欧洲山杨bHLH转录因子家族全基因组分析
2021-06-03沈方圆王岚春
沈方圆, 王岚春, 李 校
(四川大学生命科学学院 生物资源与生态环境教育部重点实验室, 成都 610064)
1 引 言
bHLH (basic helix-loop-helix)是存在于真核生中最广泛的一类转录因子家族,也是植物中最大转录因子家族之一,可以通过与靶基因中特定的基序结合来调控基因的表达.bHLH转录因子包含保守的bHLH结构域,由一个位于N′端的碱性区域(basic region)和一个位于C’端的α螺旋1-环-α螺旋2 (helix-loop-helix)结构组合而成.碱性区域是bHLH转录因子与靶基因的识别区域,含有大量的碱性氨基酸,能与靶基因中保守的六核苷酸E-box (5′-CANNTG-3′)结合,且多数可识别G-box(5′-CACGTG-3′).HLH区域为一个长度可变的环连接两个由疏水氨基酸组成的亲水α螺旋构成.通常,依赖HLH中疏水氨基酸的相互作用,形成同二聚体或异二聚体的二聚结构域,促进蛋白质间的互作.目前,bHLH转录因子家族全基因组水平的系统分析已在多种模式植物中进行.2003年,Toeldo-Oritz等人[1]在拟南芥中鉴定出162个成员,被分为21个亚家族.2006年,Li等人[2]从水稻中鉴定到了167个成员,分为22个亚家族.番木瓜、玉米和土豆中分别被鉴定出73、208和159个bHLH家族成员,且被划分为15~25个亚家族不等[3-5].
在植物中,bHLH是仅次于MYB类的第二大转录因子家族,其家族成员广泛参与植物的新陈代谢、活性成分合成、金属离子内稳态以及信号激素调节等重要生物学过程,还在植物的生长发育和环境应激中扮演着重要角色[6].SPATULA是植物中第一个已知的在花器官发生中起作用的bHLH基因,基因突变的研究表明,它促进心皮边缘和由心皮产生的花粉道组织的生长[7].2011年,Ju-Seok等人[8]在水稻中发现水稻bHLH转录因子家族成员OsbHLH148,构成水稻中OsbHLH148-OsJAZ-OsCOI1信号模块,作用于茉莉酸调控的抗旱性基因表达的初始响应.Ji等人[9]从红毛柳中克隆出一个bHLH基因——ThbHLH1,该基因特异性结合G-box motif,可以正向调控P5CS、BADH、CAM、POD、SOD、LEA、HSP等一系列提升非生物胁迫耐受性的基因.这些基因的高表达导致脯氨酸和甜菜碱水平升高,提高作为第二应激信使的Ca2+水平,增强过氧化物酶(POD)和超氧化物歧化酶(SOD)活性,从而减少活性氧(ROS)的积累,增强植物的非生物抗逆性.SlPRE2作为番茄中果实发育的调节剂,通过赤霉素途径影响植物对赤霉素的响应[10].ZmPTF1在玉米中是磷酸饥饿诱导bHLH转录因子,参与玉米根系发育和是ABA合成、信号通路和耐旱性的正向调节因子[11].
欧洲山杨(Populustremula),杨柳科杨属植物,耐寒、喜光,广泛分布于北半球,在中国分布于新疆阿尔泰等地区.欧洲山杨同时具有观赏性和实用性的树种,可作为造张和人造纤维的原材料.同时,作为云杉林的主要先锋树种和伴生树种之一,拥有绿化荒山保持水土的作用,此外也因其秋叶变红,常作为风景林区、城乡绿化的候选树种.2018年,欧洲山杨全基因组组装完成,使得从全基因组水平上研究欧洲山杨抗逆基因家族和进行后续转录分析成为可能,为科研工作者增加杨柳科植物的研究材料.本研究使用一系列生物信息学方法对欧洲山杨基因组中bHLH转录因子家族成员进行系统地鉴定,并对保守残基、染色体分布、基因结构和系统发育关系进行分析,最后通过转录组分析筛选出参与干旱胁迫应激的bHLH转录因子.这些结果和信息为进一步研究欧洲山杨bHLH基因的进化和生物学功能提供良好的基础.
