午潮山常绿次生阔叶林主要木本植物功能群及其演替特征
2021-06-04吴初平袁位高朱锦茹黄玉洁焦洁洁臧润国
金 超,吴初平,丁 易,袁位高,朱锦茹,黄玉洁,焦洁洁,臧润国,江 波,*
1 浙江农林大学 林业与生物技术学院, 杭州 311300 2 浙江省林业科学技术研究院, 杭州 310023 3 中国林业科学研究院森林生态环境与保护研究所,国家林业和草原局森林生态环境重点实验室, 北京 100091 4 南京林业大学南方现代林业协同创新中心, 南京 210037
次生林是目前大多数森林所处的演替状态[1],了解其群落组成和物种关系对于保护森林多样性和碳汇具有重要意义[2- 3]。研究群落次生演替过程中的物种组成和生态系统功能可以提供促进群落正向演替的信息[4]。其中,分析不同生活史策略的物种相互关系及其与生态系统功能的联系可以帮助理解和预测群落演替过程[5]。探索次生林不同演替阶段的物种间关系为管理和修复森林资源提供极大的便利。
评价并划分群落中具有相似功能的物种可以帮助量化物种对环境的响应程度和理解群落构建的过程[6]。根据物种演替地位和最大潜在高度将其划分功能群是被广泛接受的方法[7],而数量分类通过计算植物功能性状相似性划分功能群,为本研究提供了新视角[8]。植物功能性状能反映其生态策略[9],其中,叶经济谱[10]和木材经济谱[11]能较好地描述物种性能的功能性状。比如,相较于檫木(Sassafrastzumu)、麻栎(Quercusacutissima)等喜光的先锋树种,青冈(Cyclobalanopsisglauca)等耐阴树种在弱光照下的寿命更长且生长速度缓慢,其功能性状表现为低比叶面积和高木材密度[12- 14]。根据实验目的选取植物功能性状将物种划分到对应的功能群,为研究复杂的生态系统过程建立基础[15- 16]。有研究利用功能群划分来研究物种组成复杂的热带山地雨林[17],此外,还有分析不同生境条件下植物功能群和物种性状的响应,揭示了土壤异质性的重要性[18]。
功能群内和功能群之间的生态位和物种联结可以反映物种对生境资源的竞争关系,进而加强对群落结构、功能及演替的理解。生态位指物种和环境之间相互影响和相互作用的关系[19],它表达了群落中物种对资源的竞争程度,竞争者间的生态位差异代表良好的共存关系[20],而种间联结反映物种的空间分布和功能依赖关系[21]。其概念广泛应用于研究群落组成及指导植被的恢复与重建[22- 24],但由于缺乏植物功能的机制基础,直接比较群落物种的生态位和种间关系仅能得出片面的结论。Fry等指出通过植物性状划分功能群比简单的草本、灌木和乔木划分更能体现生物多样性和生态系统功能的关系[25]。因此,利用功能群探索物种关系能为研究森林生态系统提供重要的实际应用价值。
午潮山位于杭州西郊,曾经过度的森林砍伐和不合理的土地利用导致大部分原始植被被破坏,经过长期的改造和恢复,形成以木荷(Schimasuperba)、青冈、苦槠(Castanopsissclerophylla)为建群种的次生林[26]。由于受到早期种植的针叶树种影响,午潮山森林的林分结构和演替进度在一定程度上被影响。鉴于此,本研究选取午潮山常绿次生阔叶林70个样地中的主要物种进行功能群划分,并评估不同功能群间的物种关系。旨在回答以下问题:1)午潮山森林群落功能群内和功能群间的植物生态位和种间关系如何?其处于什么演替阶段?2)如何根据功能群和种间关系的相关结论对午潮山常绿次生阔叶林进行生态系统修复以及促进群落演替?
