表没食子儿茶素没食子酸酯的生理功能及其在畜禽生产中的应用
2021-05-20窦秀静吕文静管舒琪单安山
窦秀静 吕文静 管舒琪 单安山
(东北农业大学动物科学技术学院,哈尔滨 150030)
表没食子儿茶素没食子酸酯(epigallocatechin gallate,EGCG)是从绿茶中提取出来的一种天然的儿茶素类单体物质。近年来,多项研究表明EGCG具有抗氧化、抗炎、抗病毒、调节脂质代谢、提高机体免疫力等生理功能[5],EGCG被陆续应用于畜禽生产中,能够显著改善畜禽的生产性能,提高生物体的免疫调节及抗病能力。因此,为给当前畜牧业面临的饲用抗生素禁用带来的系列问题提供重要思路,本文就EGCG的生理功能及在畜禽生产中的应用进行综述,以期为其进一步开发和安全利用奠定重要基础。
1 EGCG的理化性质
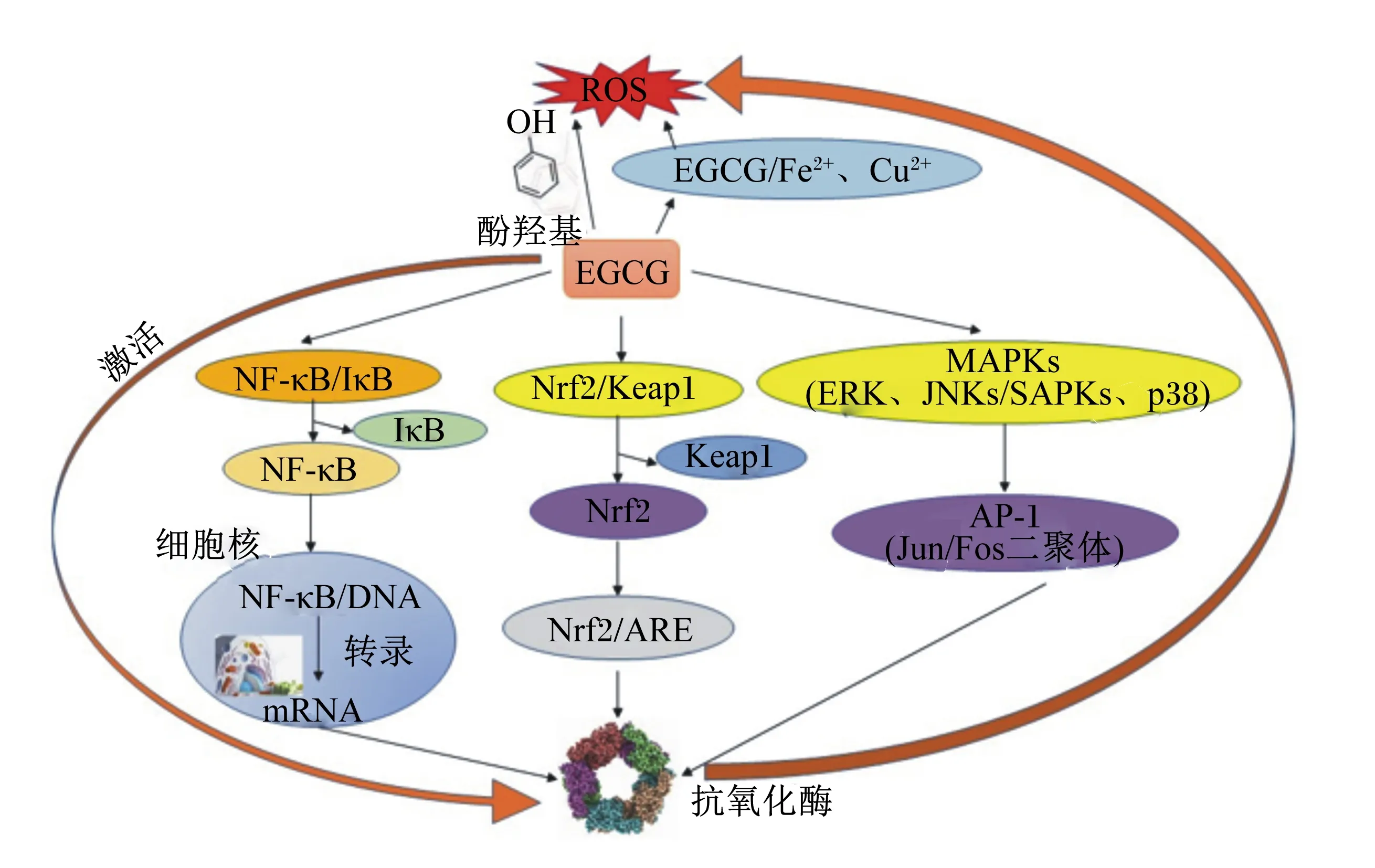
EGCG的分子式为C22H18O11,纯品EGCG是白色、粉红色或奶油色的无味晶体或粉末,可溶于水、甲醇、四氢呋喃、丙酮、吡啶和乙醇。EGCG对热、酸较稳定,在低浓度和低温条件下易被氧化,在高温(大于44 ℃)和酸性条件(2.0 生物体内不断进行着氧化反应,氧化代谢过程中通常伴随活性氧(reactive oxygen species,ROS)的产生。氧化应激是指当生物体内氧化与还原作用不平衡(倾向于氧化)时,机体内会产生多种氧化中间产物,这是自由基在生物体内产生的负面作用[10]。杨涪等]研究表明,EGCG的抗氧化能力分别是维生素E、维生素C的25、100倍。 2.1.1 EGCG利用酚羟基发挥抗氧化功能 酚羟基是EGCG发挥抗氧化作用的基础,能够提供活跃的氢来清除多种自由基。Nanjo等[11]试验结果表明,EGCG分子内含有的8个酚羟基可以清除15个1,1-二苯基-2-三硝基苯肼[1,1-diphenyl-2-picrylhydrazyl radical 2,2-diphenyl-1-(2,4,6-trinitrophenyl)hydrazyl,DPPH],即EGCG能够清除比自身酚羟基数更多的自由基。Frei等[12]研究表明,EGCG能与ROS反应生成稳定的酚氧ROS,从而清除ROS。 2.1.2 EGCG通过金属螯合作用发挥抗氧化功能 过渡金属如铁、铜等是许多自由基生成的催化剂,金属不平衡会导致机体内氧化应激的发生。EGCG由于自身的邻苯二酚结构,具有较强的金属离子螯合能力,能够抑制氧化应激的发生。Zhong等[13]发现,20 μg/mL的EGCG对亚铁离子的螯合能力约为8%。另外,EGCG对铜[14]、铬[15]和镉[16]也有螯合能力。Hider等[17]研究表明,茶多酚B环结构中的三羟基可能与金属螯合作用有关。Kumamoto等[18]研究显示,没食子酸酯基也是金属离子的结合位点。 2.1.3 EGCG通过调控细胞信号转导通路发挥抗氧化功能 核因子-κB(nuclear factor-κB,NF-κB)是重要的核转录因子,研究表明,ROS可以激活NF-κB,但是某些抗氧化剂会抑制NF-κB的活化[19]。环氧合酶和脂氧合酶会加剧机体内的氧化应激损伤,茶多酚可能是通过NF-κB途径抑制环氧合酶和脂氧合酶活性[20]。