超等长阻力训练对足踝屈肌肌构筑学及生物力学特征影响的研究
2021-05-31井兰香赵延治
井兰香 赵延治
摘 要:目的:探討超等长阻力训练对足踝屈肌横截面积、体积、羽状角等的影响,观察足底肌力迁移和跖屈肌三元素应变、刚度、做功等的变化,为深入理解训练引起肌-腱复合体构筑和生物力学特征变化的机制及意义提供新的思路。方法:将18名健康男性青年高级花样跳绳运动员随机分成实验组和对照组,对实验组实施跖、趾屈肌超等长阻力训练。于训练前、后对小腿三头肌、跟腱和足底固有肌做影像学测量,计算肌纤维厚度、横截面积、体积等;测试趾屈肌力,计算肌力增加率及肌力/横截面积比值;测试踝单关节活动和下肢多关节活动运动学、动力学及小腿三头肌影像学,计算腓肠肌外侧头各弹性元成分的应变、刚度、做功等参数。结果:训练后实验组小腿三头肌厚度、羽状角和足底固有肌横截面积均明显增加(P<0.05),跟腱横截面积没有明显变化(P>0.05);2~5趾屈力增加率明显大于1~5趾屈力增加率(P<0.01),2~5趾屈力/趾短屈肌横截面积的比值明显提高(P<0.05)。不同条件下腓肠肌外侧头肌-腱复合体各成分的被动刚度均未发生明显变化(P>0.05),主动刚度均明显提高(P<0.05),以大负荷(P<0.01)和快速应变(P<0.01)时提高最显著。肌纤维拉长应变明显降低(P<0.05),大负荷时降低最显著(P<0.05)。腱膜应变和滞后均未发生明显变化(P>0.05)。肌腱拉长速度未发生明显改变(P>0.05),缩短速度明显升高(P<0.05),弹性回缩率明显提高(P<0.05)。结论:超等长阻力训练能优化肌-腱复合体的构筑,较小的肌纤维体积增加即可获得较大的力量增幅;能增强支撑后期推进力,并稳固内侧纵弓;能增加肌-腱复合体的弹性,对大负荷和快速拉长-缩短周期运动表现更加有利。超等长阻力训练具有易化肌-腱复合体能量吸收和促进能量快速转化的双效性作用。
关键词:超等长阻力训练;肌-腱复合体;肌构筑;刚度;做功;能量
中图分类号:G804.63 文献标识码:A文章编号:1006-2076(2021)01-0038-10
Abstract:Objective: To explore effects of plyometric resistance training on plantar muscles cross-sectional area, thickness and pinnate angle and examine the changes in foot force transfer and strain, stiffness, work in flexor muscle. By this study we will present a newer researching way for insighting into the mechanism and significance of training-induced changes in muscle-tendon complex architecture and biomechanics. Methods: 18 male health young elite figure rope skipping athletes were randomly divided into experimental and control groups. Plyometric resistance training program was used in plantar and toe flexor muscles of experimental group subjects. Imageology of triceps surae muscle, achilles tendon, intrinsic foot muscles were measured before/after training and parameters of muscle thickness, cross-sectional area, volume etc. were calculated. Toe flexion force was tested for calculating the increasing rate of force and the force/cross-sectional area rate. Dynamics, kinetics, triceps surae muscle imageology during single-joint test in ankle and multi-joint test in leg were tested for calculating strain, stiffness, work etc. of each elastic element in lateral gastrocnemius. Results: After training, thickness and pinnate angle of triceps surae muscle and cross-sectional area of intrinsic foot muscles were all significantly increased (P<0.05). Theres no obvious change in tendon cross-sectional area (P>0.05). Increasing rate () of toe flexion force was more obviously in 2~5 than in 1~5 (P<0.01). 2~5 toe flexion force/flexor digitorum brevis cross-sectional area rate was significantly increased (P<0.05). Passive stiffness of each element in lateral gastrocnemius was not significantly changed when tested under different conditions (P>0.05) but their active stiffness were significantly increased (P<0.05), the most obvious exhibition were under heavy load (P<0.01) and fast strain (P<0.01) conditions. Fascicle elongation was significantly decreased (P<0.05), the most obvious exhibition was under heavy load (P<0.05) condition. Therere no significant changes in aponeurosis strain and hysteresis (P>0.05). Theres no significant change in tendon elongating velocity (P>0.05) but there were significant elevation in shortening velocity (P<0.05) and in elastic recoil rate (P<0.05). Conclusion: Architecture of muscle-tendon complex can be optimized through plyometric resistance training, moreover smaller increasing in volume can bring about greater force. Also propulsive force during later contact phase can be improved and medial longitudinal arch can be stabilized. Elasticity of muscle-tendon complex can be enhanced, which would be more advantageous to stretch-shortening cycle sport performance under heavy-loaded and fast conditions. Plyometric resistance training has double effect in facilitating energy absorption and expediting energy transformation.