2 材料与方法
2.1 材 料
从PopGenIE[12]数据库(http://popgenie.org/)下载欧洲山杨(P.tremula)全基因组序列、蛋白质序列及注释文件;拟南芥bHLH转录因子家族的蛋白质序列从拟南芥tair[13]数据库(https://www.arabidopsis.org/)中获取.
2.2 方 法
2.2.1 欧洲山杨bHLH转录因子家族的鉴定 为了鉴定欧洲山杨bHLH家族,从Pfam(http://pfam.xfam.org/)[14]中获取植物bHLH转录因子家族的HMM模型(PF00010),使用HMMER 3.3[15]软件对欧洲山杨的蛋白质数据库进行本地搜索,然后以拟南芥bHLH蛋白质序列作为目标序列用blastp对欧洲山杨的蛋白质数据库进行比对,将两种方法得到的结果去冗余.最后使用SMART[16]在线软件(http://smart.embl-heidelberg.de/)和CDD[17]数据库(https://www.ncbi.nlm.nih.gov/cdd)验证预测的蛋白序列中bHLH结构域的完整性.
2.2.2 多序列比对及系统发育分析 使用ClustalW[18]软件对167条欧洲山杨bHLH蛋白的bHLH保守结构域进行多序列比对,参数为默认设定.系统进化树在MEGA5[19]软件中使用邻接法构建,其Bootstrap值为1000.
2.2.3 motif分析及基因结构的鉴定 将预测的欧洲山杨bHLH家族蛋白序列提交至MEME[20](http://meme-suite.org/)检索序列中的保守基序,motif的预期搜索个数设置为10.使用GSDS2.0(http://gsds.cbi.pku.edu.cn)[21]对bHLH基因序列的外显子/内含子结构进行解析.
2.2.4 染色体定位和基因复制分析 利用欧洲山杨基因组的注释信息,将各个bHLH转录因子家族成员的基因定位到染色体中.再使用MCScanX[22]软件扫描bHLH基因间的共线性以检测基因复制事件.
2.2.5 欧洲山杨bHLH基因的表达分析 从ENA数据库(https://www.ebi.ac.uk/ena)[23]下载欧洲山杨受干旱胁迫根部样本(SRA号:ERR380521)和正常生长的野生型根部样本(SRA号:ERR380515)转录组数据,运用HISAT2+Stringtie[24-25]流程计算各基因表达量,再使用edgeR[26]对两组数据进行差异表达分析,选择logFC绝对值大于1且FDR小于0.001的基因为差异表达基因.
3 结果分析
3.1 欧洲山杨bHLH转录因子家族鉴定
为了全基因组鉴定欧洲山杨中bHLH转录因子家族,我们首先使用方法中描述的两种方法,再根据Carretero-Paulet等人[27]计算的标准最终从欧洲山杨的蛋白质序列中,鉴定出167个含有保守bHLH结构域(PF00010)的PtbHLH蛋白,且欧洲山杨bHLH转录因子蛋白的数量与其他植物,如拟南芥(162个)、水稻(170个)、芝麻(130个)中bHLH转录因子数量基本吻合[2,28].根据Toeldo-Oritz等人[1]对拟南芥bHLH基因家族进行命名的规则,将欧洲山杨bHLH转录因子命名为PtbHLH1到PtbHLH167.
3.2 欧洲山杨bHLH保守残基鉴定
通过使用clustalx软件对167个欧洲山杨bHLH结构域的蛋白质序列进行多序列比对,并根据保守度大于50%的残基生成一致性序列.欧洲山杨bHLH结构域包含35保守的氨基酸残基,其中10个存在于咸性区域部分,9个在第一个α螺旋,1个在loop中,还有15个在第二个α螺旋中(图1).两个螺旋间的环状区域在氨基酸数量和排列顺序上显示出显著的差异.PtbHLH蛋白家族中大多数成员的环状区域一般由6~16个氨基酸组成,但有三个成员有较长的环形区域分别是PtbHLH17(80个)、PtbHLH73(63个)和PtbHLH16(39个).