1 研究方法
1.1 研究区概况及样地调查
本研究在浙江午潮山国家森林公园林区内进行(33°41′ N, 120°00′ E),午潮山林区为典型的亚热带常绿次生阔叶林。午潮山平均海拔264 m,最高494.7 m,森林覆盖率达93%,土壤类型为红壤。地处亚热带气候类型,年平均温度为16.1℃,最冷一月的月平均温度为3.6℃。全年的降雨量为1400.7 mm,年无霜期为240 d,年平均相对湿度为76%,年平均日照时数为1800—2100 h。午潮山原始天然林植被破坏后人为种植大量经济树种:湿地松(Pinuselliottii)和杉木(Cunninghamialanceolata),对森林进行保护和恢复后逐渐演替到目前的常绿次生阔叶林。目前,研究区森林群落的乔木层以木荷、青冈、杉木和麻栎为优势树种,灌木层的主要树种包括毛柄连蕊茶(Camelliafraterna)、薄叶山矾(Symplocosanomala)、马银花(Rhododendronovatum)、窄基红褐柃(Euryarubiginosavar.attenuata)和檵木(Loropetalumchinense)。
分别于2018年4月和2019年10月参照热带森林科学研究中心(Center For Tropical Forest Science, CTFS)的原则[27]在午潮山林区的北坡和南坡建立不重叠的森林样地。在整个景观中用全站仪设置了70个0.04 hm2(20 m×20 m)的样地,每个样地继续划分成16个5 m×5 m的小样方。对样方内1.3 m位置处胸径(DBH)≥1 cm的存活木本树种进行标定并挂牌,记录树种名称、胸径、树高和空间坐标,同时记录每个样方的海拔、经纬度、坡度、坡向等生境特征。
1.2 重要值和生态位特征
重要值可以通过参数量化物种在群落中地位和作用[28]。生态位宽度能衡量物种利用资源的范围,可使用Levins指数(BL)和Shannon指数(BS)来表示,而Pianka生态位重叠指数(Oik)表示生态位重叠程度,其数值范围从0(无重叠)到1(完全共享资源)。
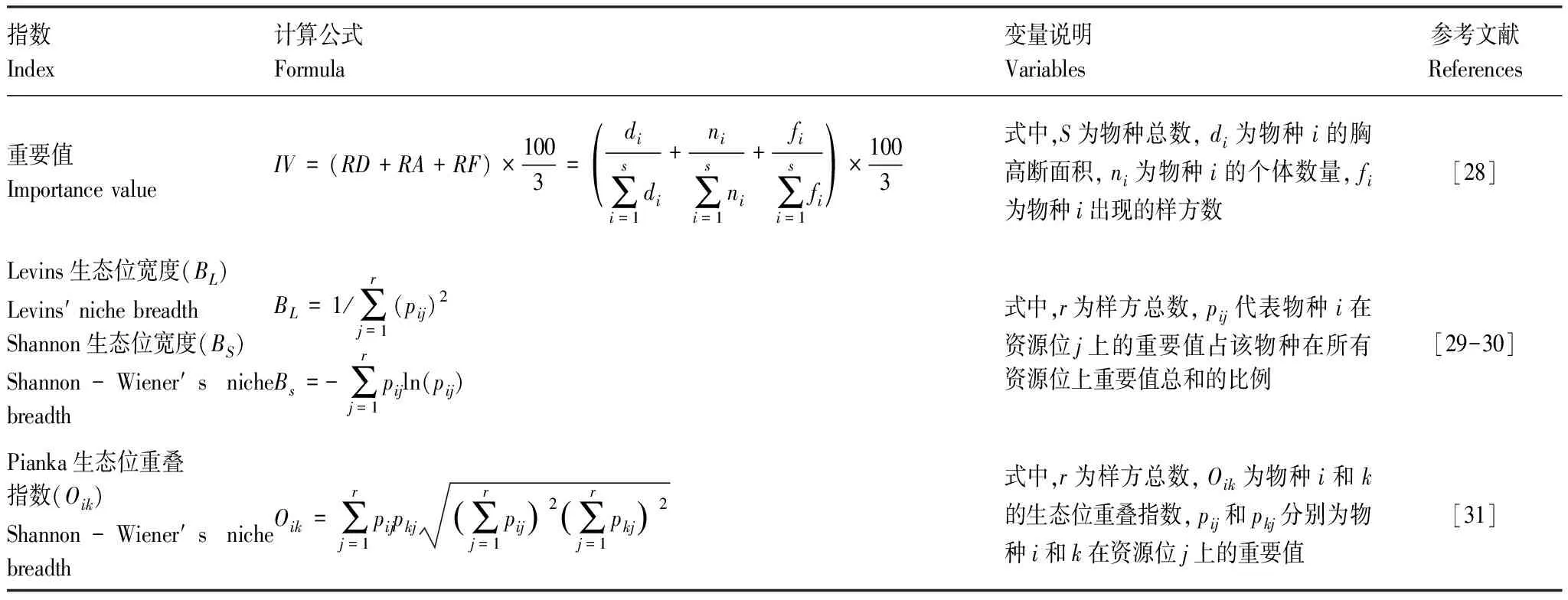
表1 重要值和生态位指数计算公式
1.3 功能群划分性状的选择
计算70个样地中物种的重要值,将排序前45(重要值占全部物种95%)的木本植物作为划分功能群的主要物种。演替早期和后期的物种在习性和生态策略有明显的区别,拥有较高资源获取率(如喜阳、生长速度快、种子小且多、木材密度低等)的物种在资源丰富的环境占据主导地位,但物种会倾向于发展对极端环境更有忍耐能力和能保存资源的性状[5]。本文使用与植物生理和形态相关的功能性状划分功能群[7, 9- 11],分为分类性状和连续性状。