Key words:plyometric resistance training; muscle-tendon complex; muscle architecture; stiffness; work; energy
人体在完成行走、跑、跳等下肢拉长-缩短周期运动(stretch-shortening cycle, SSC)时,通过支撑后期跖屈力与地面间相互作用而获得动能[1],此时小腿三头肌和跨越踝、跖趾关节的足底非固有肌共同产生内在的跖屈力矩使足跟离地,随即足底固有、非固有肌协同收缩产生趾屈力矩使前足获得腾空推进力。尽管静姿站立时趾长屈肌可以不参与维持人体平衡[2],但在SSC运动支撑期第二峰值期间此肌至少可产生61体重比的地面反作用力(ground reaction force, GRF)[3]。特定训练后足底固有、非固有肌产生的跖趾关节屈曲力矩的增加能使人体在行走、跑、跳和改变运动方向时加强跗骨间关节的稳定性[4-5]。若趾屈肌力不足,足弓柔性及刚性无法协调可导致膝和跗跖关节代偿性过度运动[6],甚至会增加踝关节功能代偿引起跖屈肌劳损[7]。只有具备了足够的趾屈肌力,才能于向心收缩期产生趾跖屈力矩以平衡GRF引起的趾背屈力矩[8],趾屈肌力在SSC运动支撑期起着不可或缺的作用。含有较多下肢SSC运动的项目,不仅要强化支撑期踝关节跖屈能力,还应重视跖趾关节屈曲功能的稳定[9]。为达到此目标,应采取积极措施优化跖屈、趾屈肌肌-腱复合体(muscle-tendon complex, MTC)的构筑,以提高其生物力学性能[10]。
MTC构筑和生物力学特征可通过阻力训练得到调整[11],单关节趾屈肌阻力训练,即能明显增加趾屈肌力与足底固有肌厚度的比值[12]。阻力训练负荷增加后,明显提高跖趾关节力矩而提高走、跑、跳的运动表现[13-14]。有研究认为,超等长训练提高运动表现的效果优于阻力训练[15],但两种训练在提高MTC构筑特征方面的效果接近[16]。前期研究则表明,中等负荷超等长阻力训练可同时提高肌力和爆发力[17],若联合跖屈和趾屈动作,对跖屈肌、足底固有和非固有趾屈肌进行超等长阻力训练,应更加有利于促进足/踝部肌-骨系统的稳定性,并可对下肢多关节运动表现产生积极效应。故为了探究超等长阻力训练后足/踝部屈肌MTC的构筑和不同条件下踝关节运动时跖屈肌MTC的生物力学特征的变化,从MTC构筑和三元素生物力学角度阐明该训练提高运动表现的机制,本研究对足/踝部超等长阻力训练前、后高级花样跳绳运动员跖屈肌、趾屈肌构筑及力量变化进行测量,对踝单关节运动和下肢多关节运动过程进行足/踝部运动学、动力学、影像学等测量,以获得MTC可收缩成分、并联弹性元、串联弹性元生物力学数据,藉此为深入理解训练引起肌-腱复合体构筑及生物力学特征变化对SSC运动表现所起作用的机制和意义提供新的方法与思路。
1 对象与方法
1.1 实验对象
随机选取某高校体育学院健康男性青年高级花样跳绳运动员作为调查对象,于实验前发放调查问卷,确定受试者的身体机能状态良好。受试者须具有3年以上专业跳绳经历,连续跳跃时跖趾关节屈曲具有较大速度梯度,双脚交替单摇、双脚并单摇、双摇连续跳绳成绩分别不低于270次/min、 190次/min 、130次/min,能连续、无失误地完成三摇跳50次及以上,且均未经历过系统地足/踝部超等长阻力训练以及阻力训练。近5年内不存在下肢损伤史,踝关节功能稳定,无足部内、外翻畸形及拇外翻等现象,踝关节及跖趾关节跖/背屈动作不存在活动受限及其它不适。实验前经生物学伦理学委员会审查、批准后,受试者自愿参与本实验并签署知情协议书。
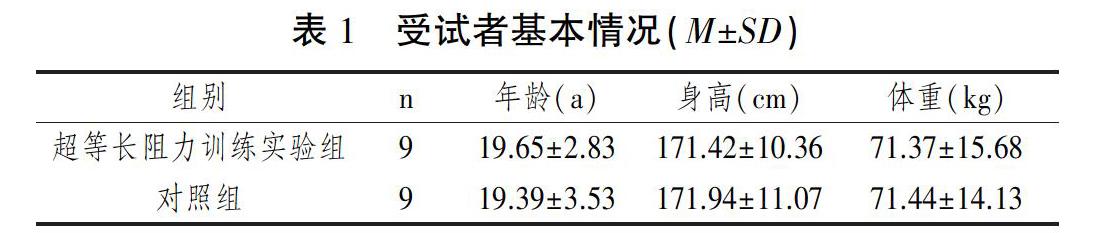
使用G*Power 3.1.9.2进行最小样本量计算。在进行重复测量的双因素方差分析假设检验时,为使统计结果达到80统计功效、显著水平为0.05、效应量为0.4、非球形相关0.5,需要最少样本量16人。本研究将符合纳入标准要求的18名受试者(统计功效=85)随机分成日常训练+足/踝部超等长阻力训练组(n=9)和日常训练对照组(n=9)。两组基本情况描述性分析见表1。两组间年龄、身高、体重三个变量数据均数间进行独立样本t检验的P值均大于0.05,两组间没有统计学差异。
1.2 训练方案
受试者以30最大等长自主收缩(Maximum Voluntary Contraction, MVC) 负荷完成双足连续跳跃训练,保持踝关节背屈绝对角度大于90°,前足着地后以足底肌和跖屈肌离心收缩缓冲,离地前跖趾关节屈肌尽力快速向心收缩跳起,向心收缩期具有明显“抓地”动作。支撑期髋、膝关节自然伸展,将这2个关节的角位移控制在0°~20°。共进行12周每周3次的训练,训练组数为3组,每组10个,组间休息2分钟。由专业教练全程指导训练,确保安全、有效。对照组除日常训练外不增加任何形式的下肢训练,实验期间两组受试者不改变原有训练日程。
