早材管孔分布对环孔材栎木蠕变特性的影响
2021-05-31尹业桥侯俊峰姜志宏俞友明
尹业桥,侯俊峰,姜志宏,俞友明
(浙江农林大学工程学院,杭州 311300)
木材作为一种可持续发展的建筑材料,也是一种复杂的高分子多孔材料,其主要化学成分为纤维素、半纤维素、木质素等,由于具有黏弹性而会表现出蠕变特性[1-3]。
木材的黏弹性主要取决于温度、含水率、应力水平和木材构造等因素。前人研究表明,试验温度的升高会使得试样在试验过程中产生的蠕变增大[4-6]。随着温度的升高,木材中木质素和半纤维素等无定形聚合物分子间相互作用的化学键键能增加,相互作用力减弱,导致分子的活动空间增大,使得木材蠕变速率加快[7]。水分作为木材中的增塑剂,通过与木质素中亲水性基团(羟基、羧基和羰基等)的结合,导致木材细胞壁中的分子链发生断裂,分子链间的相互作用力减弱,伸展性流动性增强,分子运动速率加快,木材蠕变形变增大[8-9]。此外,木材在恒定应力作用下,随着应力水平的增大,其蠕变应变也随之增大[7,9-10]。
导管是阔叶树材的重要组成细胞类型,管孔是导管在阔叶材横断面上的孔洞。根据管孔的分布类型,木材可分为环孔材、半环(散)孔材、散孔材等。然而,目前关于木材管孔分布,尤其是环孔材早材管孔带对阔叶树材黏弹性的影响鲜见相关报道。因此,本研究在表征环孔材栎木横切面早材管孔带分布特性的基础上,探究不同温度条件下早材管孔带对环孔材栎木板材弯曲蠕变行为的影响,利用Burger模型和五参数模型对其蠕变特性进行模拟预测,以期为木材干燥、木材弯曲加工成型、木质材料热压成型和木结构设计提供理论依据。
1 材料与方法
1.1 试验材料
环孔材栎木(Quercusspp.)取自浙江省湖州市南浔建材市场,试样尺寸为40 mm(纵向)×12 mm(弦向)×2.2 mm(径向)。根据导管的分布位置制备了2种试样:试样A为早材管孔带位于试样中部的木材试样,试样B为对照组-无早材管孔带木材试样。在试样制备过程中,要求试样端面结构均匀一致,且纤维方向平行于试样的4个侧面,试样两端面和上下表面分别对应于横切面和弦切面。
1.2 试样早材管孔带和早材管孔组织比量表征
采用扫描电子显微镜(SEM,SS-550型,日本日立有限公司)研究试样A横切面内早材管孔的分布特性。试验中通过生物图像软件Image-pro plus(IPP 6.0)计算试样A横切面内早材管孔带厚度和试样的厚度(图1),以及早材管孔总面积和试样的整个横切面面积[11]。然后根据SEM图像中早材管孔带的厚度和试样厚度计算试样A横切面内早材管孔带组织比量(RT)。此外,根据SEM图像中的早材管孔总面积和试样的整个横切面面积,计算试样A横切面内早材管孔组织比量(RC),并结合RT和RC研究试样A横切面内早材管孔的分布特性。

