籽粒高效特异性表达启动子的克隆与表达分析
2021-06-09王美华高洁李玉莲张淑娟宋国琦张荣志李玮李吉虎李根英
王美华,高洁,李玉莲,张淑娟,宋国琦,张荣志,李玮,李吉虎,李根英
(山东省农业科学院作物研究所/农业部黄淮北部小麦生物学与遗传育种重点实验室/小麦玉米国家工程实验室,山东 济南 250100)
启动子是一段位于基因上游能够与RNA聚合酶结合的DNA序列,也是RNA聚合酶的识别和结合序列,CAAT-box和TATA-box是启动子的典型特征。启动子上游的顺式作用元件是转录因子结合位点,也是决定转录激活和抑制关键因子,因此启动子是调控基因表达的重要元件[1]。启动子分为组成型启动子、组织或器官特异性启动子和诱导型启动子[2]。组成型启动子调控下的基因,在不同组织器官中没有明显差异。植物中最常使用的组成型启动子是花椰菜花叶病毒(CaMV)35S启动子[3]、玉米的泛素蛋白(ubiquitin)启动子[4]、水稻的肌动蛋白(actin)启动子等[5]。组成型启动子驱动的外源基因在整株中表达,而有些器官人们并不想让它表达外源蛋白,因为异源蛋白质或代谢产物的积累容易打破植物原有的生理生化平衡,影响植株正常生长,有些产物甚至有毒,造成植物死亡。另外,同一种启动子用于多个外源基因的驱动,可能引起基因沉默或共抑制现象[6]。因此,寻找多种类型的组织特异性启动子,减少组成型启动子的使用是科学家一直努力的方向。
组织特异性启动子驱动的基因一般在根、茎、叶、果实和种子等某一个特异性的器官组织中特异表达。随着基因工程的发展,人们从禾谷类作物中克隆了许多种子特异性启动子,并将它们应用于禾谷类籽粒品质的改良之中。应用较多的是来源于普通小麦的高分子量麦谷蛋白亚基1DX5、Bx17和水稻的G1t等[7-9]。但这些启动子的启动外源基因表达活性存在差异,不能满足基因工程的需要,而且在需要多基因串联时,如果利用同一启动子驱动不同的基因表达,很容易导致基因沉默。本研究拟寻找转录效率更高的种子特异性启动子,用于提高目标基因的表达,改良禾谷类植物的加工和营养品质。
1 材料与方法
1.1 试验材料
质粒pGEM-Teasy和pCAMBIA3301购自长沙赢润生物技术有限公司;质粒和转基因小麦基因组DNA的提取试剂盒购自天根生化科技(北京)有限公司;配制培养基所需无机盐、维生素、抗生素以及激素、GUS染色所需药品购自Sigma-Aldrich中国公司;农杆菌菌株购自北京华越洋生物科技有限公司;PCR扩增所需的Pfu、rTaq酶购自宝生物工程(大连)有限公司。
用于克隆胚乳特异性启动子(glo启动子)的燕麦品种为四倍体大燕麦,由张家口农业科学院杨才研究员提供;用于基因转化的小麦品种为Fielder。
1.2 引物设计与PCR扩增
采用DNA提取试剂盒从黑暗中萌发的燕麦幼苗中提取基因组DNA,用作扩增启动子的模版,扩增引物根据NCBI数据库中的参考序列AY795082进行设计,上游引物5′-CGAAGCTTTGGAAAGTCATTTTGCCTC-3′,下游引物5′-GCCCATGGTAGATTGTAGAAGGTGGATTGG-3′(划线处分别为Hin dⅢ和NcoⅠ的酶切位点)。PCR反应体系(20μL):基因组DNA(50 ng/μL)1μL,上、下游引物(10 pmol/L)各2 μL,dNTPs(250μmol/L)2μL,10×buffer(50 mmol/L KCl,10 mmol/L Tris-HCl,1.5 mmol/L MgCl2,pH 8.3)2μL,Pfu高保真酶1μL,水10 μL。PCR扩增程序:94℃3 min;94℃45 s,60℃45 s,72℃1 min,36个循环;72℃10 min。PCR产物在1.2%琼脂糖凝胶中分离,EB染色后进行紫外观察并拍照保存。
1.3 基因转化与转基因植株的PCR检测
转化普通小麦参考Zhang等[10]的方法,PCR检测所用引物为:上游引物5′-GGTCACTCATTACGGCAAAGT-3′,下游引物5′-GACGACCAAAGCCAGTAAAGT-3′。PCR反应体系(20 μL):基因组DNA(50 ng/μL)1μL,上、下游引物(10 pmol/L)各2μL,dNTPs(250μmol/L)2μL,10×buffer(50 mmol/L KCl,10 mmol/L Tris-HCl,1.5 mmol/L MgCl2,pH 8.3)2μL,rTaq高保真酶1μL,水10μL。PCR扩增程序:94℃3 min;94℃30 s,58℃30 s,72℃45 s,36个循环;72℃8 min。PCR产物在1.2%琼脂糖凝胶中电泳分离,EB染色后进行紫外观察并拍照保存。
1.4 不同部位GUS基因表达活性分析
GUS组织染色液的配制:0.2 mol/L磷酸钠缓冲液100 mL;0.1 mol/L亚铁氰化钾和0.1 mol/L铁氰化钾各1 mL;0.5 mol/L Na2-EDTA 8 mL;已溶于400μL DMSO中的200 mg X-gluc;水90 mL。
GUS组织染色:待转基因植株开花后两周,挑选PCR鉴定为阳性的转基因苗,取其部分籽粒、叶和根分别装入5 mL离心管中,用GUS染色液完全浸没,37℃振荡过夜,次日用70%乙醇冲洗3次,观察GUS基因表达强度。
2 结果与分析
2.1 胚乳特异性启动子的克隆
胚乳特异性启动子大小约960 bp(图1),将其与pGEM-Teasy载体进行连接,含有该启动子的重组载体命名为pTglo。