1.3 测试总体流程
分别于训练前、训练中期每隔2周、训练后测量每位受试者跖屈+趾屈MVC值。训练前、后1周内采集小腿三头肌和足底固有肌影像和趾屈肌力数据。
1.4 数据采集及处理
1.4.1 MTC构筑
采用美国GE公司生产的型号为Voluson E8的黑白B型超声影像采集系统一台,用于采集小腿三头肌、足底固有肌的二维黑白超声影像。外置9L-D型、扫描频率为3.5 MHz的线阵探头,其测量边界长度×深度为43×70 mm,采样频率设定为50 Hz。
测量腓肠肌内侧头(Medial Gastrocnemius, GM)、腓肠肌外侧头(Lateral Gastrocnemius, LG)肌纤维厚度和羽状角的位置约为小腿长度近侧端30水平,测量比目鱼肌(Soleus Muscle, Sol)肌纤维厚度和羽状角的位置约为小腿长度近侧端50水平,分别将探头足印中心置于MG、LG、Sol外側半肌腹最发达处皮肤表面。测量跟腱横截面积时将探头足印中心置于外踝水平,跟腱中心处皮肤表面[18-19]。图1-A中的角α、β分别代表LG、Sol肌纤维与腱膜的夹角,即羽状角。图1-B中跟腱标识的范围为跟腱横截面。
测得趾短屈肌最发达处横截面积之后,以50足长为基线,向足底近端、远端方向每隔1 cm划一条平行线,测量1 cm间隔的足底固有肌连续影像图,依据图像测量出每间隔1 cm处趾短屈肌的横截面积(图2)。以连续趾短屈肌横截面积之和乘以总长度得到该肌的体积[20-21]。其余足底固有肌如拇展肌、小趾展肌、拇短屈肌,均测量其最发达处的横截面积。
1.4.2 趾屈肌力
采用普通掌握式测力计测量趾屈肌力[22],以裸足第1~5个跖趾关节屈伸的方式做简单准备活动后,将非利侧足抬起,利侧单足站立位双手推墙姿势,保持踝背屈位70°~90[23]。先测量2次第1~5趾屈力,记录最大力值。间隔2 min后,再测量2次第2~5趾尽力,记录最大力值。
1.4.3 纵跳
测量自15 cm高度下落后下肢自然伸展缓冲动作(Drop Landing, DL)和60 cm高度下落下肢屈曲-跳起动作(Drop Jump, DJ)。测量前慢跑5 min后,以DL、DJ的先后顺序,同步采集踝关节运动学、动力学、LG影像学。2种纵跳各测量2次,间隔2 min。DL取充分缓冲时的数据、DJ取跳跃最高时的数据。
(1)纵跳动力学。瑞士Kistler公司生产三维测力系统1台,型号为9286BA,边界范围60 cm×40 cm×22cm。用于采集着地期三维GRF,采样频率为1000 Hz。对三维GRF数据进行10 Hz~50 Hz截止频率的滤波。
(2)纵跳运动学。瑞典生产的Qualisys MCU500三维运动捕捉系统,6个红外高速摄像头,用于采集DL和DJ的运动学数据,采样频率为200 Hz。
热身后,更换实验用紧身衣及统一品牌鞋袜,根据运动学数据采集系统模型标准和3-D 软件建模要求,为受试者粘贴49处主要标记点,Mark反光球直径为12.5 mm,拍摄受试者静态站姿并进行3-D建模(图3-A)。
动态测试时,仅对下肢进行主要标记,共28处。运动学、动力学坐标系为:Z-垂直轴,Y-矢状轴,X-冠状轴。将膝内/外上髁连线中点-内/外踝连线中点-第2、5跖趾关节连线中点定义为踝关节角(图3-B)。
VISUAL3-D图像后期处理时(美国c-motion公司,Visual3-D Version 3.34.0),采用截止频率为7 Hz的二阶双向低通滤波,通过3-D软件模型进行数据计算,获取踝关节关节中心运动轨迹后,经逆动力学法计算获得踝净关节力矩(Net joint moment, NJM),以运用于DL和DJ支撑期LG串联弹性元生物力学参数的计算。
1.5 参数的获得
1.5.1 可收缩成分刚度
Contrex等速肌力仪(瑞士CMV AG公司生产,型号为PM-MJ/LP/WS/TA/TP,采样频率4000 Hz),与超声影像仪同步测量,计算LG肌纤维被动和主动刚度。
慢拉式被动刚度:测量前不热身,受试者仰卧于测试平台,保持膝关节伸展,足固定,踝关节置于跖屈位10°(踝解剖位为0°),肌完全放松。测量开始后,以5°/s的恒定速度连续被动拉伸小腿三头肌至踝背屈位10°。按F肌纤维=k·TQ·d-1式获得肌纤维力量,k为LG生理横截面积占小腿三头肌总生理横截面积的比率,取值15.4;TQ为踝关节被动力矩Torque;d为小腿三头肌总肌腱的力臂,取值5 cm[24]。0°~背屈位10°范围内肌力-肌纤维长度变化曲线斜率为肌纤维的被动刚度[18]。
快拉式主动刚度:人体固定的方法及关节方位定义同被动刚度测量。5 min常规热身后受试者完成2次跖屈位10°条件下的等长背屈MVC,取此时最大力矩值作为快速拉伸测量时的负荷依据。受试者分别以30、50、70MVC时对应的力矩为负荷,完成自跖屈位10°至背屈位10°范围内爆发性等张背屈。为避免牵张反射的影响,记录背屈开始后60 ms时间内最大肌纤维长度[25]。将跖屈位10°~2°范围内肌纤维力-肌纤维长度变化曲线斜率定义为肌纤维主动刚度[18]。每种负荷背屈动作均记录2次,各取平均值。肌纤维力量、长度测量方法同被动刚度。