EGCG通过抑制NF-κB的活化以及下调下游的信号蛋白细胞外信号调节蛋白激酶(extra-cellular signal regulated kinase,ERK)和蛋白激酶B(protein kinase B,Akt)磷酸化,抑制环氧化酶-2(cyclooxygenas-2,COX-2)的mRNA及其蛋白的表达量上调[21]。c-Jun氨基末端激酶(c-Jun N-terminal kinase,JNK)是丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)家族中的重要组成部分,JNK信号通路是MAPK三大主要信号通路之一[22]。Cao等[23]研究发现,EGCG可以降低视网膜色素上皮ARPE19细胞中ROS的水平,增强JNK的磷酸化水平。核转录因子E2相关因子2(nuclear factor erythroid 2-related factor 2,Nrf2)/抗氧化反应元件(antioxidant response element,ARE)信号通路是目前发现的主要的机体内源性抗氧化应激通路[24]。机体在应对ROS损害时能产生多种保护蛋白,来缓解细胞遭受的损伤,这一过程是由1个DNA-启动子结合序列ARE调控的,Nrf2是ARE的激活因子[25]。研究表明,在EGCG的作用下,细胞内Nrf2及其下游血红素氧合酶-1(heme oxygenase-1,HO-1)蛋白表达均有所增加,二者的表达量与EGCG呈现剂量依赖性关系,进一步说明EGCG可以通过Nrf2/ARE通路抑制细胞的氧化应激反应[26-27]。Thangapandiyan等[28]研究表明,EGCG通过激活Nrf2/ARE信号通路,能抑制氟化物(fluoride,FI)中毒引起的睾丸Nrf2、HO-1、Kelch样环氧氯丙烷相关蛋白1(Kelch-like ECH-associated protein 1,Keap1)和醌氧化还原酶[NAD(P)H:quinone oxidoreductase,NQO1的mRNA的过表达,使FI中毒大鼠睾丸的Nrf2、Keap1、NQO1]和HO-1蛋白水平正常化,降低FI通过氧化应激引起的毒性。 2.1.4 EGCG通过激活体内抗氧化系统发挥抗氧化功能 综上所述,EGCG通过多种途径发挥抗氧化功能,其作用机制如图1所示。 ROS:活性氧reactive oxygen species;EGCG:表没食子儿茶素没食子酸酯epigallocatechin gallate;NF-κB:核因子-κB nuclear factor-κB;IκB:NF-κB抑制蛋白NF-κB inhibitor;Nrf2:核因子红细胞2相关因子2 nuclear factor erythroid 2-related factor 2;Keap1:Kelch样环氧氯丙烷相关蛋白1 Kelch-like ECH-associated protein-1;ARE:抗氧化反应元件antioxidant responsive element;MAPKs:丝裂原活化蛋白激酶mitogen-activated