et al.(2013)对人工管理的温带森林中的附生苔藓的物种组成格局和驱动因素进行了研究,表明苔藓植物对微气候(microclimate)(包括光照、温度和空气湿度)的连续性非常敏感。本研究的热带雨林具有更高的β和γ多样性,说明热带雨林附生苔藓植物群落组成变化较明显,反映了热带雨林能提供更多样的生境,孕育更为丰富的附生苔藓植物。
一般认为,宿主胸径、树皮粗糙度越大,附载的附生植物种数和数量越多(Wagner et al., 2015; Zhao et al., 2015; Wang et al., 2016)。本调查的结果表明:宿主径级和树皮粗糙度与苔藓物种的分布是显著相关的,不同径级和不同树皮粗糙度宿主上的苔藓植物盖度差异显著,但是物种丰富度差异不显著。其可能原因之一是先锋种率先在有限的树皮上发展成为优势种群,限制了其他物种的定殖和建立,使得大径级和粗树皮表面的物种丰富度没有明显增加,而先锋优势种群随着时间的累积在树皮表面发展壮大,因此只在盖度方面表现出显著差异,而物种丰富度方面差异不显著。而Gradstein & Culmsee(2010)关于苔藓植物与宿主胸径、树皮粗糙度关系的研究则显示,一些苔藓物种表现出对大胸径和粗树皮有明显偏好,但是物种丰富度与胸径不相关,与树皮粗糙度仅微弱相关。另外,除了本研究关注的宿主径级和粗糙度,宿主的其他特性,包括宿主树皮pH值、持水力和质地等,均与附生植物的附着生长紧密关联(Ma et al., 2009; Song et al., 2011)。和其他植物一样,附生植物群落的组成和多样性格局受到环境因子、生物相互作用和中性过程的综合影响(Gtzenberger et al., 2012; Ovaskainen et al., 2017)。Patio et al.(2018)发现,由于受到扩散的限制(中性过程),小空间尺度的附生植物群落具有很高的相似性,即在相互接近的树木之间,附生植物的物种组成非常相似。生物间的相互作用关系同样可能影响附生植物群落构建和物种共存,但目前我们对附生植物之间的竞争、共存等相互作用关系仍知之甚少(Rydin, 2009),有待将来进一步深入研究。
3.2 兩种植被类型附生苔藓植物的生活型组成及其与宿主特性之间的关系
苔藓植物的生活型是其对环境条件的适应表现,通过分析不同生境下苔藓植物的生活型特征,能够揭示苔藓植物与环境之间的关系(Kürschner, 2004)。本研究中,RDA排序图明显地把附生苔藓植物的生活型分别向两类植被集聚,表现为树型、交织型、悬垂型和扇型更更多分布于热带雨林,而垫状型和丛集型更多分布于热带山地常绿阔叶林。通常树型、交织型、悬垂型和扇型多出现于荫蔽、湿润的环境,而垫状型和丛集型则偏好光照充足、相对干旱的环境(Gradstein & Sporn, 2010; Glime, 2017),这与两类森林林下的微气候环境相一致,较好地验证了生活型作为反映环境条件指标的理论。对于生活型与环境条件尤其是水分和光照条件密切相关的现象,一些学者也做出了部分解释。例如:平铺型群落与基质紧密接触的特点使其能够更好地抵御干旱环境(Bates, 1998);垫状型群落位于边缘的植株由于比居于中央的植株要遭受更多的脱水风险,因此往往比中央部分生长得更少,其半球形剖面可能就是这样形成的(Bates,1998);而丛集型群落有助于降低苔藓侧枝(shoot)和大气的接触面积,减少大气在侧枝空隙之间的流动,有效地减少水分蒸发,从而适应相对干旱的生境(吴鹏程,1998)。从生理角度来看,藓类叶细胞有疣是其适应干旱环境的进化特征,而丛集型苔藓叶细胞有疣的种类比例高达36.2%,因此耐旱性较强(Bates, 1998);扇型群落形成的扁平光合表面则非常适合于在潮湿环境中截留空气中的水分,其较高的细胞壁弹性、耐干旱的渗透调节能力、低光饱和度和低光合补偿点等特点使其能够适应荫蔽、湿润的环境(Song et al., 2015b)。苔藓植物的生活型与其生存策略之间存在着很强的相关性(During, 1979; Bates, 1998; Kürschner et al., 1999)。因此,通过分析不同环境下优势苔藓植物生活型及其分布情况,可更好地掌握苔藓植物所在群落的生境状况(Kürschner et al., 1999; Oishi, 2009; Glime, 2017)。