图1 glo启动子的PCR扩增
将获得的启动子序列与已报道序列(AY795082)进行比对,相似性达99%。两个序列之间存在9个SNPs,其中在85 bp处A变为G,364 bp处T变为A,369 bp处A变为T,620 bp处发生A碱基的插入,640 bp处T变为C,701 bp和788 bp处G变为A,858 bp处T变为C,961 bp处C变为A(图2)。对启动子组成元件进行分析表明,该启动子含有胚乳特异表达所必须的5个Skn-1(GTCAT)基序,其中3个位于正链上,分别在第7、235、873 bp处,2个位于负链上,分别在第176 bp和551 bp处(绿色覆盖并加边框的区域)。在启动子的正链第622 bp和830 bp处存在胚乳表达相关的顺式调控元件GCN4(加框部分)。在启动子的正链550 bp和负链869 bp处有两个O2位点,曾在玉米中被验证为醇溶蛋白的代谢顺式调控元件(红色斜体序列)。在启动子的869 bp处有一个回文结构ACATGTCATCATGT,该序列是胚乳特异表达所必需的。这些结构元件与启动子的活性是否存在联系,还需要进一步验证。除了上述与胚乳特异性表达有关的元件,该启动序列中还有许多与逆境响应有关的序列元件,包括负链573 bp处有一个ABA顺式响应元件ABRE(红色字母部分);正链825 bp处有一个赤霉素顺式响应元件(蓝色字母部分);正链561 bp处有一个TC-rich重复序列(灰色覆盖部分),该序列在烟草中是逆境和防御反应的顺式作用元件;负链第725 bp处有一个水杨酸反应顺式调控元件TCA-element(绿色覆盖部分);正链的694 bp和负链的697 bp处各有一个TGACG-motif,该序列在大麦中与茉莉酸的顺式调控有关。这些与抗逆有关的结构单元的存在或许与燕麦的高度抗逆性有关。
2.2 表达载体构建
为了验证新克隆的启动子与已报道的glo启动子启动后续基因表达强度的差异,委托生工生物工程(上海)股份有限公司对NCBI数据库中提供的glo启动子序列(AY795082)进行人工合成,并直接克隆到质粒pCAMBIA3301的Hin dⅢ和NcoⅠ酶切位点之间,完成的重组质粒命名为 pGloUdiA1。
将从大燕麦中新克隆到的glo启动子序列利用Hin dⅢ和NcoⅠ从pTglo上切下,电泳分离并做凝胶回收后与经过同样酶切的pCAMBIA3301大片段进行连接,PCR鉴定阳性克隆子(图3)。
挑选部分阳性克隆子进行酶切鉴定(图4),并进行测序验证,完成的重组质粒命名为pGloUdiA2,基因表达框见图5。
选用载有GUS基因但没有启动子的植物表达载体pCAMBIA1381xb作为阴性对照。利用GUS基因的特异性引物进行PCR扩增,扩增产物长度为616 bp(图6)。PCR结果显示,经过筛选后最终获得转pGloUdiA1、pGloUdiA2和pCAMBIA1381xb的阳性小麦植株分别为8、12、5个。