图1 生物图像软件Image-pro plus(IPP 6.0)计算示意图Fig. 1 Calculation schematic diagram of biographic software Image-pro plus(IPP 6.0)
1.3 试验方法
采用动态热机械分析仪(DMA-Q800型,美国TA Instruments公司)测试试样的蠕变变化特性。该分析仪配备的湿度附件专门用于通过调节干燥氮气和水饱和气流的比例,将试样环境控制在0%~90%相对湿度环境,温度范围为5~90 ℃,并选择跨距为35 mm的双悬臂夹具对试样径向加载恒定载荷并进行蠕变应变测试(图2)。测试温度分别为20,35,50,65和80 ℃,试样的相对湿度分别控制为67%,70%,74%,78%和81%,上述测试条件试样的对应平衡含水率均为(12±0.1)%。具体步骤:1)用电子天平(FA2004型,上海良平仪器仪表有限公司)称出试样质量,精度为0.000 1 g[12];2)将试样置于恒温恒湿箱(EK-10KA型,广州爱斯佩克环境仪器有限公司)中,并调节至相对应的温湿度环境进行调质,直至试样质量达到稳定值;3)将已调质好的试样取出并固定于双悬臂夹具中,为确保试样在试验过程中不发生弯曲,需施加0.01 N的预紧力;4)试样固定后关闭测试炉,开始进行测试,等待测试炉中温度和相对湿度达到预设的试验要求参数,保持该环境100 min以使试样含水率达到平衡含水率(12±0.1)%;5)当试样含水率达到平衡时,对试样施加5 MPa的应力,作用时间设置为90 min,在应力作用时间范围内进行应力应变扫描[6]。试样蠕变测试试验全程由DMA中的传感器采集数据并输入计算机中,最后生成相应的应变(百分比)随时间变化的数据图。
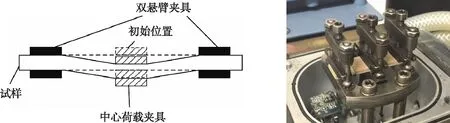
图2 DMA双悬臂夹具应变测试Fig. 2 Strain testing of DMA double cantilever bending
2 结果与分析
2.1 试样早材管孔带和管孔组织比量表征分析
试样A和B的横切面内早材管孔带分布特征见图3。由图3可知,在试样A的横切面内,早材管孔相对均匀地分布在早材管孔带中。本研究分别选取20幅具有代表性的整个横切面的SEM图像计算试样A和试样B的早材管孔带组织比量(RT)和早材管孔组织比量(RC)。通过计算可知,试样A的RT和RC分别为(32.41±1.59)%和(25.62±0.85)%;而试样B的RT和RC均为0。结果表明,试样A横切面内的早材管孔带组织比量和早材管孔组织比量基本保持一致。

图3 试样A和B横切面内早材管孔带分布Fig. 3 Earlywood vessel belt distribution in the cross section of specimens A and B