1.5.2 并联弹性元刚度及滞后
测量前经5 min热身及适应性踝跖屈后,共测量两种收缩形式:(1)渐变式用力,跖屈肌自放松状态逐渐加载应力,5 s内增至MVC,随后逐渐卸载应力5 s内放松;(2)爆发式用力,跖屈肌以最快速度加载和卸载应力。每个动作重复2次,间隔2 min,取最大值。
与跖屈力矩数据同步采集LG影像,测得腱膜长度变化值,结合F肌纤维=k·TQ·d-1 和F腱膜=F肌纤维·cosα公式计算腱膜的力值,其中,α为LG羽状角。腱成分力-腱成分长度变化關系曲线如图4所示,本研究将加载应力达50MVC以上时曲线的斜率定义为腱成分刚度,将滞后环面积与加载应力时曲线下面积的比值定义为滞后[26]。
1.5.3 串联弹性元刚度及做功
DL和DJ动作测试时,与下肢运动学、动力学数据同步采集LG超声影像学。
依据受试者个体的下肢环节长度及LG缩短与拉长比值估测其MTC总长度[27-28],按L腱=LMTC-L肌纤维×cosα计算MTC结构中与LG肌纤维串联肌腱的长度,其中LMTC 表示MTC总长度,L肌纤维 表示肌纤维长度,α为LG羽状角。自脚尖落地时刻至离心收缩结束期间L腱最大拉长变化量即为拉长应变,自最大拉长至缓冲结束期间L腱最大缩短变化量即为缩短应变。离心收缩期间L腱最大变化量对离心收缩时间求微分,计为肌腱拉长应变速率v拉长;DL离心结束时刻至恢复静止期间和DJ向心收缩期间L腱最大变化量对相应时间求微分,计为肌腱缩短应变速率v缩短。
按F腱=k·NJM·d-1计算肌腱力[29],其中NJM表示踝跖屈力矩。以峰值F腱除以离心期L腱最大拉长变化量计算被动刚度,以峰值F腱除以离心结束至静止期或向心期L腱的最大主动回弹缩短变化量计算主动刚度[30]。
以F腱乘以v拉长或v缩短获得功率,再将功率对相应时间求积分得肌腱拉长/缩短做功,缩短做功与拉长做功的比值,即为肌腱的弹性回缩率[31]。
1.6 统计方法
用SPSS 24.0统计软件对实验结果数据进行重复测量的双因素方差分析,观察自变量(2种训练方式×2个测试时间)对肌-腱复合体形态学、生物力学特征、下肢及下肢关节动力学特征的影响。经studentized residual分析、Shapiro-Wilk检验,判断两组数据是否服从正态分布,有无异常值。当方差分析输出结果列表中出现干预×时间交互效应P<0.05时,则分析2个自变量的单独效应,以Post hoc-Bonferroni和Tukey两两比较结果。经单因素方差分析比较实验组不同负荷快拉伸拉长应变和主动刚度的差异,比较训练后两组趾屈肌力增加率间的差异性,以Post hoc-LSD两两比较结果。经独立样本t检验进行组间受试者基本情况比较。统计结果以均数±标准差(M±SD)表示,显著性水平为0.05。
2 实验结果
2.1 訓练前后足踝屈肌构筑特征比较
双因素方差分析干预、时间、交互3自变量对跟腱横截面积影响的主体间效应的检验P值分别为0.532、0.844、0.657,主效应和交互效应均不具有统计学意义。对MG、LG、Sol的肌纤维厚度、羽状角影响主体间效应的检验P值均<0.05,主效应及交互效应显著。
单独效应为:训练前两组间比较MG、LG、Sol的肌纤维厚度(P值分别为0.718、0792、0.903)、羽状角(P值分别为0.802、0.743、0.822)均无统计学意义。训练后实验组MG、LG、Sol的肌纤维厚度(P值分别为0.021、0. 020、0.027)、羽状角(P值分别为0.013、0.013、0.011)均明显大于对照组。对照实验中,时间因素对MG、LG、Sol肌纤维厚度(P值分别为0.634、0729、0.817)、羽状角(P值分别为0.843、0.761、0.789)的影响不具有统计学意义。干预实验中,时间因素对MG、LG、Sol肌纤维厚度(P值分别为0.033、0.021、0.018)、羽状角(P值分别为0.022、0.018、0.020)的影响具有统计学意义,训练后MG、LG、Sol肌纤维厚度(P值分别为0.038、0.031、0.030)、羽状角(P值分别为0.023、0.021、0.019)均显著升高(表2)。
双因素方差分析干预、时间、交互3自变量对足底固有肌横截面积及趾短屈肌体积影响的主体间效应的检验P值均<0.05,主效应及交互效应显著。