图3 PCR鉴定阳性克隆子

图4 阳性克隆子的酶切鉴定
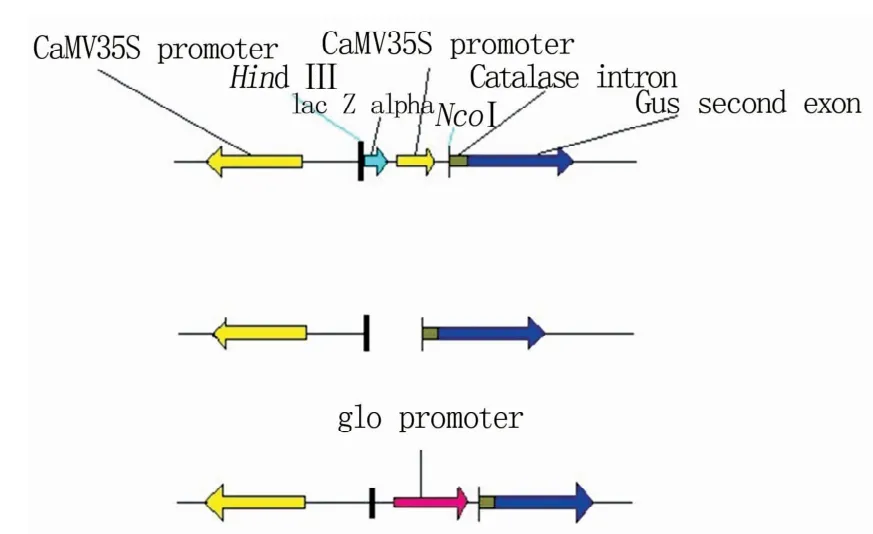
图5 pGloUdiA2的表达框结构
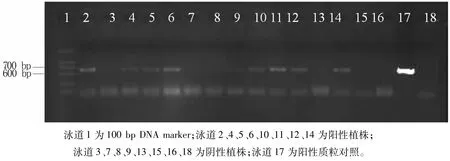
图6 转基因植株的PCR鉴定
2.3 基因特异性表达检测
GUS染色结果显示,无论是转化质粒pGloUdiA1还是pGloUdiA2,其根和叶中都无蓝色信号出现(图7),与转化pCAMBIA1381xb的阴性对照表现一致。但在转化pGloUdiA1和pGloUdiA2的小麦籽粒中都有蓝色信号出现(图8),说明该启动子具有良好的组织特异性。转化pGloUdiA2的籽粒中蓝色信号强度大约是转化pGloUdiA1的5倍,表明新克隆的glo启动子序列的基因表达活性更强。转化阴性对照质粒pCAMBIA1381xb的籽粒中没有观察到蓝色信号,说明GUS基因表达差异是由启动子的表达所造成的。新克隆glo启动子序列的高效启动活性和高度的组织特异性表达特性为利用转基因技术改良小麦籽粒性状提供了良好的基础材料。

图7 转基因植株根和叶的GUS染色

图8 转基因小麦植株的籽粒GUS染色
3 讨论与结论
启动子是调控基因表达的重要元件,本研究利用已经报道的燕麦启动子序列作为参考序列,从我国四倍体燕麦中克隆了燕麦球蛋白基因新的启动子序列,经过比对发现新克隆的启动子序列长960 bp,与参考序列AY795082存在9个SNPs。对该启动子组成元件进行分析表明,该启动子含有胚乳特异表达所必须的5个Skn-1(GTCAT)基序,其中3个位于正链上,2个位于负链上;在启动子的正链第622 bp和830 bp处存在胚乳表达相关的顺式调控元件GCN4;在启动子的正链550 bp和负链869 bp处有两个O2位点,曾在玉米中被验证为醇溶蛋白代谢顺式调控元件;在启动子的869 bp处有一个回文结构ACATGTCATCATGT,该序列是胚乳特异表达所必需的。利用上述启动子驱动GUS基因表达,结果表明该启动子启动功能基因表达的强度约为参考序列的5倍,为利用转基因技术改良小麦籽粒性状提供了良好的基因表达驱动元件。