另外,RDA结果还表明宿主特性特别是树皮粗糙度显著影响苔藓植物生活型的组成和分布。Culberson(1955)认为树皮因子是决定森林附生植物组成和分布的最重要因素。因为附生植物群落主要受湿度的控制,与其他基质相比,树皮是一种干燥的生境,通常含水量很少,因此与树皮含水量直接相关的树皮粗糙度对附生其上的植物起决定作用,是解释附生植物组成和分布的一个重要因素(Barkman, 1958)。生活型作为苔藓植物适应环境的一种功能性状,在不同粗糙度的树皮上也会表现出不同的适应性状。如Gradstein & Sporn(2010)的研究发现,光滑树皮上多见平铺型群落,认为平铺型是苔藓植物在光滑基质上成功建立的一种策略。生活型作为苔藓植物适应其所处环境条件的一种表现,其组成和分布状况可有效反映微生境中的水热条件。考虑到不同森林类型或宿主特性条件下孕育着生活型组成迥异的苔藓植物,因此可考虑将苔藓生活型作为今后森林监测的一项重要指标。
致谢 感谢中国科学院昆明植物研究所马文章老师协助鉴定标本,感谢云南西双版纳森林生态系统国家野外科学观测研究站对野外调查工作的支持,感谢中国科学院-昆士兰州生物技术合作项目组(编号:GJHZ1130)提供部分样地背景资料(表2)。
参考文献:
AZUELO AG, SARIANA LG, PABUALAN MP, 2011.Some medicinal bryophytes: Their ethnobotanical uses and morphology [J]. Asian J Biodivers, 2(1): 49-80.
BARKMAN JJ, 1958. Phytosociology and ecology of cryptogamic epiphytes [M]. Assen: Van Gorcum: 628.
BATES JW, 1998. Is life-forma useful concept in bryophyte ecology? [J]. Oikos, 82(2): 223-237.
BIRSE EM, 1958. Ecological studies on growth-form in bryophytes: III. The relationship between the growth-form of mosses and ground-water supply [J]. J Ecol, 46(1): 9-27.
BOCEDI G, ZURELL D, REINEKING B, et al., 2014. Mechanistic modelling of animal dispersal offers new insights into range expansion dynamics across fragmented landscapes [J]. Ecography, 37(12): 1240-1253.
CAMPOS RI, VASCONCELOS HL, RIBEIRO SP, et al., 2006. Relationship between tree size and insect assemblages associated with Anadenanthera macrocarpa [J]. Ecography, 29(3): 442-450.
CORLETT R, 2014. The ecology of tropical East Asia [M]. 2nd ed. Oxford: Oxford University Press: 336.
CROSBY MR, MAGILL RE, ALLEN B, et al., 2000. A checklist of the mosses [M]. St. Louis: Missouri Botanical Garden: 55-246.
CULBERSON WL, 1995. The corticolous communities of lichens and bryophytes in the upland forests of northern Wisconsin [J]. Ecol Monogr, 25(2): 215-231.
DURING HJ, 1979. Life strategies of bryophytes: A preliminary review [J]. Lindbergia, 5: 2-18.
FREGO KA, 2007. Bryophytes as potential indicators of forest integrity [J]. For Ecol Manag, 242(1): 65-75.
GAO Q, CAO T, 2000. Flora yunnanica tomus (Vol. 17) [M]. Beijing: Science Press: 1-650. [高謙, 曹同, 2000. 云南植物志 (第17卷) [M]. 北京: 科学出版社:1-650.]