protein kinases;ERK:细胞外调节蛋白激酶extracellular regulated protein kinase;JNKs:c-Jun氨基末端激酶c-Jun N-terminal kinases;SAPKs:应激活化蛋白激酶stress activate protein kinases;p38:应激活化蛋白激酶;AP-1:激活蛋白-1 activator protein-1。 大量研究证实EGCG具有良好的抗炎功能,EGCG可减少炎症因子水平。但是,由于炎症过程的复杂性以及可能存在的干扰因子,EGCG发挥抗炎作用的机制尚未明确。 Nair等[34]研究表明,Toll样受体4(Toll like receptor 4,TLR4)在脂多糖(lipopolysaccharide,LPS)诱导的急性肾损伤(acute kidney injury,AKI)模型中表达增加,这可能与炎症因子介导的受体表达增加有关,引起机体炎症反应。李木子等[35]研究发现,EGCG在LPS诱导的AKI模型中可以抑制TLR4及其下游因子髓样分化因子(myeloid differentiation factor 88,MyD88)和NF-κB的表达,说明EGCG发挥抗炎功能可能与TLR4/MyD88/NF-κB通路有关。Xie等[36]试验表明,在体外试验中,人脐静脉内皮细胞(human umbilical vein endothelial cells,HUVEC)在经过EGCG干预后,NF-кB的下游炎症因子单核细胞趋化蛋白-1(monocyte chemotactic protein-1,MCP-1)、细胞间黏附分子-1(intercellular cell adhesion molecule-1,ICAM-1)、肿瘤坏死因子-α(tumour necrosis factor-α,TNF-α)表达均显著降低。在体内试验中,该试验结果同样适用[37]。在赵悦伶等[38]研究中,试验人员利用葡聚糖硫酸钠(dextran sulphate sodium,DSS)建立小鼠肠道炎症模型,测得EGCG(0.5%)可以减少白细胞介素-6(interleukin-6,IL-6)和TNF-α的表达,说明EGCG可减少炎症因子的表达,保护患IBD的小鼠。另外,EGCG可通过降低炎症因子IL-6、TNF-α、白细胞介素-1β(interleukin-1β,IL-1β)的表达来保护大鼠肾脏[35],EGCG与姜黄素发挥抗炎作用的机制不同,Fan等[39]研究表明,姜黄素可上调衔接因子蛋白含PH域蛋白1(adaptor protein,phosphotyrosine interaction,PH domain and leucine zipper containing 1,APPL1)的表达量,抑制Akt磷酸化,保护缺血再灌注引起的急性肾损伤。NLRP3炎症小体能够促进促炎症细胞因子释放,Lu等[40]发现姜黄素可抑制NLRP3炎症小体活性,改善糖尿病肾病。 