分类性状包括生长型、喜光性和寿命;连续性状包括叶片氮含量(LNC, mg/g)、叶片磷含量(LPC, mg/g)、叶片钾含量(LKC, mg/g)、比叶面积(SLA, cm2/g)、木材密度(WD, g/cm3)、叶片干物质含量(LDMC, g/g)、叶片叶绿素含量(ChL, SPAD)、最大潜在树高(Hmax, m)。生长型通过物种的形态和生长特性划分为乔木、小乔木或灌木和灌木;物种喜光性则基于亚热带常绿阔叶林植被演替阶段不同物种的分布特征从喜光到耐阴分为5个等级;物种寿命根据物种早期演替特征利用二分法分为长寿命和短寿命。这些分类变量均反映了物种对环境因子的要求和响应,以及物种在生态系统中的作用和地位。分类变量的数据主要来源于中国植物志网络版(http://www.iplant.cn/frps),部分缺失的则通过样地勘测和咨询植物学教授。
物种的连续性状数据主要通过实验获取。按照国际植物性状选取标准,选取胸径10 cm以上的目标物种10株(个体数少的除外)[32],在每株个体的中冠位置(高度≤5 m)采取5片完整、健康的成熟叶片,同时取5—10根枝条(1 cm≤DBH≤2 cm)。使用电子天平、叶面积仪(LI-COR 3100C)和叶绿素仪(SPAD 502 Plus Meter, Konica Minolta, Japan)分别测量叶片鲜重、叶面积和叶绿素含量。最后,将叶片置于65℃的烘箱内烘干至恒重后进行叶片干重测定,随后对叶片进行叶片氮、磷、钾含量的测定[33]。用排水法测定去皮后的植物枝条体积,然后将其置于103℃的烘箱内烘干,恒重后称重。枝条密度利用枝条干重与体积之比计算得出,为了减少样地内树种的破坏,使用物种的枝条密度代替木材密度。每个物种的最大潜在树高信息参考中国植物志网络版(http://www.iplant.cn/frps)。
1.4 总体联结性检验
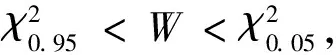

表2 物种联结性指数计算公式
1.5 种间联结性分析
1.5.1χ2检验
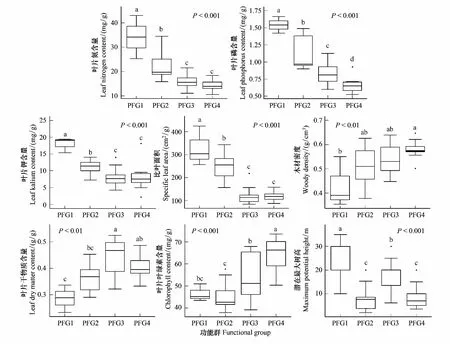
χ2统计量通过使用一个基于2×2列联表来对种间联结性进行定性分析[23],由于本实验为非连续取样,因此采用Yates的连续校正公式来矫正χ2统计量。若ad-bc>0,种间联结为正联结,反之为负联结。当χ2<3.841,表明物种间是独立且联结不显著(P>0.05);若3.841≤χ2<6.635,说明物种间联结显著(0.01 1.5.2Dice值(DI) χ2检验仅提供了种间联结的定性结果,但是不能量化物种对的联结程度[19],而DI值可以确定其联结程度。