单独效应为:训练前两组间比较趾短屈肌横截面积(P=0.818)、体积(P=0.911)无统计学意义,比较拇短屈肌(P=0.917)、拇展肌(P=0.882)、小趾展肌(P=0.701)横截面积均无统计学意义。训练后实验组趾短屈肌横截面积(P=0.009)、体积(P=0.003)均非常明显的大于对照组,拇短屈肌(P=0.026)、拇展肌(P=0.032)、小趾展肌(P=0.011)横截面积均明显大于对照组。对照实验中,时间因素对趾短屈肌横截面积(P=0.675)、体积(P=0.737)的影响无统计学意义,对拇短屈肌(P=0.576)、拇展肌(P=0.815)、小趾展肌(P=0.721)横截面积的影响均无统计学意义。干预实验中,时间因素对趾短屈肌横截面积(P=0.005)、体积(P=0.007)的影响有统计学意义,对拇短屈肌(P=0.026)、拇展肌(P=0.025)、小趾展肌(P=0.021)横截面积的影响均有统计学意义。训练后实验组短屈肌横截面积(P=0.008)、体积(P=0.003)增加非常明显,拇短屈肌(P=0.025)、拇展肌(P=0.029)、小趾展肌(P=0.012)横截面积均明显增加(表3)。
2.2 训练前后趾屈肌力特征比较
双因素方差分析干预、时间、交互3自变量对2~5趾屈力/趾短屈肌横截面积比值影响的主体间效应的检验P值分别为0.017、0.025、0.022,主效应及交互效应显著。
单独效应为:训练前两组间比较2~5趾屈力/趾短屈肌横截面积的比值无统计学意义(P=0.898),训练后实验组明显大于对照组(P=0.014)。对照实验中,时间因素对2~5趾屈力/趾短屈肌横截面积比值的影响无统计学意义(P=0.834)。干预实验中,时间因素对2~5趾屈力/趾短屈肌横截面积比值的影响具有统计学意义(P=0.021),训练后明显增加(P=0.015)(图5)。
单因素方差分析训练后实验组2~5趾屈力增加率 (等于训练后趾屈肌力减去训练前趾屈肌力的差除以训练前趾屈肌力)明显大于对照组(P=0.021),1~5趾屈力增加率非常明显的大于对照组(P=0.002)。训练后实验组2~5趾屈力增加率非常明显的大于1~5趾屈力增加率(P=0.006)(图6)。
2.3 训练前后MTC各成分生物力学特征比较
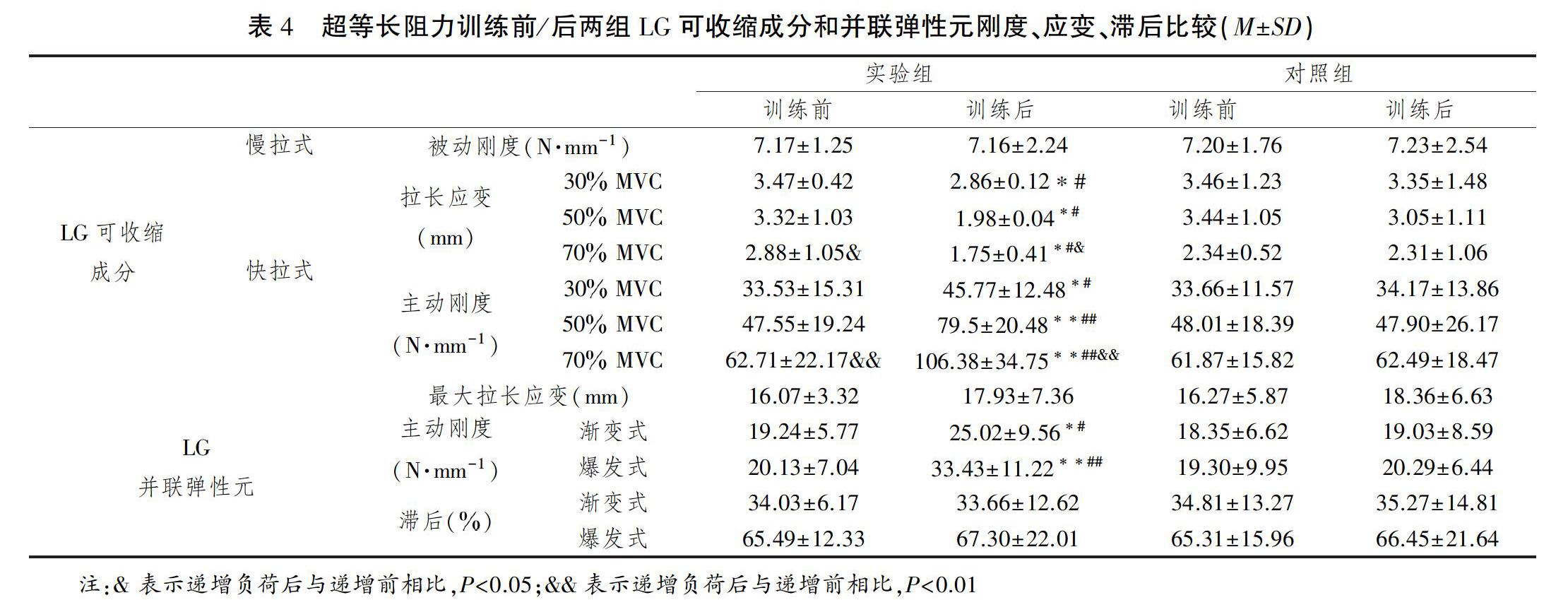
双因素方差分析干预、时间、交互3自变量对LG可收缩成分被动刚度、LG并联弹性元最大拉长应变、滞后影响的主体间效应的检验P值均>0.05,主效应和交互效应均不具有统计学意义。对LG可收缩成分拉长应变、二成分主动刚度影响主体间效应的检验P值均<0.05,主效应及交互效应显著。
单独效应为:训练前两组间比较LG可收缩成分30、50、70MVC负荷快拉式拉长应变(P值分别为0.679、0.665、0.754)及其主动刚度(P值分别为0.902、0.673、0.706)均无统计学意义,比较LG并联弹性元渐变式、爆发式主动刚度(P值分别为0.574、0.649)无统计学意义。训练后实验组LG可收缩成分30、50、70MVC负荷快拉式拉长应变(P值分别为0.017、0.021、0.015)明显低于对照组, 30MVC负荷主动刚度明显高于对照组(P=0.018),50、70MVC负荷主动刚度非常明显的高于对照组(P值分别为0.007、0.000)。训练后实验组LG并联弹性元渐变式主动刚度明显高于对照组(P=0.015)、爆发式主动刚度非常明显的高于对照组(P=0.003)。对照实验中,时间因素对LG可收缩成分30、50、70MVC负荷快拉式拉长应变(P值分别为0.717、0.622、0.729)及其主动刚度(P值分别为0.833、0.732、0.736)的影响均无统计学意义,对LG并联弹性元渐变式、爆发式主动刚度的影响无统计学意义(P值分别为0.644、0.652)。干预实验中,时间因素对LG可收缩成分30、50、70MVC负荷快拉式拉长应变(P值分别为0.017、0.017、0.012)及其主动刚度(P值分别为0.013、0.009、0.001)的影响均具有统计学意义,对LG并联弹性元渐变式、爆发式主动刚度的影响具有统计学意义(P值分别为0.014、0.002)。训练后实验组LG可收缩成分30、50、70MVC负荷快拉式拉长应变明显降低(P值分别为0.016、0.017、0.014),30MVC负荷主动刚度明显提高(P=0.011)、50、70MVC负荷主动刚度提高非常明显 (P值分别为0.002、0.000)。训练后实验组LG并联弹性元渐变式主动刚度明显提高(P=0.013)、爆发式主动刚度提高非常明显(P=0.004)。
单因素方差分析实验组70MVC负荷时LG可收缩成分快拉式拉长应变显著低于30、50MVC负荷时(训练前不同负荷间比较的P值分别为0.021、0.013,训练后不同负荷间比较的P值分别为0.022、0.016);70MVC负荷时LG可收缩成分快拉式主动刚度非常显著的高于负荷时30、50MVC负荷时(训练前不同负荷间比较的P值分别为0.002、0.003,训练后不同负荷间比较的P值分别为0.001、0.000)(表4)。
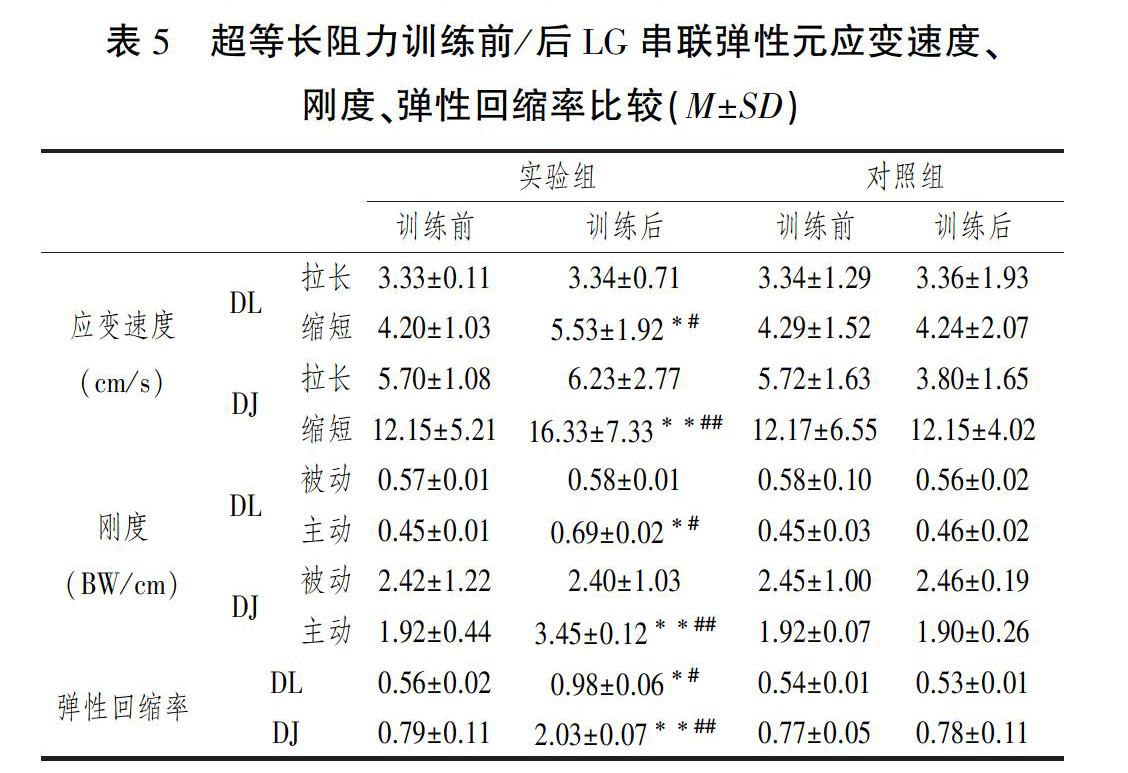