Ge等[41]发现,EGCG能够有效抑制猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)在猪肺泡巨噬细胞(porcine alveolar macrophages,PAMs)中的感染和复制,降低PAMs细胞的毒性。同样是PRRSV病毒,芦荟多糖可阻止病毒颗粒对靶细胞的吸附,这可能是通过提高细胞膜的稳定性实现的[42]。EGCG同样能够防止病毒粒子吸附在细胞上,Li等[43]研究表明,EGCG可以直接靶向猪圆环病毒2型(porcine circovirus type 2,PCV2)病毒粒子,防止病毒粒子附着在宿主细胞上。Noémie等[44]研究表明,EGCG能够直接作用于丙型肝炎病毒(hepatitis C virus,HCV),阻止病毒直接接触细胞表面,抑制病毒在细胞间的传播。曾闵等[45]研究发现,EGCG可以降低血清中心肌肌钙蛋白T(cardiac troponin T,cTnT)、肌酸激酶同工酶MB(creatine kinase-MB,CK-MB)的活性,保护患病毒性心肌炎的小鼠。另外,EGCG棕榈酸单酯(由EGCG与棕榈酰氯的酯化制备获得)可以抑制人和家禽类流感病毒以及抗药性病毒,比EGCG的抗流感病毒能力高24倍,而且在鸡产蛋时期对病毒的抑制效果更好[46]。 在畜禽生产过程中,母畜妊娠期或肉种鸡(或蛋鸡)产蛋高峰后期,容易发生脂肪的沉积,且以腹脂沉积最多,从而降低畜禽的生产性能[47]。Friedrich等[48]发现,服用EGCG的小鼠体重减轻的可能原因是EGCG降低了体内的热量,并减少了常量营养素的吸收。Forester等[49]用EGCG喂养小鼠发现,EGCG通过抑制机体内淀粉酶的活性降低小鼠体内血糖浓度。陈粉粉等[50]发现,200 μmol/L的EGCG能够抑制猪前体脂细胞的繁殖生长,同时EGCG通过降低脂肪细胞分化转录因子过氧化物酶体增殖物激活受体γ2(peroxisome proliferator-activated receptor γ2,PPARγ2)和CCAAT增强子结合蛋白α(CCAAT/ enhance binding protein α,C/EBPα)mRNA的表达量减少脂肪细胞的分化。Chen等[51]研究表明,EGCG抗肥胖的作用机制之一是其能在白色脂肪细胞上诱导与棕色脂肪细胞相关的基因表达。何帅等[52]试验结果显示,EGCG能预防高脂饮食大鼠的体重升高,能明显降低甘油三酯(triglyceride,TG)和低密度脂蛋白(low density lipoprotein,LDL)含量,升高高密度脂蛋白(high density lipoprotein,HDL)含量,改善脂代谢紊乱。EGCG与葡萄籽缩合单宁、女贞子提取物作用途径不同,葡萄籽缩合单宁可降低脂肪酸合成酶基因的表达量,抑制肝脏中脂肪的形成[53]。女贞子中红景天苷可下调磷酸烯醇式丙酮酸激酶(phosphoenolpyruvate kinase,PEPCK)和葡萄糖-6-磷酸酶(glucose-6-phosphatase,G6Pase)的表达,减少小鼠体内脂质的含量[54]。 Yi等[55]研究发现,EGCG可提高经6 Gy60Co γ射线照射小鼠的肝脏指数、脾脏指数和胸腺指数,提高小鼠的脾细胞转化率和单核细胞吞噬指数,预防辐照引起的免疫系统损伤。