2.2 温度与环孔材栎木蠕变特性的关系分析
不同温度条件下试样A和B的应变随时间的变化曲线见图4。试验结果表明,试样A与B的应变均与温度的关系较大。由图4可知,在试验开始的45 min内,试样A和B的应变均随着温度的升高而逐渐增大。在温度为50~80 ℃范围内,试样A的应变增量随着温度的升高而显著增大,此时温度对其应变的影响更为明显;而试样B的应变从温度为50 ℃开始就随着温度的升高而显著增大,其应变增量随着温度的升高而显著增大。这些显著的变化主要是由于随着温度的升高(52.6~82.0 ℃),木材中的半纤维素[5,13]发生软化,特别是当测试温度为80 ℃时,由于湿木材的木质素玻璃化转变温度为72.0~128.0 ℃[14],环孔材栎木的木质素分子链段运动状况更为活跃;而且本试验持续时间为100 min,试验过程中栎木木质素发生一定程度的软化,宏观表现为木材蠕变应变增大。
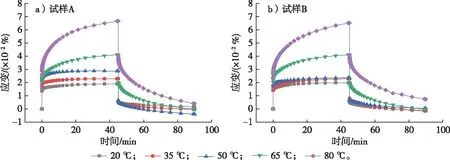
图4 不同温度下环孔材栎木试样蠕变曲线Fig. 4 Creep curves of ring-porous oak specimens at different temperatures
不同温度下试样A和B的瞬时应变和45 min应变参数见图5。由图5可知,试样A和B的瞬时应变和45 min应变均随温度升高而增大,并且其应变增量也随温度的升高而增大。当温度升高到80 ℃时,试样A的瞬时应变和45 min应变依次增加了85.3%和261.3%,应变增量显著增大,试样的蠕变现象较明显。当温度升高到80 ℃时,试样B的瞬时应变和45 min应变的增量依次为91.9%和249.2%,试样的蠕变现象较明显。此外,当温度从60 ℃升高到80 ℃时,试样A的瞬时应变由156.6% 升高到185.3%,增加了28.7%;其45 min应变由207.2%增加到361.3%,增加了154.1%。同等条件下试样B的瞬时应变和45 min应变分别增加了32.4%和133.5%。分析其原因:随着温度的升高,栎木中原本处于惰性的运动单元分子获得了足够的能量,使其由基态向激发态跃迁,环孔材栎木细胞壁内分子链发生伸展或者滑移,导致分子间化学键断裂,分子链的流动性和延展性增强,分子间距离增加,相互作用力减弱,运动单元分子活动范围增加[15-16]。此外,木材纤维素的热膨胀程度随着温度的升高而增大,纤维素分子间的内聚力降低,进一步促进了环孔材栎木中纤维素的软化,使试样宏观表现出的蠕变现象更加明显[16]。

图5 不同温度下环孔材栎木试样的瞬时应变与45 min应变Fig. 5 Instantaneous strain and 45 min strain of ring-porous oak specimens at different temperatures