图1 欧洲山杨bHLH转录因子家族的保守结构域Fig.1 The conserve domain of P.tremula bHLH proteins
3.3 bHLH转录因子亚家族分类及进化分析
为了分析PtbHLH的进化关系,利用拟南芥和欧洲山杨中保守的bHLH蛋白质结构域使用邻接法构建系统发育树(图2).系统发育树表明,拟南芥和欧洲山杨共被分为了22个亚家族,其中Ⅶ、ⅩⅥ、ⅩⅦ、ⅩⅧ、ⅩⅩⅩ、ⅩⅪ和ⅩⅫ这7个亚家族中只有拟南芥bHLH,这可能是由于个别PtbHLH在进化过程中的丢失所造成.在22个亚家族中,Subfamily Ⅱ是最大的一个亚家族同时也是PtbHLH数量最多的家族,分别拥有11个AtbHLH和28个PtbHLH.Ⅶ和Ⅺ为最小的亚家族,只有4个成员,其中Ⅶ只含有拟南芥bHLH,Ⅺ只含有欧洲山杨bHLH.所有的PtbHLH转录因子家族成员被分到15个亚家族中.早先的研究结果表明,植物中bHLH转录因子家族一般可分为15~25个亚家族,因此我们的结果与此一致.
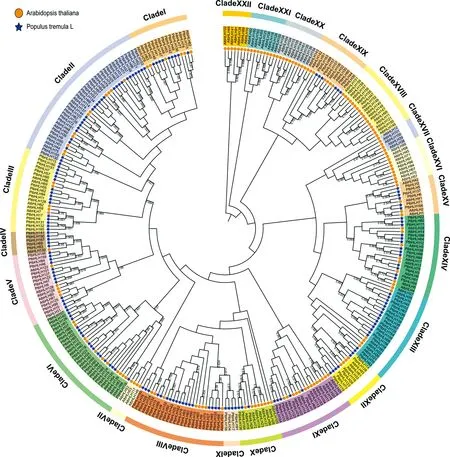
图2 欧洲山杨和拟南芥bHLH转录因子家族的系统发育树Fig.2 Phylogenic analysis of bHLHs from P.tremula and A.rabidopsis
Pires和Dolan[29]的研究结果表明,被子植物中大多数bHLH蛋白的多样性早在4.43亿年前早期陆地植物中就建立起来,且在进化过程中一直高度保守.为探究欧洲山杨bHLH基因家族的进化地位,基于六种植物的bHLH保守区进行系统发良分析,这六种植物分别为双子叶植物——拟南芥(Arabidopsisthaliana)、单子叶植物——水稻(Oryzasativa)、杨属——毛果杨(Populustrichocarpa)、苔藓植物——小立碗藓(Physcomitrellapatens)、绿藻——莱茵衣藻(Chlamydomonasreinhardtii)和欧洲山杨(图3).欧洲山杨167个bHLHs分布在除2、9和14亚家族外的11个亚家族中,同样也可能存在成员在进化过程中丢失的情况.根据亚家族分类,6个PtbHLH与绿藻植物bHLH在2号亚家族中聚为一类,151个与苔藓植物bHLH在8个亚家族(3-4、6-8、10-11和13)中聚在一起.因此推测,这6个和151个PtbHLH可能分别由藻类和苔藓的祖先进化而来.

图3 欧洲山杨等6种植物的系统进化关系Fig.3 Phylogenic relationships of six plants including P.tremula
3.4 bHLH蛋白的GO注释和功能分析
为了理解欧洲山杨bHLH的功能,我们对PtbHLH进行了GO注释,结果如图4所示.大多数预测的PtbHLH蛋白定位在细胞核内,且具有蛋白二聚体活性和DNA结合活性.涉及植物生长发育过程的PtbHLH蛋白最多,约占58%,其次是光刺激反应和代谢过程占40%.此外,PtbHLH蛋白还参与到胁迫反应、细胞分化和生物合成等过程.目前为止,大约40%的拟南芥bHLH基因家族的功能已被鉴定.系统发育分析表明,有39个PtbHLH与已知功能的AtbHLH有紧密的关系,因此它们可能与其拟南芥同源蛋白有相似的功能(表1).