巨噬细胞在免疫应答中发挥重要作用,Lucia等[56]发现,EGCG能增加脊髓损伤(spinal cord injury,SCI)中M2型巨噬细胞数目,降低M1型巨噬细胞数目。Huang等[57]以白血病小鼠为模型发现,EGCG能够增加白血病小鼠体内巨噬细胞的数目,提高巨噬细胞的吞噬能力。Matsunag等[58]对感染了嗜肺性军团病杆菌的巨噬细胞进行体外培养,发现EGCG能够增加巨噬细胞内TNF-α、干扰素-γ(interferon-γ,IFN-γ)和白细胞介素-12(interleukin-12,IL-12)的分泌,提高巨噬细胞的免疫能力,降低肿瘤发生的可能性。EGCG与生姜提取物发挥促免疫作用的机制类似,Li等[59]研究表明,生姜提取物可提高淋巴细胞活性,活化巨噬细胞,促进免疫球蛋白的产生。另外,生姜提取物可提高巨噬细胞迁移指数,增强机体的体液免疫功能[60]。以上研究表明,EGCG可提高机体免疫器官指数,增加免疫细胞数目,提高免疫细胞活性和分泌能力,进而提高机体免疫力。 随着动物产业的迅速发展和人们对动物产品需求的提高,EGCG因具有较好的抗氧化、抗炎、抗病毒、调节脂质代谢、提高机体免疫力等作用而被作为一种新型高效饲料添加剂应用于动物生产中,EGCG可以提高动物的生产性能、免疫能力和畜产品品质,展现出良好的应用前景。 EGCG中含有的酚羟基可以和蛋白质分子中的氨基或羟基反应,防止细菌侵入,起到杀菌功能;同时EGCG含有的酚羟基还能和细菌内的二氢叶酸合成酶结合,抑制细菌的增殖,改善猪的生长环境[61]。将500或1 000 mg/kg的茶多酚添加到氧化应激仔猪的饲粮中,能够有效提高猪的生长性能和免疫能力,但500 mg/kg的作用效果不如1 000 mg/kg的作用效果[62]。姚波等[63]发现把茶多酚添加到育肥料中能提高猪的平均日增重,降低饲料转换率,且添加量为450 g/t组效果优于650 g/t组。徐瑞等[64]研究表明,在饲粮中添加2%的茶叶渣对肥育猪饲粮利用效率以及肉质有显著的改善作用。贾娜等[65]发现,将儿茶素添加到猪肉糜中能够降低猪肉糜的弹性和硬度,提高猪肉糜的色泽和质量。综上可知,EGCG能够改善猪的生长环境,提高仔猪的免疫能力,改善育肥猪肉品质。 Sahin等[66]研究表明,在鹌鹑饲粮中添加200或400 mg/kg EGCG,能够降低肝脏中脂肪的氧化程度,提高抗氧化酶和解毒酶的活性,且作用效果与剂量呈依赖性关系。黄莉等[67]研究发现,EGCG能提高抗氧化物酶GSH-Px、SOD和CAT的活性,降低肉鸡血清中的MDA含量,提高肉鸡的抗氧化能力和生长性能;另外,EGCG能够显著促进肉鸡胸肌谷胱甘肽过氧化物酶4(glutathione peroxidase 4,GPx4)基因mRNA的表达,提高GSH-Px活性,使肉鸡货架期延长。曾亮[68]研究表明,将儿茶素添加到鸭腿肉和鸭胸肉中可以提高鸭腿肉和鸭胸肉的T-AOC和色泽,降低鸭胸肉和鸭腿肉的MDA含量,从而增加鸭肉的保鲜时间。郜卫华[69]研究表明,饲粮中添加20、40、80 mg/kg的茶多酚能够使肉母鸡的淋巴细胞转化率分别提高3.2%、11.7%和11.0%,使肉公鸡的淋巴细胞转化率分别提高12.1%、17.7%和21.0%,提高肉鸡免疫力。