2.3 早材管孔带与环孔材栎木蠕变特性的关系分析
试样A和B在20,35,50,65和80 ℃温度条件下的应变曲线见图6。由图6可知,相同的温度条件下,试样A的应变均大于试样B,说明早材管孔带的存在一定程度上增加了试样的应变。分析其原因,试样在应力作用下产生的弯曲变形与早材管孔带的存在和温度有关。根据弹性力学中孔或圆孔的应力集中效应可知,当具有小孔的弹性体受到外力作用时,有孔边的应力将远大于无孔时的应力[17],早材管孔带的存在有效地增大了试样的蠕变应变。然而,早材管孔带的影响随着温度的升高而先增大后减少。在20~50 ℃温度范围内,试样A与B的应变差值随着温度的升高而增大,并在50 ℃时达到最大值,对应的应变差值随着温度的继续升高(50~80 ℃)而逐渐减小。特别是当温度升高到80 ℃时,湿木材的木质素玻璃化转变温度为72.0~128.0 ℃[14],木质素分子链段的运动更活跃;而且本试验持续时间为100 min,试验过程中栎木木质素有一定程度的软化,发生一定程度的玻璃化转变,木材蠕变变形更加明显,温度对其蠕变的影响占据主导地位,早材管孔带对其蠕变形变的影响作用大幅度降低,试样A与B的应变曲线接近一致。

图6 不同温度和早材管孔分布下环孔材栎木蠕变曲线Fig. 6 Creep curves of ring-porous wood oak specimens at different distributions of earlywood vessel and temperatures
由图5可知,在前45 min内,试样A的瞬时应变和45 min应变均大于试样B,这是因为试样A中早材管孔带的存使得试样的蠕变应变增大。与试样B相比,试样A的瞬时应变和45 min应变增加率均随着温度的升高先增大后减小,瞬时应变和45 min应变增加率最大值对应的温度分别为35和50 ℃。当温度从20 ℃升高到35 ℃时,试样A相对于试样B的瞬时应变增加率从5.4%增大到22.7%,随着温度继续升高到80 ℃,对应的瞬时应变增加率降低到9.2%。此外,当温度从20 ℃升高到50 ℃时,试样A相对于试样B的45 min应变增加率从5.5%增大到19.6%,随着温度继续升高到80 ℃,对应的瞬时应变增加率降低到2.0%。这说明早材管孔带的存在对环孔材栎木试样瞬时应变和45 min应变的影响具有一定差异,其对瞬时应变的影响滞后于45 min应变的影响作用。
2.4 蠕变模型
木材蠕变作为一种典型的黏弹性行为,表现为在恒定外应力作用下,木材的应变随着作用时间的延长而逐渐增大。预测蠕变行为的必要性是毋庸置疑的,建立合适的数学模型对预测木材蠕变行为具有非常重要的意义[18-19]。