GEHRIG-DOWNIE C, OBREGON A, BENDIX J, et al., 2013. Diversity and vertical distribution of epiphytic liverworts in lowland rainforest and lowland cloud forest of French Guiana [J]. J Bryol, 35(4): 243-254.
GLIME JM, 2017. Bryophyte ecology subchapters. 18 : volume 1, chapt. 4-5: Adaptive strategies: growth and life forms [EB/OL]. [2020-2-08]. https://digitalcommons.mtu.edu /bryo-ecol-subchapters/18
GOFFINET B, BUCK WR, SHAW AJ, 2009. Morphology, anatomy, and classification of the Bryophyta [J]. Bryophyte Biol, 2: 55-138.
GRADSTEIN R, CULMSEE H, 2010. Bryophyte diversity on tree trunks inmontane forests of Central Sulawesi, Indonesia [J]. Trop Bryol, 31(2): 95-105.
GRADSTEIN SR, SPORN SG, 2010. Land-use change and epiphytic bryophyte diversity in theTropics [J]. Nova Hedwigia, 138: 311-323.
GTZENBERGER L, DE BELLO F, BRTHEN KA, et al., 2012. Ecological assembly rules in plant communities—approaches, patterns and prospects [J]. Biol Rev, 87(1):111-127.
HE X, HE KS, HYVNEN J, 2016. Will bryophytes survive in a warming world? [J]. Perspect Plant Ecol Evol Syst, 19: 49-60.
JAESCHKE A, BITTNER T, REINEKING B, et al., 2013. Can they keep up with climate change?-Integrating specific dispersal abilities of protected Odonata in species distribution modeling [J]. Insect Conserv Divers, 6(1):93-103.
JARI O, GUILLAUME BF, MICHAEL F, et al., 2019. vegan: community ecology package. R package version 2.5-6. https://CRAN.R-project.org/package=vegan
KIMMERER RW, 2005. Patterns of dispersal and establishment of bryophytes colonizing natural and experimental treefall mounds in northern hardwood forests [J]. The Bryologist, 108(3): 391-402.
KLIMES P, IDIGEL C, RIMANDAI M, et al., 2012. Why are there more arboreal ant species in primary than in secondary tropical forests? [J]. J Anim Ecol, 81(5): 1103-1112.
OISHI Y, 2009. A survey method for evaluating drought-sensitive bryophytes in fragmented forests: A bryophyte life-form based approach [J]. Biol Conserv, 142(12): 2854-2861.
OVASKAINEN O, TIKHONOV G, NORBERG A, et al., 2017. How to make more out of community data? A conceptual framework and its implementation as models and software [J]. Ecol Lett, 20(5): 561-576.
PATIO J, GMEZ-RODRGUEZ C, PUPO-CORREIA A, et al., 2018. Trees as habitat islands: Temporal variation in alpha and beta diversity in epiphytic laurel forest bryophyte communities [J]. J Biogeogr, 45(8): 1727-1738.
REVELLE W, 2018. psych: Procedures for personality and psychological research, Northwestern University, Evanston, Illinois, USA, https://CRAN.R-project.org/package=psych Version = 1.8.12.
RYDIN H, 2009. Population and community ecology of bryophytes [M]//GOFFINET B. Bryophyte biology. 2nd ed. Cambridge: Cambridge University Press: 393-444.
SANGER JC, KIRKPATRICK JB, 2016. Moss and vascular epiphyte distributions over host tree and elevation gradients in Australian subtropical rainforest [J]. Aust J Bot, 63(8): 696-704.
SYAGO R, OPEZARAIZA-MIKEL M, QUESADA M, et al., 2013. Evaluating factors that predict the structure of acommensalistic epiphyte-phorophyte network [J]. Proc Roy Soc B-Biol Sci, 280(1756): 20122821.
SHEN T, SONG L, GUO XL, et al., 2019. Habitat preferences of epiphytic bryophytes along the vertical gradient and their indicator functions in the tropical dipterocarp rainforest [J]. Guihaia, 39(6): 776-787. [沈婷, 宋亮, 郭新磊, 等, 2019. 龍脑香热带雨林附生苔藓沿宿主垂直梯度的微生境偏好及其指示作用 [J]. 广西植物, 39(6): 776-787.]