DI的范围是(0,1),若DI=0,则无联结,表明物种间完全独立且几乎不会同时出现。相反,DI越接近1,物种对就越有可能同时出现。 1.5.3Spearman秩相关系数(rs) χ2检验和DI值使用二元数据判断物种联结性,因此不可避免的会忽略一些要素,如,物种多度[35]。Spearman秩相关检验可以对物种间的线形关系进行定量分析,且包含物种联结和物种协方差的信息[36],分析种对同时出现的可能性。其值域为[-1,1],正值为正相关,负值为负相关。 本研究中,数据分析在R3.6.0[38]中完成。功能群的划分使用基于11个功能性状的聚类分析,采用FD包[39]中的dbFD函数,其优势是可处理不同类型的功能性状。由于存在部分数据缺失,分析过程采用Gower相异性矩阵[40- 41],分析方式采用k-均值聚类(k-means partitioning),聚类的组数采用方差比准则(Calinski-Harabasz criterion, VRC)[42],通过比较组间和组内平方和误差的比值判断得出,比值越大,聚类效果越好。不同功能群的功能性状差异用单因素方差分析检验,显著差异使用Tukey检验(Tukey Honestly Significant Difference tests,P=0.05)。生态位宽度、生态位重叠程度使用R包“spaa”[43]计算,可视化在“corrplot”包[44]中完成。 本次调查的70个样地中,共有木本植物13596株119种,分属40个科76个属。午潮山林区重要值排序前45的木本植物根据性状矩阵通过k-均值聚类分析,并结合方差比准则聚类为4个功能群。PFG1由6个物种组成,主要由麻栎和湿地松等早期喜阳先锋树种组成;PFG2由11个物种组成,主要是赛山梅(Styraxconfusus)、小叶石楠(Photiniaparvifolia)和白檀(Symplocospaniculata)等中期中性树种;PFG3包含15个物种,主要为木荷、青冈和杉木等中后期树种组成;PFG4则由13种后期耐阴树种组成,主要为毛柄连蕊茶、薄叶山矾和马银花等(表3)。4个功能群之间的连续功能性状差异显著(图1),从物种组成和性状来看,PFG1到PFG4代表了由演替初期过渡到演替后期的物种。具体来说,物种性状由喜光、短寿命、低叶片营养物质和低木材密度过渡到耐阴、长寿命、高叶片营养物质和高木材密度。 表3 功能群划分 图1 午潮山4个功能群性状差异(平均值±标准差)Fig.1 The difference of plant functional traits in four functional groups in Wuchao Mountain (mean±SD)P值代表组间差异显著性水平,不同的字母表示差异显著(P<0.05);PFG1、PFG2、PFG3和PFG4代表功能群 本实验按相对重要值比重(占各自功能群重要值80%—90%)取4个功能群主要物种来研究种间关系(表4)。