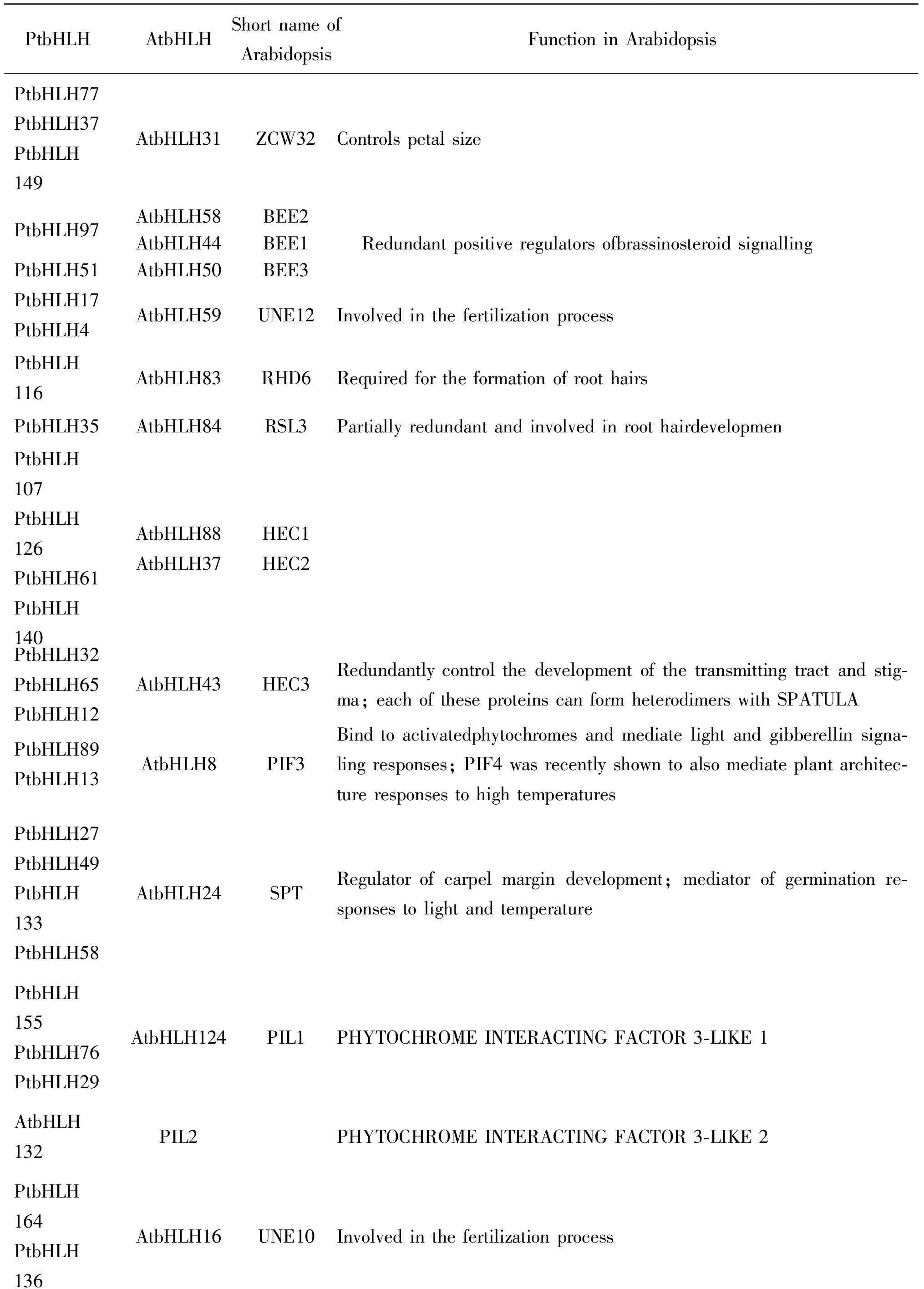
表1 欧洲山杨bHLH转录因子家族成员功能
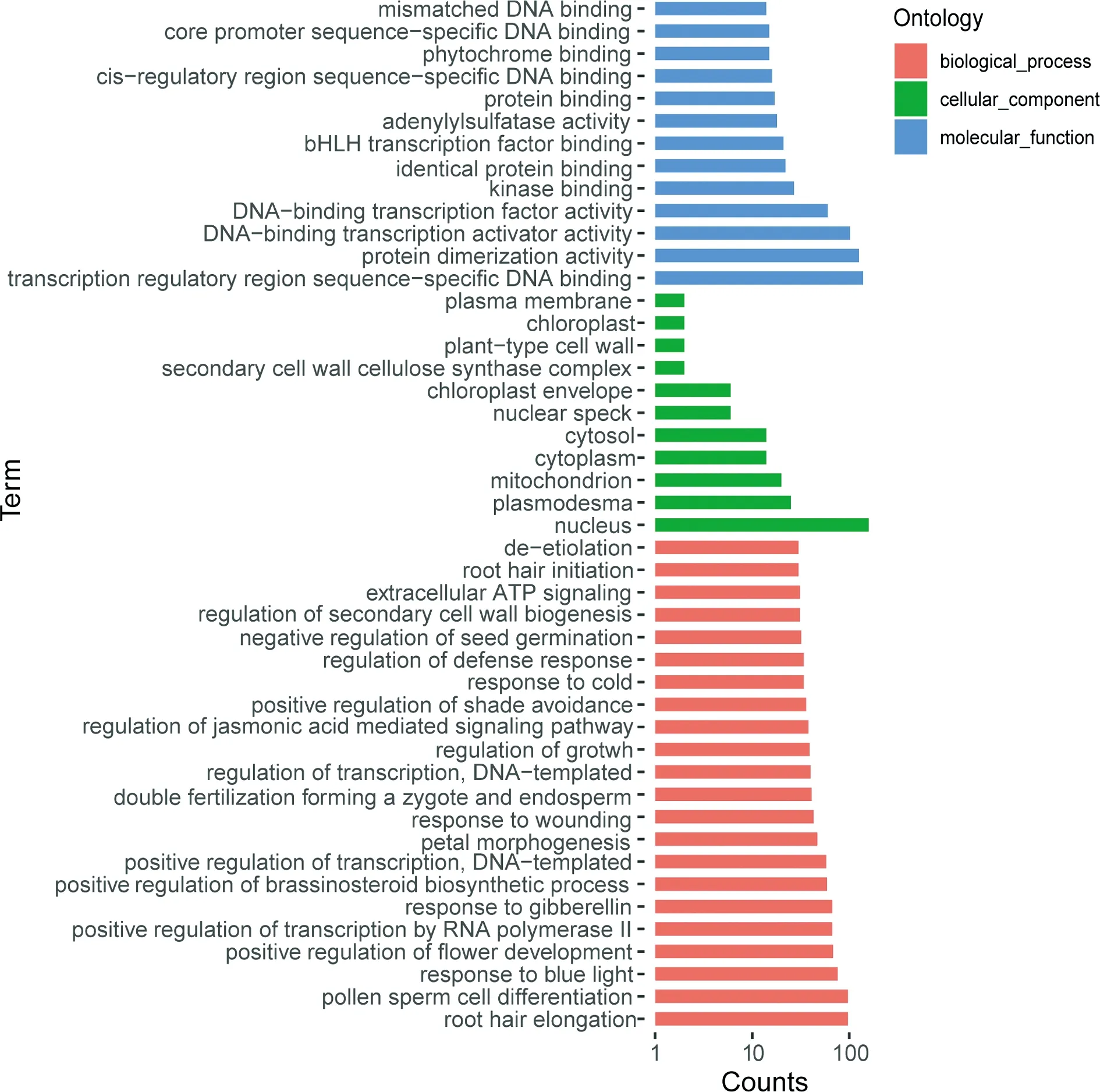
图4 PtbHLH蛋白的GO注释Fig.4 Gene ontology annotation of ptbHLH proteins
3.5 bHLH转录因子亚家族基因结构及motif分析
为了得到更多的欧洲山杨bHLH转录因子家族的信息,我们分别使用GSDS和MEME对PtbHLH的基因结构和保守基序进行预测.结果表明:7个PtbHLH基因只有外显子,110个PtbHLH基因有3-7个外显子,10个PtbHLH基因有9个及以上外显子(图5).同一亚家族内的PtbHLH基因在内含子和外显子的数量及长度上有很大的相似性,而在不同的支系中又存在着显著的差异.