2.4.1 Burger模型
对于预测木材蠕变行为,国内外学者使用最多的是Burger模型(四参数模型),并且该模型对于模拟木材蠕变行为具有较好的效果。然而,研究发现Burger模型模拟预测的结果往往高于实测结果,主要是由于Burger模型通常用一个时间的线性函数关系来表征木材蠕变行为的黏性流动部分,但实际试验过程中木材蠕变行为的黏性流动部分是一个时间的非线性函数关系。而且木材蠕变变形的增长率随时间的推移而逐渐减小,使得这种影响在较低应力水平下更为明显[18]。
Burger模型的本构方程为:
(1)
式中:ε(t)为应变;t为时间;σ为应力;E0为普弹模量,反映理想弹性,即瞬时弹性的响应;E1为高弹模量;η1为黏弹性系数,反映滞后弹性形变;η2为本体黏度,反映撤除外力恢复过程完成后留下不可恢复的形变。
Burger模型的蠕变公式可简化为:
ε(t)=p1+p2(1-e-p3t)+p4t
(2)
式中:p1为瞬时弹性变形;p2为延迟弹性变形;p3为黏弹性系数;p4为黏性系数。
2.4.2 五参数模型
为了更好地预测模拟木材的蠕变特性,Dinwoodie等[15]提出了一个五参数模型,这个模型假设了一个非线性的黏弹性阻尼器,其模型的蠕变公式为:
ε(t)=β1+β2(1-e-β3t)+β4tβ5
(3)
式中:β1为瞬时弹性变形;β2为延迟弹性变形;β3为黏弹性系数;β4为黏性流动变形;β5为额外加入的一个无任何实际意义的参数,以表征黏性流动部分呈非线性增长的问题。
2.4.3 模型模拟结果分析
利用Burger模型和五参数模型拟合试样A和B的45 min蠕变应变曲线,其拟合结果如表1和2所示。由表1和2可知,五参数模型拟合的各元素决定系数(R2)为0.987 38~0.999 94,而Burger模型拟合的各元素决定系数(R2)为0.963 90~0.993 04。此外,不同温度下试样A和B的五参数模型和Burger模型拟合曲线与试验数据曲线的对比结果见图7,可以看出,两种模型的拟合曲线与试验测试所得曲线吻合程度较高,都可较好地模拟环孔材栎木试样的蠕变应变特性。总体而言,五参数模型可以更好地反映环孔材栎木的蠕变行为,其拟合程度整体上优于Burger模型。此外,相同试验条件下,五参数模型拟合结果的R2均大于Burger模型,说明由于非线性拟合的引入,五参数模型可以更好地预测模拟环孔材栎木的蠕变行为。
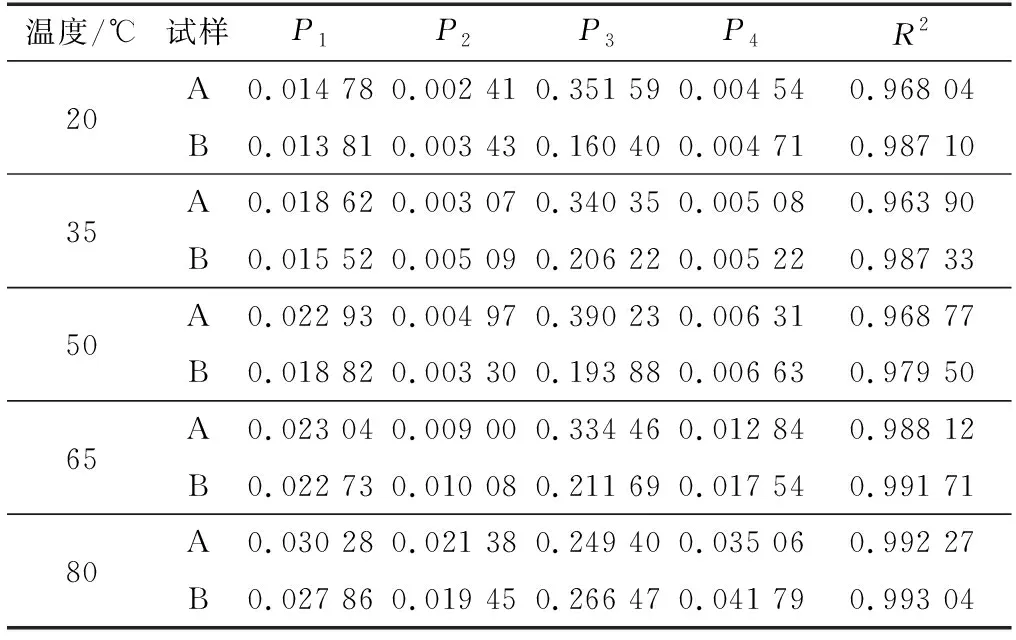
表1 Burger模型拟合结果Table 1 Fitting results of Burger model
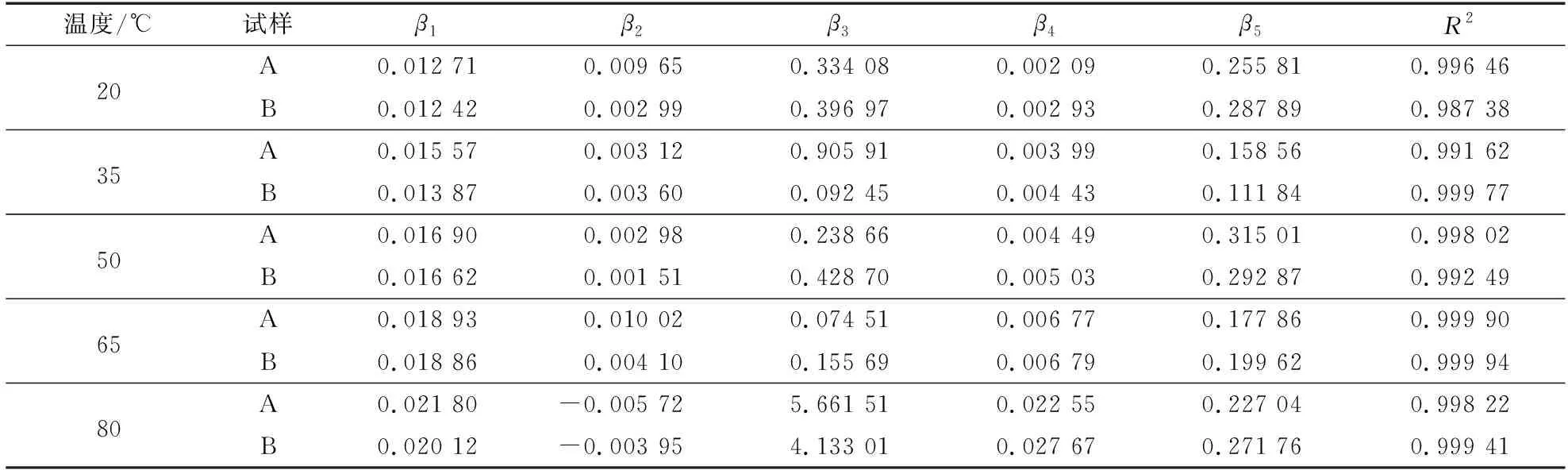