在4个功能群中,重要值最大的分别是麻栎(3.25)、赛山梅(1.29)、木荷(19.35)、毛柄连蕊茶(7.79)。 表4 午潮山4个功能群主要物种的重要值和生态位宽度 使用Levins(BL)和shannon(BS)指数测量生态位宽度,对环境适应性强的物种具有较大的生态位宽度。午潮山4个功能群主要物种的生态位宽度存在较大差异,Levins和Shannon指数测定结果差异不大,BL和BS的变化范围分别为9.45—62.32和2.27—4.18。其中檵木的生态位宽度最大,其BL和BS分别为62.32和4.18,随后是老鼠矢(Symplocosstellaris)(59.80和4.15)和木荷(58.52和4.14),湿地松最小(9.45和2.27)。重要值和生态位宽度成正比,但是也有例外,如PFG1中的湿地松重要值虽然大,但是其生态位宽度却很小,而白栎(Quercusfabri)重要值只有1.70但是有较高的生态位宽度(39.32和3.76)。不同功能群的物种数值反差更加明显。 午潮山森林群落4个功能群主要物种的Pianka生态位重叠指数(Oik)介于0—1之间,0.6≥Oik>0的有106对,占总对数的55.8%,Oik>0.6的有81对,占总数42.6%(图2)。Oik=0的仅有3对,麻栎和檫木、麻栎和赛山梅、湿地松和赛山梅。高生态位重叠值的物种对,通常有相对高的生态位宽度和重要值。PFG1物种与其他物种之间的生态位重叠程度普遍较低。高生态位重叠指数主要集中于除PFG1功能群之外的物种之间,其中,杉木和白花龙与其他树种生态位重叠指数大多低于0.6。随着演替进行,同一功能群物种间的生态位重叠值增加。 图2 午潮山4个功能群主要物种生态位重叠指数矩阵图Fig.2 Correlation matrix of the Niche overlap of the dominant species in four functional groups in Wuchao Mountain χ2检验可以准确的反映种间联结的显著度,且提供了量化显著度的参数。由于木荷出现在所有样方中以及算法的缺陷,χ2统计量为无限大,因此木荷的χ2检验结果不能考虑在内。除去木荷,共171个种对(图3),80对为正联结,占46.8%;84对负联结,占49.1%;7对没有联结。正负联结比为0.95,正相关为显著和极显著的分别为4和10,占2.3%和5.8%;负相关为显著和极显著的分别为3和14,占1.8%和8.2%。结果与总体联结性呈负联结一致,联结程度较弱。 