刘海军[70]研究表明,将30 mg/kg茶多酚添加到肉鸡饲粮中,肉鸡的法氏囊指数、胸腺指数、脾脏指数分别提高了15.38%、15.75%、6.74%,说明茶多酚可有效提高肉鸡的抗病能力。以上研究表明,饲粮中添加EGCG能够改善肉鸡的抗氧化状态,提高肉鸡的免疫能力,改善肉产品品质。 反刍动物机体内氧化应激的发生会引起多种病症,比如乳房症、腹水症等。在反刍动物的饲粮中添加适量的茶多酚,能够抑制乳酸脱氢酶(lactate dehydrogenase,LDH)活性,提高SOD活性,降低反刍动物乳腺上皮细胞中的MDA含量,显著缓解氧化应激对反刍动物的损害,有效提升反刍动物免疫力[71]。王振云等[72]发现茶多酚能够抑制LDH和半胱氨酸蛋白酶3的活性,使乳腺上皮细胞清除自由基的能力提高,抑制脂质过氧化反应,保护受氧化应激损害的奶牛乳腺上皮细胞。Zhong等[73]研究表明,在山羊饲粮中添加儿茶素能够增加细胞膜的稳定性,减少脂质氧化和鲜肉的滴水损失,改善鲜肉的色泽。 茶多酚在其他动物生产中也表现出良好功效,具有较好的市场前景。防御素是抗菌肽的一类重要分支,具有良好的杀菌功能[74]。Bedran等[75]研究表明,茶黄素(表儿茶素没食子酸酯)可以提高口腔上皮细胞分泌抗菌肽人源β-防御素1(human β-defensin 1,hBD1)、抗菌肽人源β-防御素2(human β-defensin 2,hBD2)和抗菌肽人源β-防御素4(human β-defensin 4,hBD4)的能力。绿茶提取物和EGCG通过诱导上皮细胞分泌hBD,保护hBDs免受齿龈磷蛋白酶降解,提高上皮抑菌能力[76]。将0.05%的茶多酚添加到水貂饲粮中,能够显著提升水貂对钙、磷的消化率和平均日增重[77]。 近些年来,EGCG由于具有多种对生物有益的功能而备受关注,EGCG可以抑制细胞的氧化应激,对炎症和病毒也具有显著的抑制作用,还可以调节脂质代谢并提高机体免疫力。但是,EGCG发挥各种生理功能的分子机制,仍有待深入研究。 EGCG的结构不稳定,这导致它在到达作用靶点发挥作用前就很容易被氧化或转化成其他结构形式,从而失去其生理性能。目前,许多输送系统都是为了保护EGCG的结构完整,防止它退化[78-80]。酚羟基修饰法是对EGCG进行结构修饰的主要手段,酚羟基修饰主要可分为糖苷化修饰、甲基化修饰、酰基化修饰3种[81],此外,EGCG的衍生物合成方法还有苯环H原子取代法、C环羟基酯化法。 综上所述,对EGCG进行保护或结构改造,使其可以作为饲料添加剂发挥应有的作用,可以作为新的研究方向。目前,EGCG在畜禽生产中的应用还不够广泛,对于不同动物品种以及同种动物不同生理阶段下的添加量、添加方式仍然需要进一步研究确定。2 EGCG的生理功能
2.1 抗氧化功能

2.2 抗炎功能
2.3 抗病毒功能
2.4 调节脂质代谢功能
2.5 提高机体免疫力功能
3 EGCG在畜禽生产中的应用
3.1 EGCG在猪生产中的应用
3.2 EGCG在禽类生产中的应用
3.3 EGCG在反刍动物生产中的应用
4 小 结