例如每个属于亚家族ⅩⅨ的成员,除了PtbHLH157,他们的基因都包含2个外显子;分支Ⅸ中成员的外显子和内含子数目为7~8个.PtbHLH家族蛋白的motif分析确定了10个假定的保守蛋白基序(图5).大多数PtbHLH蛋白序列(不包括PtbHLH112和PtbHLH64)都拥有两个高度保守的基序:motif1和motif2,分别用红色和天蓝色方块表示.Motif1含有32个氨基酸,由碱性区域、α螺旋1和环状结构组成;Motif2有29个氨基酸,代表bHLH结构中的第二个α螺旋.在每个亚家族中,大多数蛋白质保守基序的组成是相似的.例如,亚家族Ⅰ的所有成员都含有motif1、2、3、5和7;亚家族Ⅰ的所有16个成员都鉴定出motif1、2和4.基因结构和motif分析结果都进一步支持了转录因子家族的进化分类关系.
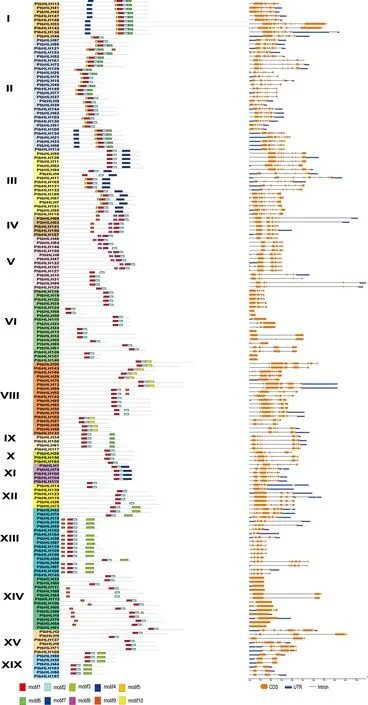
图5 欧洲山杨bHLH家族的基因结构和motif分布Fig.5 Conserved motifs and gene structures analyses of PtbHLHs
3.6 bHLH转录因子亚家族染色体定位及基因复制分析
根据注释信息将每个PtbHLH基因定位到欧洲山杨染色体上,结果如图4所示.所有167个bHLH转录因子家族成员无规律、不均匀的分布在欧洲山杨19条染色体上,其中2号染色体上有23个,是拥有PtbHLH数量最多的染色体;其次是5号和6号染色体,有19个基因数量最少的是17号染色体,只含有1个PtbHLH.之前的研究表明,串联重复是植物基因家族扩展的主要原因之一,我们在欧洲山杨中发现了2个串联复制事件分别是位于3号染色体的PtbHLH60/PtbHLHL111和位于18号染色体的PtbHLH38/PtbHLHL108.形成串联复制的基因事件的基因来自同一个亚家族,且拥有相似的基因结构.