图3 午潮山3个功能群主要物种χ2指数矩阵图Fig.3 Correlation matrix of the χ2 text results of the dominant species in three functional groups in Wuchao Mountain3.841≤χ2<6.635:物种间显著正联结(0.01 DI指数被用于定量表达种间联结性。190对物种中,112对(58.9%)表现为极强联结,24对(12.6%)表现为强联结(图4)。此外,42对为弱联结,12对无联结,分别占总物种对的22.1%和6.3%。种间联结的结果与生态位重叠基本一致,强联结的种对和高重叠的生态位代表了相似的环境需求。 图4 午潮山3个功能群主要物种DI指数矩阵图Fig.4 Correlation matrix of the Dice index (DI) results of the dominant species in three functional groups in Wuchao MountainDI<0.2:无联结;0.2≤DI<0.4:弱联结;0.4≤DI<0.6:强联结;0.6≤DI:极强联结 Spearman秩相关系数可以用来分析物种协方差(图5)。56对物种(29.5%)表现为极显著正相关,10对(5.3%)表现为显著正相关。此外,55对物种(28.9%)表现为极显著负相关,9对(4.7%)表现为显著负相关,60对不相关。此结果补充了木荷与其他物种的联结性,且正负相关种对的比值为1.03,与χ2检验结果基本一致,但是种间显极著联结的对数明显多于χ2检验,这是由于χ2检验不能准确表达物种间联结差异性[35]。与生态位重叠结果类似,演替后期物种间的联结强度要大于演替早期物种。 图5 午潮山3个功能群主要物种Spearman秩相关系数矩阵图Fig.5 Correlation matrix of the Spearman′s rank correlation coefficient of the dominant species in three functional groups in Wuchao Mountainrs≥0.1492:极显著正相关(P≤0.01);0.1492>rs≥0.1133:显著正相关(0.01 本文应用k-均值聚类法根据物种的叶经济谱和木材密度等11个功能性状将其客观地划分为4类功能群,并分析不同功能群间的性状差异定义其演替阶段。随着次生演替的进行,植物从大比叶面积、叶片氮磷钾含量高的资源获取性策略向比叶面积小、叶片氮磷钾含量匮乏的保守型策略转化;关于叶经济谱的研究结果同样证明了这一点[45]。