表2 五参数模型拟合结果Table 2 Fitting results of five-parameter model
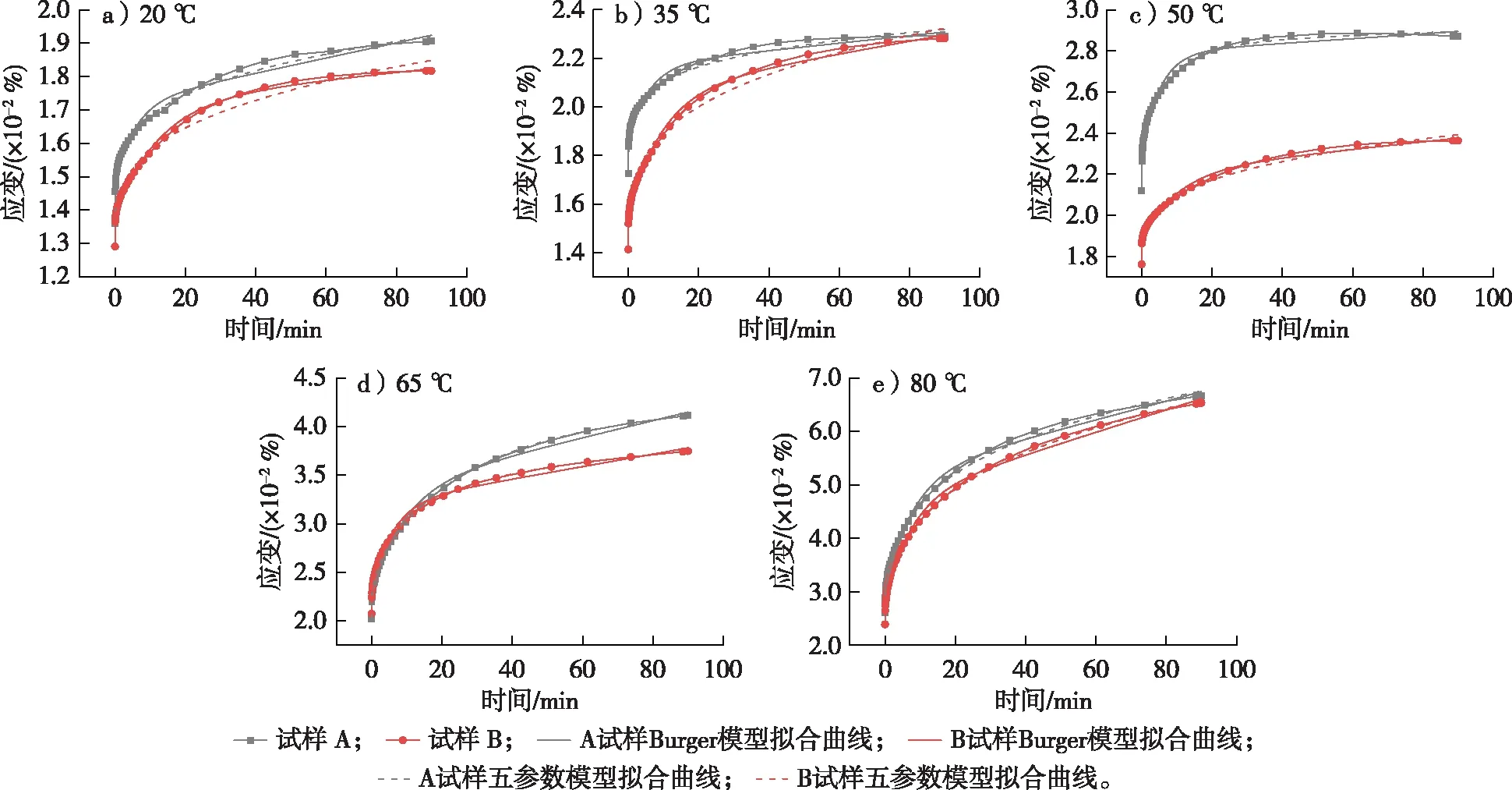
图7 不同温度下试样A和B的蠕变曲线和模型拟合曲线Fig. 7 Creep curves and model fitting curves of Specimens A and B at different temperatures
由表1和2所示的Burger模型和五参数模型拟合结果可知,试样A和B的瞬时弹性变形均随温度的升高而增加。这主要是由于木材在较小的外加恒定应力作用下,微纤丝及其细胞产生变形构成其瞬时弹性变形;随着温度的升高,木材细胞壁分子链发生伸展或者滑移,相邻分子间相互作用的化学键吸收足够的能量产生断裂,导致之前相互作用的运动单元活动空间范围增大[14-15],并且温度升高使得微纤丝发生热膨胀,导致纤维素分子间距离变小,使其分子间作用力减弱。当温度升高到木材半纤维素和木质素的玻璃化转变温度时,半纤维素(52.6~82.0 ℃)[5,13]和木质素(72.0~128.0 ℃)[14]发生玻璃化转变,试样表现为木材蠕变应变增长速率变快,相应的瞬时弹性变形程度也随温度的升高而增大,蠕变现象更加明显[5,20-21]。此外,温度的升高还使栎木试样更易于产生黏性转变,其黏性流动形变随温度升高而增大[6]。
由表1和2所示的Burger模型和五参数模型的拟合结果可知,在相同的试验条件下,试样A的瞬时弹性变形均大于试样B,主要是因为试样A中早材管孔带的存在一定程度上增强了试样在试验过程中的部分蠕变应变。此外,试样A的黏性流动形变均小于试样B。这是因为蠕变弯曲变形过程中试样A的导管细胞受到一定程度的拉伸,导管细胞的拉伸程度随着温度的升高而逐渐增大,温度过高导致导管细胞胞间层发生开裂[6],进而导致早材管孔对试样蠕变应变的增强效果减弱,并减小了试样A蠕变应变的黏性流动形变。
3 结 论
1)本试验条件下,试样A和B的应变均随着温度的升高而逐渐增大,而且试样A的应变均大于试样B。早材管孔带的存在对于环孔材栎木蠕变应变具有较大的影响,具体表现为试样A的瞬时应变和45 min应变均大于试样B。
2)早材管孔带对环孔材栎木蠕变应变的影响随着温度的升高先增大后减小。试样A与B的瞬时应变和45 min应变增加率均随着温度的升高先增大后减小,瞬时应变和45 min应变增加率最大值对应的温度分别为35和50 ℃。
3)Burger模型和五参数模型拟合数据与实测数据吻合,均可以较好地预测模拟环孔材栎木蠕变行为。此外,在相同试验条件下,五参数模型拟合结果的决定系数均大于 Burger模型,说明由于非线性拟合的引入,五参数模型可以更好地预测模拟环孔材栎木蠕变行为。