双因素方差分析干预、时间、交互3自变量对DL和DJ两种纵跳LG串联弹性元拉长应变速度及被动刚度影响的主体间效应的检验P值均>0.05,主效应和交互效应均不具有统计学意义。对DL和DJ两种纵跳LG串联弹性元缩短应变速度、主动刚度、弹性回缩率影响主体间效应的检验P值均<0.05,主效应及交互效应显著。
单独效应为:训练前两组间比较DL过程LG串聯弹性元缩短应变速度(P=0.579)、主动刚度 (P=0.634)、弹性回缩率(P=0.722)均无统计学意义,比较DJ过程LG串联弹性元缩短应变速度(P=0.622)、主动刚度 (P=0.653)、弹性回缩率(P=0.715)均无统计学意义。训练后DL过程LG串联弹性元缩短应变速度(P=0.019)、主动刚度 (P=0.021)、弹性回缩率(P=0.032)均明显大于对照组,DJ过程LG串联弹性元缩短应变速度(P=0.001)、主动刚度 (P=0.006)、弹性回缩率(P=0.005)均非常明显的大于对照组。
对照实验中,时间因素对DL过程LG串联弹性元缩短应变速度(P=0.643)、主动刚度 (P=0.674)、弹性回缩率(P=0.692)的影响均无统计学意义,对DJ过程LG串联弹性元缩短应变速度(P=0.731)、主动刚度 (P=0.676)、弹性回缩率(P=0.802)的影响均无统计学意义。干预实验中,时间因素对DL过程LG串联弹性元缩短应变速度(P=0.013)、主动刚度 (P=0.014)、弹性回缩率(P=0.022)的影响均具有统计学意义,对DJ过程LG串联弹性元缩短应变速度(P=0.004)、主动刚度 (P=0.006)、弹性回缩率(P=0.008)的影响均具有统计学意义。训练后实验组DL过程LG串联弹性元缩短应变速度(P=0.024)、主动刚度 (P=0.026)、弹性回缩率(P=0.032)均明显增加,DJ过程LG串联弹性元缩短应变速度(P=0.005)、主动刚度 (P=0.006)、弹性回缩率(P=0.008)均增加非常明显(表5)。
3 讨 论
本研究发现超等长阻力训练后跖屈肌、趾屈肌肌纤维构筑均有一定程度的增加,跟腱构筑不受训练的影响,表明踝跖屈能力的提高不是通过跟腱的肥大实现的,可能是跟腱内部发生了组织重建[32]。足底固有肌力量和体积增加,足底力量向足底外侧迁移,且较小的体积增加可带来较大肌力的提高。不同条件下测试小腿三头肌MTC各成分的生物力学特征出现了特定的规律性变化:训练后被动刚度不变,主动刚度明显增加;滞后不变,弹性回缩率明显提高。训练后越是在大负荷和快速MTC应变时,肌纤维的拉长越能够有效控制,并联、串联弹性元的主动刚度、回弹做功就越突出。提示,超等长阻力训练通过优化MTC构筑和生物力学特征,对提高受试者足踝功能稳定、促进能量吸收和快速转化产生作用。
3.1 优化足踝屈肌构筑的益处
由研究结果可知,超等长阻力训练增加趾屈肌肌纤维厚度、体积,较小的足底固有肌构筑增加便可获得较大的趾屈力量增幅。本实验足踝受训肌MTC可收缩成分构筑的这种适应性变化,可能的解释是因为腱成分主动刚度增加的缘故[33]。腱成分主动刚度增加,并联弹性元主动刚度增加,变得强劲有力,与肌力增强相辅相成。另一种解释是训练可能会使肌纤维本身肌小节数量增加而提升构筑[34]。这会使肌肉力量在有更多横桥位点结合时变得强大。研究结果证实,无论何种理由引起肌纤维构筑提高,其所带来的肌力增加效应都是比较可观的。
训练后实验组2~5趾屈力增加率大于1~5趾屈力增加率,表明足底肌力增加的同时向外侧纵弓侧发生了迁移,能更好地协调足旋前和旋后动态平衡。积极旋后可避免拇屈肌、拇展肌过度使用而发生劳损,保护内侧纵弓。加强大拇指的屈和外展力不仅在足旋前时延展前足的有效接触面积,使SSC支撑后期获得较大的推进力,还在完成第一跖趾关节屈和外展时促进旋后。可见,拇指屈曲、外展力量增加使足踝在旋前/旋后的合理动态平衡之中更加稳定[35]。因足底固有肌对内侧纵弓起着支撑作用[36],一旦这些肌疲劳加重旋前,则会造成内侧纵弓压力过大[37]。足底肌力向外侧迁移,阻止过度旋前保护内侧纵弓,防止拇展肌被过度使用[14]。尤其是,从事反复高频跳跃动作、常发内侧纵弓疼痛的高级跳绳运动员,趾短屈肌构筑显著提升,足底压力向外侧迁移无疑对足踝稳定有着积极意义。
3.2 提高足踝屈肌主动刚度的益处
某种训练最理想的状态是减少能量损失,本研究的结果并未出现MTC各弹性元滞后环的明显变化,说明超等长阻力训练的主要目标并不致力于如何降低MTC能量损失,但也不会因此造成能量损失的增加。本研究的结果是训练后DL、DJ向心期肌腱缩短应变速度明显增加所带来的益处——弹性回缩率提高。同时,因羽状角的增大,肌纤维密度增加使投影于腱膜的实际有效力值也是增加了的,而腱膜应变不受训练的影响,使腱膜刚度明显增加,特别是爆发性测试时刚度增加更显著。这些现象表明,超等长阻力训练是通过提高MTC腱成分的弹性达到提高能量转换利用率的。