同时,本实验观察到采取积极生存策略的演替早期树种具有低木材密度、喜阳、短寿命的特征,与之前的研究一致[17]。4个功能群有效地体现了对不同生态环境响应和生态系统功能贡献的植物组合,这为深入研究植物种间关系和群落演替趋势提供框架基础[15, 46- 47]。 群落是物种连续更迭的结果,这些物种被环境筛选,同时又构成了新的环境[48]。演替初期,人工种植的杉木和湿地松以及自然更新的麻栎等先锋种改变了植被毁坏的午潮山地区的环境条件。随着演替进行,木荷[49]、青冈、毛柄连蕊茶和薄叶山矾等中性或耐阴树种逐渐增多,而喜阳的针叶树种更新缓慢甚至死亡[50]。经过不同种群的长期竞争和适应,最终形成了午潮山以木荷为优势种的森林群落。 物种对环境资源的利用程度可以通过物种生态位和种间关系的变化格局体现。麻栎和湿地松等树种(PFG1)有较高的重要值,主要由于其均为大径材,对周围其他树种幼苗有较大影响。但是,这些喜阳的先锋树种拥有较窄的生态位宽度且缺乏幼苗,意味着它们适应和获取当前生境资源的能力要弱于其他物种,生长更新缓慢。PFG2、PFG3中大多是耐阴适阳的乔木树种,如小叶石楠、木荷和青冈等,它们生态位宽度较大,说明在群落中分布较广,对环境资源有较强的利用能力[24]。PFG4中的耐阴灌木拥有大生态位宽度,侧面反映了当前生境是阔叶林密闭树冠下的湿冷生境。生态位重叠程度越低表示物种对相似资源的需求越低,反之,则代表种间竞争激烈[51]。从资源竞争和利用的角度来看,耐阴树种均拥有大生态位宽度和生态位重叠,表明它们具有较强地适应当前生境的能力以及需求相似生境资源的生存策略。Gu等研究了不同演替动态次生林的生态位和种间联机,发现随着演替进行,耐阴树种与其他树种的生态位重叠值随之增加[19]。综上,随着不同种群的长期竞争和适应,午潮山森林群落正向后期演替阶段进行。 群落结构和物种组成的稳定性会随着群落演替不断上升,越接近顶级群落越稳定[52- 53]。本实验中,主要物种的整体联结性和正负联结比均显示种间水平以负联结为主或与正联结持平,但是显著正联结种对占优势。负联结种对较多可能是由于湿地松作为样地中的外来种,胸径均超过20 cm,生长速度快且对资源占有能力较强,与大部分树种都成显著负联结。有研究指出,正负联结比小于1的情况除了次生演替还有可能是人为连续干扰造成的[54]。午潮山在1960年作为科研基地后,人为砍伐和偷盗树木的行为被禁止,而1992年后,其作为国家森林公园开放,次生演替明显受到周边人为活动的影响,而这些因素是如何影响森林群落结构还需要进一步的调查和研究。 不同功能群物种的种间联结强度不同,而极显著的种间联结物种对达到了半数以上,要高于一些亚热带常绿阔叶林的结果[55]。正联结的物种对意味着分享相似资源同时拥有较宽的生态位重叠,如石栎(Lithocarpusglaber)和毛柄连蕊茶,这与之前的研究一致[19, 24, 56]。