3.7 欧洲山杨根部bHLH基因对干旱胁迫的应答分析
在植物中bHLH转录因子主要通过调节相关基因的表达从而对非生物胁迫进行适应性应答.为了解欧洲山杨根部组织中bHLH在应对干旱胁迫时的作用,对正常生长和干旱条件下生长的欧洲山杨根部bHLH基因进行差异表达分析.依据logFC>1或<-1和FDR<0.001的条件筛选出了4个差异表达的PtbHLH基因,其中PtbHLH102和PtbHLH122表达量显著上升,PtbHLH138和PtbHLH10显著下调.PtbHLH122是拟南芥AtbHLH130的直系同源基因,AtbHLH130能促进钾离子的吸收从而调节根木质部的渗透,保证营养从根部运输到芽,从而维持质膜的稳定,防止植物体内水分持续流失[30].

表2 干旱和正常植株中表达差异显著的PtbHLH基因
4 讨 论
bHLH转录因子家族在植物种是仅次于MYB类的第二大转录因子家族,对植物生长发育过程中存在广泛的影响,在抗逆相关的调控网络和信号通路中起关键作用.但目前对bHLH转录因子家族的研究大多集中在拟南芥、土豆和水稻等模式植物中,杨柳科植物中涉及较少.
本研究利用生物信息学方法从欧洲山杨全基因组中鉴定出167个bHLH转录因子.碱性区域(basic region)和α螺旋1-环-α螺旋2(helix-loop-helix)组成的bHLH保守结构域在PtbHLH中由62个氨基酸组成,其中有32个保守度大于50%的氨基酸.N端碱性区域高度保守的Glu-15和Arg-58是决定DNA绑定E-box(CANNTG)区域的关键氨基酸残基;两个螺旋中保守性大于95%的Leu-69、Pro-81和Leu-176都与之前在其他植物bHLH结构域的研究一致.随后,对提取的欧洲山杨和拟南芥的bHLH蛋白质结构域进行系统发育分析,并根据拟南芥bHLH家族的亚家族分类和进化树的拓扑结构,将欧洲山杨bHLH家族分为15个亚家族.基因结构和Motif分析的结果显示,同一亚家族中的成员大多都具有相似的保守基序组成结构和相似的内含子/外显子分布.这一结果进一步证明PtbHLH亚家族分类的合理性,同时也暗示了属于相同亚家族的不同成员可能具有共同的进化起源,和相似的生理功能.根据注释信息,这167个bHLH基因的全部随机无规律的分布在19条染色体上,其中chr2是拥有PtbHLH最多的染色体.使用MCScanX软件,我们检测到PtbHLH60/PtbHLHL111和PtbHLH38/PtbHLHL108两个基因复制事件,说明串联复制在基因家族的扩张过程中起到一定的作用.从欧洲山杨与其他5种植物bHLH的进化关系推测有90%的PtbHLH源自于早期陆地植物,4%源自于藻类植物祖先.这一结论与Pires和Dolan[29]的研究结果一致.
虽然目前对欧洲山杨bHLH转录因子的功能尚未有深入的研究,但在此我们根据拟南芥中已知功能的AtbHLH和进化树中的聚类情况推测出39个PtbHLH的功能.例如,PtbHLH77、PtbHLH37和PtbHLH149与AtbHLH31在进化树中被聚为一簇,而AtbHLH31已知在拟南芥中具有控制花瓣大小的功能,因此推测PtbHLH77、PtbHLH37和PtbHLH149同样也具有相似的功能.这39个PtbHLH分别涉及繁殖和调节生长发育等过程,广泛地参与各类信号通路.PtbHLH116调节根毛生长,PtbHLH17和PtbHLH4参与植物精子与卵细胞的结合过程,PtbHLH97和PtbHLH51涉及油菜素甾醇信号转导途径等.除此之外,本研究对欧洲山杨根部干旱胁迫应答相关的bHLH进行了分析.4个PtbHLH基因在干旱根部组织和正常根部组织中差异表达,其中PtbHLH122是拟南芥bHLH130的直系同源基因,可能通过促进钾离子吸收,调节根部细胞渗透压以应对干旱胁迫.本研究对欧洲山杨bHLH转录因子家族进行全基因组水平的鉴定和分析,为进一步研究PtbHLH在欧洲山杨生长发育、抗逆调节中的作用提供了基础资源.