因小腿三头肌可收缩成分羽状角构筑提升,使肌纤维拉长应变能够在快拉式运动中得到有效控制,随着负荷的增加而产生较强劲的主动刚度,负荷越大肌纤维刚度越大。并联弹性元的主动刚度因此得到加强,尤其是爆发性用力时更加明显。MTC腱成分的主动刚度是调节肌纤维应变的关键因素[26],因为肌腱是离心期能量吸收的“缓冲器”,能够调节肌纤维拉长应变比率和峰值应力[38];肌腱还能起到“功率衰减器”的作用,在MTC快速和过力拉长应变过程中,有效保护肌纤维[39]。本研究结果表明,无论是在踝单关节运动还是下肢多关节运动的过程中,MTC在快速拉长和大负荷条件下肌纤维均能明显控制拉长应变,这进一步验证了肌腱主动刚度增加的积极意义。
3.3 足踝屈肌生物力学特征变化对SSC过程的益处
训练后MTC串联弹性元生物力学特征发生了不同的变化,结果呈现出双效性作用:有效控制离心期收缩速度梯度,提高向心期收缩效益。上述MTC生物力学特征的表现对下肢快速SSC过程极其有利:不仅促使运动员能更好地利用MTC的粘弹性帮助缓冲,落地变得更加轻盈,避免关节过于僵硬,还降低地面反作用力对关节的直接作用,使能量吸收、储存更加充分。在下肢快速回弹过程中,还可使储存的能量得以充分快速释放,从而使提高运动表现成为可能。
4 结 论
超等长阻力训练能优化肌-腱复合体的构筑,较小的肌纤维体积增加即可获得较大的力量增幅;能增强SSC支撑后期推进力,并稳固内侧纵弓;能增加肌-腱复合体的弹性,对大负荷和快速拉长-缩短周期运动表现更加有利。超等长阻力训练具有易化肌-腱复合体能量吸收和促进能量快速转化的双效作用。
参考文献:
[1]Dominic J F, Gregory S S. Human medial gastrocnemius force-velocity behavior shifts with locomotion speed and gait[J]. PNAS, 2012, 109(3):977-982.
[2]Junichiro Y, Keiji K. Force-generating Capacity of the Toe Flexor Muscles and Dynamic Function of the Foot Arch in Upright Standing[J]. J Anat,2019, 234(4):515-522.
[3]Jacob H A. Forces acting in the forefoot during normal gait—an estimate[J]. Clin Biomech (Bristol, Avon),2001, 16(9):783-792.
[4]Goldmann J P, Sanno M, Willwacher S, et al. The potential of toe flexor muscles to enhance performance[J]. J Sports Sci,2013, 31(4):424-433.
[5]Xiao S L, Zhang X N, Deng L Q et al. Relationships Between Foot Morphology and Foot Muscle Strength in Healthy Adults[J]. Int J Environ Res Public Health, 2020, 17(4):1274-1274.
[6]张峻霞, 司莹, 苏海龙, 等. 湿滑环境下第一跖趾关节弯曲对人体行走步态稳定性的影响[J]. 生物醫学工程学杂志, 2015, 32(1):38-42.
[7]万祥林, 曲峰, 杨辰, 等. 人体跖趾关节活动受限对行走时下肢生物力学特征的影响[J]. 中国运动医学杂志, 2015, 34(6):553-558.
[8]Nicole L G, Charlotte M, Daniel S, et al. An investigation of the dynamic relationship between navicular drop and first metatarsophalangeal joint dorsal excursion[J]. J Anat,2013, 222(6):598-607.
[9]张海斌. 排球运动员急停起跳动作中跖趾关节的生物力学分析[J]. 山东体育学院学报, 2014, 10(5):71-75.
[10]Alonso F D, Docampo B P, Martinez F J. Changes in muscle architecture of biceps femoris induced by eccentric strength training with nordic hamstring exercise[J]. Scand J Med Sci Sports,2018, 28(1):88-94.