负联结的种对说明物种有不同的环境需求,例如,青冈在林下形成的环境可供毛柄连蕊茶等耐阴物种生长,但是不适合檫木幼苗这类喜光的物种;早期喜阳先锋种(PFG1)与其他功能群物种均处于弱强度负联结,在群落中生态位宽度和生态位重叠均比较低。以上提到的物种间负联结的原因属于生境差异的范畴,而类似PFG3和PFG4之间的种间关系则是由于资源竞争导致的负联结[22, 56],如木荷和毛柄连蕊茶幼苗对林下的资源竞争激烈。因此,在演替中后期,资源竞争是物种竞争的主要原因。 尽管人工种植针叶树种能加快群落早期演替,但也限制演替中后期(PFG3和PFG4)的树种更新。人类活动同样影响森林演替和群落组成结构,如森林片段化和边缘效应会影响种子的传播机制[57]。在此,本文针对午潮山森林群落的演替和结构稳定性,结合实验结果为森林管理提供一些意见。 首先,自然演替需要相当漫长的时间,选择性的砍伐先锋种和人为补植演替中后期的物种是加速林分演替不错的方式。考虑到一些有经济和景观价值的先锋种(如檫木),在林冠层间隙或者与耐阴树种(如窄基红褐柃)成正联结搭配种植,既充分利用了有限的环境资源又维护了生物多样性和群落稳定性。此外,白栎、马银花和杉木等价值低的树种,有较大的生态宽度并与青冈、木荷等分享相似的资源,适当伐除可释放供中后期树种的生长资源。从而推动中后期目的树种更新演替,加速午潮山的森林群落向顶级群落发展。其次,湿地松作为外来物种虽然在造林初期生长快、成活率高[58],但是在森林演替中后期与其他物种成负联结,减缓群落演替进程。根据文本结论,可在湿地松周围搭配种植白花龙和木荷等喜光的植物,减少湿地松造成的开放地。 最后,本研究在关于生态系统功能和群落结构的基础上明确了功能群对理解物种利用资源的作用及其对森林经营的实际意义,为分析和指导次生林更新恢复提供了实用的框架,而生态位和种间联结为生境和资源利用提供详细的指示。未来的研究还需改善这种方法,如选用更加有代表性的性状或者能更全面展示物种相互关系的方式,以期有效地提升森林结构和经营模式。1.6 数据分析
2 结果
2.1 功能群的划分及性状差异
2.2 重要值和生态位宽度
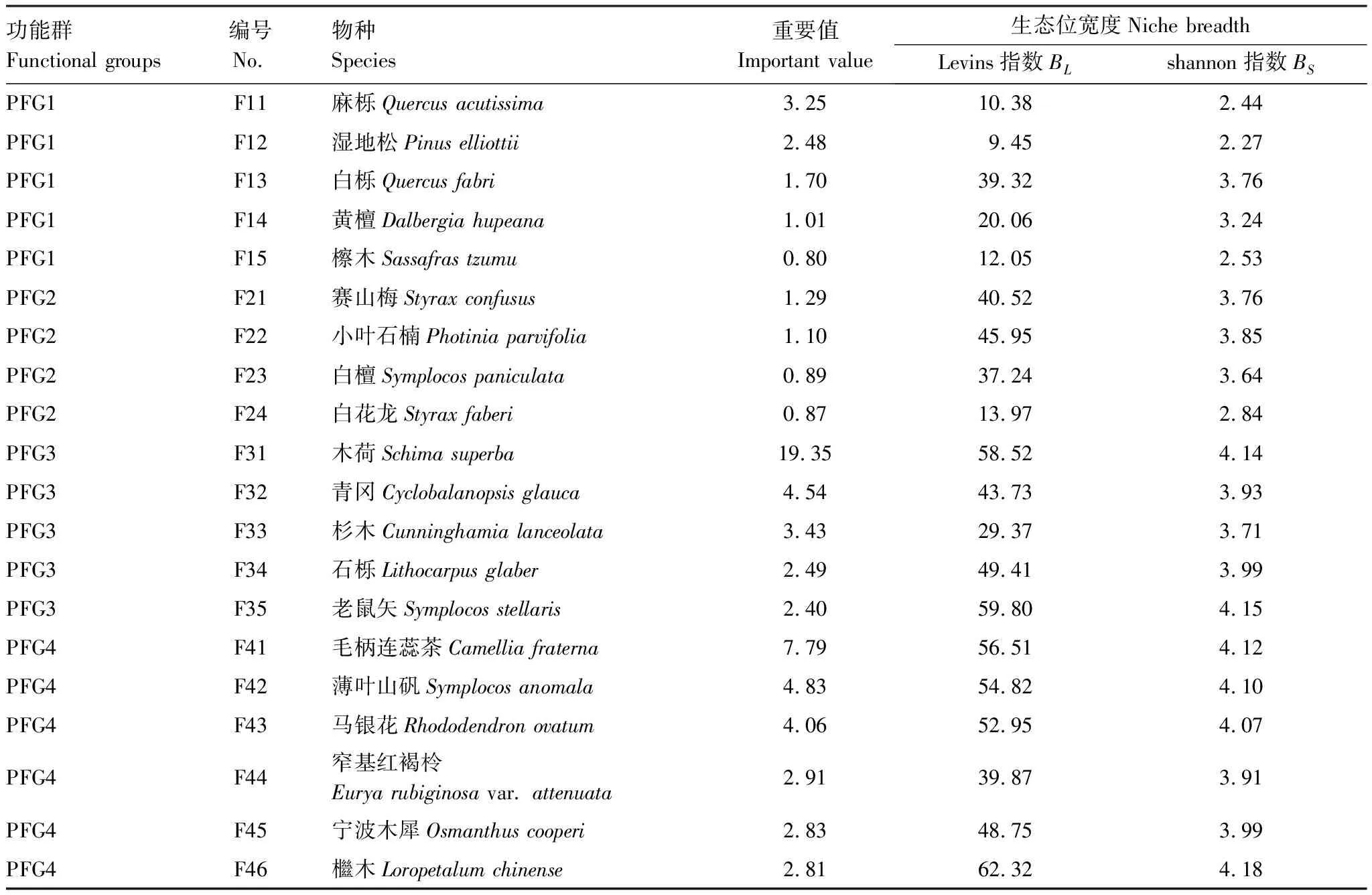
2.3 生态位重叠

2.4 物种的总体联结性
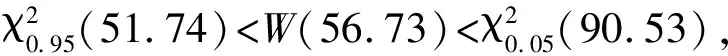
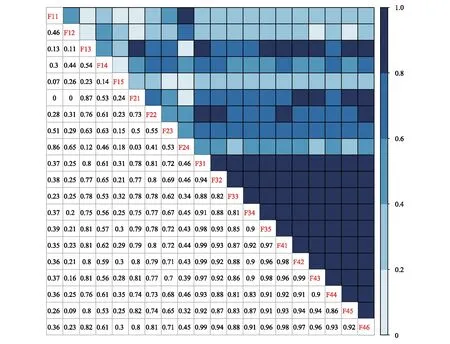
2.5 种对间的联结性

3 讨论
3.1 午潮山植物功能群划分
3.2 午潮山次生林的演替趋势
3.3 午潮山优势物种的种间竞争
3.4 森林管理建议