SHI XM, SONG L, LIU WY, et al., 2017. Epiphytic bryophytes as bio-indicators of atmospheric nitrogen deposition in a subtropical montane cloud forest: Response patterns, mechanism, and critical load [J]. Environ Pollut, 229:932-941.
SONG L, LIU WY, MA WZ, et al., 2011. Bole epiphytic bryophytes on Lithocarpus xylocarpus (Kurz) Markgr. in the Ailao Mountains, SW China [J]. Ecol Res, 26(2): 351-363.
SONG L, LIU WY, NADKARMI NM, 2012. Response of non-vascular epiphytes to simulated climate change in a montane moist evergreen broad-leaved forest in Southwest China [J]. Biol Conserv, 152: 127-135.
SONG L, MA WZ, YAO YL, et al., 2015a. Bole bryophyte diversity and distribution patterns along three altitudinal gradients in Yunnan, China [J]. J Veg Sci, 26(3):576-587.
SONG L, ZHANG YJ, CHEN X, et al., 2015b. Water relations and gas exchange of fan bryophytes and their adaptations to microhabitats in an Asian subtropicalmontane cloud forest [J]. J Plant Res, 128(4): 573-584.
SPORN SG, BOS MM, KESSLER M, et al., 2010. Vertical distribution of epiphytic bryophytes in an Indonesian rainforest [J]. Biodivers Conserv, 19(3): 745-760.
STEINHORST L, KUDLA J, 2017. Plant science: sexual attraction channelled in moss [J]. Nature, 549(7670): 35.
TAYLOR A, BURNS K, 2015. Epiphyte community development throughout tree ontogeny: An island ontogeny framework [J]. J Veg Sci, 26(5): 902-910.
TINA F, MRIALIGETI S, KIRLY I, et al., 2009. The effect of light conditions on herbs, bryophytes and seedlings of temperate mixed forests in rség, Western Hungary [J]. Plant Ecol, 204(1): 69-81.
TORDOFF AW, BALTZER MC, FELLOWES JR, 2012. Key biodiversity areas in the Indo-Burma hotspot: Process, progress and future directions [J]. J Threatened Taxa,4(8): 2779-2787.
WAGNER K, MENDIETA-LEIVA G, ZOTZ G, 2015. Host specificity in vascular epiphytes: A review of methodology, empirical evidence and potential mechanisms [J]. AoB Plants, 7: plu092.
WANG DH, WANG YF, ZUO Q, et al., 2012. Bryophyte species diversity in seven typical forests of the West Tianmu Mountain in Zhejiang, China [J]. Chin J Plant Ecol, 36(6): 550-559. [汪岱华, 王幼芳, 左勤, 等, 2012. 浙江西天目山主要森林类型的苔藓多样性比较 [J]. 植物生态学报, 36(6): 550-559.]
WANG X, LONG W, SCHAMP BS, et al., 2016. Vascular epiphyte diversity differs with host crown zone and diameter, but not orientation in a tropical cloud forest [J]. PLoS ONE,11(7): e0158548.
WILLIS KJ, MACDONALD GM, 2011. Long-term ecological records and their relevance to climate change predictions for a warmer world [J]. Ann Rev Ecol Evol Syst, 42:267-287.
WU PC, 1998. Bryological biology, introduction and diverse branches [M]. Beijing: Science Press: 1-357. [吳鹏程, 1998. 苔藓植物生物学 [M]. 北京: 科学出版社:1-357.]
ZHAO M, GEEKIYANAGE N, XU J, et al., 2015. Structure of the epiphyte community in a tropical montane forest in SW China [J]. PLoS ONE, 10(4): e0122210.
ZHU H, WANG H, LI BG, et al., 2015. Studies on the forest vegetation of Xishuangbanna [J]. Plant Sci J, 33(5): 641-726. [朱华, 王洪, 李保贵, 等, 2015. 西双版纳森林植被研究 [J]. 植物科学学报, 33(5):641-726.]
(责任编辑 周翠鸣)