[11]Hashimoto T, Sakuraba K. Strength training for the intrinsic flexor muscles of the foot:effects on muscle strength, the foot arch, and dynamic parameters before and after the training[J]. J Phys Ther Sci, 2014, 26(3):373-376.
[12]Koyama K, Hirokawa M, Yoshitaka Y, et al. Toe Flexor Muscle Strength and Morphological Characteristics of the Foot in Judo Athletes[J]. Int J Sports Med, 2019, 40(4):263-268.
[13]Kazunori O, Shusaku K, Sadaaki O, et al. Does the weakening of intrinsic foot muscles cause the decrease of medial longitudinal arch height[J].J Phys Ther Sci,2017, 29(6):1001-1005.
[14]Franois F, Sami K, Olivier G, et al. Effects of Combined Foot/Ankle Electromyostimulation and Resistance Training on the In-Shoe Plantar Pressure Patterns during Sprint in Young Athletes[J]. J Sports Sci Med, 2011, 10(2):292-300.
[15]Hammami M, Gaamouri N, Shephard R J, et al. Effects of Contrast Strength vs. Plyometric Training on Lower-Limb Explosive Performance, Ability to Change Direction and Neuromuscular Adaptation in Soccer Players[J]. J Strength Cond Res, 2019, 33(8):2094-2103.
[16]McKinlay B J, Wallace P, Dotan R, et al. Effects of Plyometric and Resistance Training on Muscle Strength, Explosiveness, and Neuromuscular Function in Young Adolescent Soccer Players[J]. J Strength Cond Res, 2018, 32(11):3039-3050.
[17]井兰香, 刘 宇, 田石榴. 阻力训练及超等长阻力训练对下肢肌肉活性和力量的影响[J]. 中国体育科技, 2011, 47(6):90-93.
[18]Kubo K, Teshima T, Hirose N, et al. A cross-sectional study of the plantar flexor muscle and tendon during growth[J]. Int. J. Sports Med, 2014, 35(10):828-834.
[19]Mogi Y, Torii S, Kawakami Y, et al. A cross-sectional study on the mechanical properties of the Achilles tendon with growth[J]. Eur J Appl Physiol, 2018, 118(1):185-194.
[20]Takashi A, Kota T, Miyuki N, et al. Relationships of ultrasound measures of intrinsic foot muscle cross-sectional area and muscle volume with maximum toe flexor muscle strength and physical performance in young adults[J]. J Phys Ther Sci, 2016, 28(1):14-19.
[21]許正勇, 张先熠. 足部内在肌形态的测量和与人体静态平衡稳定性的关系[J].体育学刊,2016, 23(2):133-137.
[22]Soma M, Murata S, Kai Y, et al. An examination of limb position for measuring toe-grip strength[J]. J Phys Ther Sci, 2014, 26(12):1955-1957.
[23]JunichiroY, Keiji K. Relation between the ankle joint angle and the maximum isometric force of the toe flexor muscles[J]. Journal of Biomechanics, 2019, 85(6):1-5.
[24]Winter D A. Biomechanics and motor control of human movement[M]. 4th Edition. Waterloo:University of Waterloo Press,2009.
[25]Kubo K, Miyazaki D, Ikebukuro T, Yata H, Okada M, Tsunoda N. Active muscle and tendon stiffness of plantar flexors in sprinters[J]. J Sports Sci, 2017, 35(8):742-748.
[26]Kubo K, Miyazaki D, Yata H, et al. Is the stiffness of human muscle and tendon structures related to muscle fiber composition in vivo[J].J Sports Med Phys Fitness,2018, 58(5):622-629.
[27]Hawkins D, Hull M L. A method for determine lower extremity muscle-tendon lengths during flexion-extension movements[J]. J Biomech, 1990; 23(5):487-494.
[28]單大卯, 魏文仪. 股四头肌长度和力臂的实验性研究[J]. 天津体育学院学报, 2005, 20(2):17-20.
[29]Kubo K. Active muscle stiffness in the human medial gastrocnemius muscle in vivo[J]. J Appl Physiol, 2014, 117(9):1020-1026.
[30]Kubo K, Miyazaki D, Yamada K, et al. Passive and active muscle stiffness in plantar flexors of long distance runners[J]. J Biomech, 2015, 48(10):1937-1943.
[31]Lai A, Schache A G, Lin Y C, et al. Tendon elastic strain energy in the human ankle plantar-flexors and its role with increased running speed[J]. J. Exp. Biol, 2014, 217(17):3159-3168.
[32]艾进伟, 黄昌林, 韩叶萍, 等. 步兵踝关节肌力和跟腱横截面积与强化训练的关系[J]. 中国组织工程研究与临床康复, 2007, 11(5):984-986.
[33]Butterfield T A, Herzog W. Quantification of muscle fiber strain during in vivo repetitive stretch-shortening cycles[J]. J Appl Physiol (1985), 2005, 99(2):593-602.
[34]Proske U, Morgan D L. Muscle damage from eccentric exercise:mechanism, mechanical signs, adaptation and clinical applications[J]. J Physiol, 2001, 537(Pt 2):333-345.
[35]杨平, 蔡丽飞. 足过度旋前对人体力线的影响及治疗方法[J]. 中国康复理论与实践, 2016, 22(1):72-73.
[36]Fiolkowski P M, Brunt D S, Woo R, et al. Intrinsic pedal musculature support of the medial longitudinal arch:an electromyography study[J]. J foot ankle surg, 2003, 42(6):327-333.
[37]Headlee D L, Leonard J L, Hart J M. Fatigue of the plantar intrinsic foot muscles increases navicular drop[J]. J electromyogr kines, 2007, 18(3):420-425.
[38]Roberts T J, Azizi E. The series-elastic shock absorber:tendons attenuate muscle power during eccentric actions[J]. J Appl Physiol (1985), 2010, 109(2):396-404.
[39]Konow N, Azizi E, Roberts T J. Muscle power attenuation by tendon during energy dissipation[J]. Proc Biol Sci, 2012, 279(1731):1108-1113.
