蜡蝉次目(昆虫纲:半翅目)昆虫化石研究进展
2021-05-14姚云志
张 晓,姚云志,任 东,庞 虹
(1. 中山大学生命科学学院/生态学院,中山大学生物博物馆,有害生物控制与资源利用国家重点实验室,广州 510275;2. 首都师范大学生命科学学院,昆虫演化与环境变迁重点实验室,北京 100048)
蜡蝉次目Fulgoromorpha 隶属于昆虫纲Insecta半翅目Hemiptera头喙亚目Auchenorrhyncha,是一类种类繁多的植食性昆虫,已知现生类群12 635种(Bartlettetal., 2018),广泛分布于世界各地。它们以刺吸式口器吸食植物汁液,很多种类是重要的农林业害虫,危害玉米、小麦、水稻、葡萄、甘蔗等。部分蜡蝉具有强烈延伸的头部、明显的雌雄异形、能够分泌蜜露、产生蜡质等特点(Urban and Cryan, 2007)。蜡蝉次目目前包括3个总科,其中两个为灭绝总科,这一类群的系统位置以及次目下总科级和科级阶元的系统关系一直存在着较大争议(Urban and Cryan, 2007; Song and Liang, 2013)。
蜡蝉次目昆虫出现时间较早,最早的化石记录发现于晚二叠世(Carpenter, 1992; Shcherbakov and Popov, 2002)。本文对世界蜡蝉次目化石昆虫研究情况加以总结,并对存在问题和研究前景进行分析。
1 蜡蝉次目化石昆虫研究简史
1.1 世界蜡蝉次目化石昆虫研究简史
世界蜡蝉次目化石昆虫研究已近200年,已记载蜡蝉次目化石昆虫28科、216属、407种,其中主要化石产地分布于德国、波罗的海、美国、英国、俄罗斯、巴西、缅甸以及中国。根据其化石研究历史可以分为以下几个时期:
蜡蝉化石研究的初始期(从19世纪初期到末期):1825年,Dalman首次从更新世东非柯巴脂中报道广翅蜡蝉科Ricaniidae 1属1种——马广翅蜡蝉Ricaniaequestris;此后,Brodie(1845)报道了一个来自早白垩世英国的蜡蝉化石;Germar and Berendt(1856)从始新世波罗的海琥珀中发现11个新类群;同时,Giebel(1856, 1862)分别对渐新世法国和更新世东非柯巴脂中的蜡蝉进行了研究;之后,Scudder(1867, 1877, 1878, 1890, 1895)记录了新生代美国和加拿大的近22个化石类群;Geinitz(1880, 1884)发现了德国早侏罗世时期的类蜡蝉科Fulgoridiidae化石标本。这个时期研究人员和所报道的化石记录均较少,研究工作仅停留在分类描述上。
蜡蝉次目化石昆虫研究快速发展期(20世纪初期到中期):Handlirsch(1906, 1921, 1939)、Bode(1905)、Martynov(1926, 1937, 1939)报道了来自侏罗纪欧洲和亚洲的类蜡蝉科77种;同时,Martynov(1935)发现了晚二叠世俄罗斯地区古老的蜡蝉化石;Cockerell(1909, 1910, 1917, 1920, 1924, 1926)对来自英国、美国、俄罗斯、阿根廷蜡蝉化石进行了大量研究,包括波罗的海琥珀及缅甸琥珀中发现的类群。此外,相关研究学者Henriksen(1922)、Tillyard(1923)、Piton(1940)等报道了新生代产自丹麦、法国和澳大利亚的蜡蝉化石6种。这一时期的研究仍以经典分类为主。
蜡蝉次目化石昆虫研究深入发展期(20世纪中期到末期):这期间有大量新类群被报道。Bode(1953)补充了早侏罗世德国类蜡蝉科化石58个新类群;Becker-Migdisova(1955, 1960, 1961)对俄罗斯地区二叠纪和三叠纪早期蜡蝉进行了研究;Hamilton(1990)从早白垩世巴西发现的拉蜡蝉科Lalacidae和颖蜡蝉科Achilidae化石记录,共报道24种;Haupt(1956)、Fennah(1961,1968,1987)、Hong(1979)、Emeljanov(1983)等研究学者对德国、英国、美国、黎巴嫩、中国、俄罗斯多地蜡蝉次目化石昆虫进行了深入研究。
进入21世纪后,蜡蝉次目化石研究进入繁荣阶段。Szwedo and Stroiński(2001, 2010, 2017)、Bourgoin and Lefèbvre(2002)、Emeljanov(2002, 2008)、Shcherbakov(2006, 2007, 2017)、Zhangetal.(2017, 2019)、Jiangetal.(2018, 2019)、Fuetal.(2019)、Songetal.(2019)等一大批学者对蜡蝉次目化石昆虫展开研究。其中波兰古昆虫学家Szwedo的贡献最为突出,共报道世界蜡蝉化石新记录86种,在2004年完成著作 《Fossil Planthopper (Hemiptera: Fulgoromorpha)of the World》,对整个蜡蝉次目昆虫化石进行了全面系统的总结、修订,是目前研究中重要的参考资料。值得一提的是,这一时期从多米尼加琥珀、波罗的海琥珀、乌克兰琥珀、缅甸琥珀、黎巴嫩琥珀、墨西哥琥珀、德国比特菲尔德琥珀、法国瓦兹琥珀以及东非柯巴脂中发现了大量的蜡蝉化石(Bourgoin and Lefèbvre, 2002; Stroiński and Szwedo, 2002; Solórzano Kraemer and Petrulevicius, 2007; Szwedo, 2007; Emeljanov and Shcherbakov, 2009, 2011; Szwedo, 2011; Zhangetal., 2019; Emeljanov and Shcherbakov, 2020),这些琥珀非常完好的保存了虫体三维立体结构,为经典分类、系统发育、生物地理、功能形态、生物学习性等方面的研究提供了非常好的研究材料,同时也解答了蜡蝉次目部分演化上的难题。
1.2 中国蜡蝉次目化石昆虫研究简史
我国蜡蝉次目化石昆虫研究起步较晚,洪友崇先生于1979年首次从中新世山东山旺组报道蜡蝉科Fulgoridae化石1属1种,开启了我国蜡蝉化石研究的篇章;之后,林启斌、张俊峰、任东、张海春、李姝等研究学者继续此项工作的研究。近年来国外古昆虫学家Szwedo和Stroiński(2011, 2013, 2015)也参与了我国蜡蝉次目化石的研究。截止目前为止,我国共报道蜡蝉次目化石昆虫 8科 14属18种。化石产地广布于中国11个地区:青海、西藏、山东、内蒙古、广西、新疆、北京、湖南、云南、甘肃、辽宁。化石产出时代从早侏罗世到中新世均有发现。
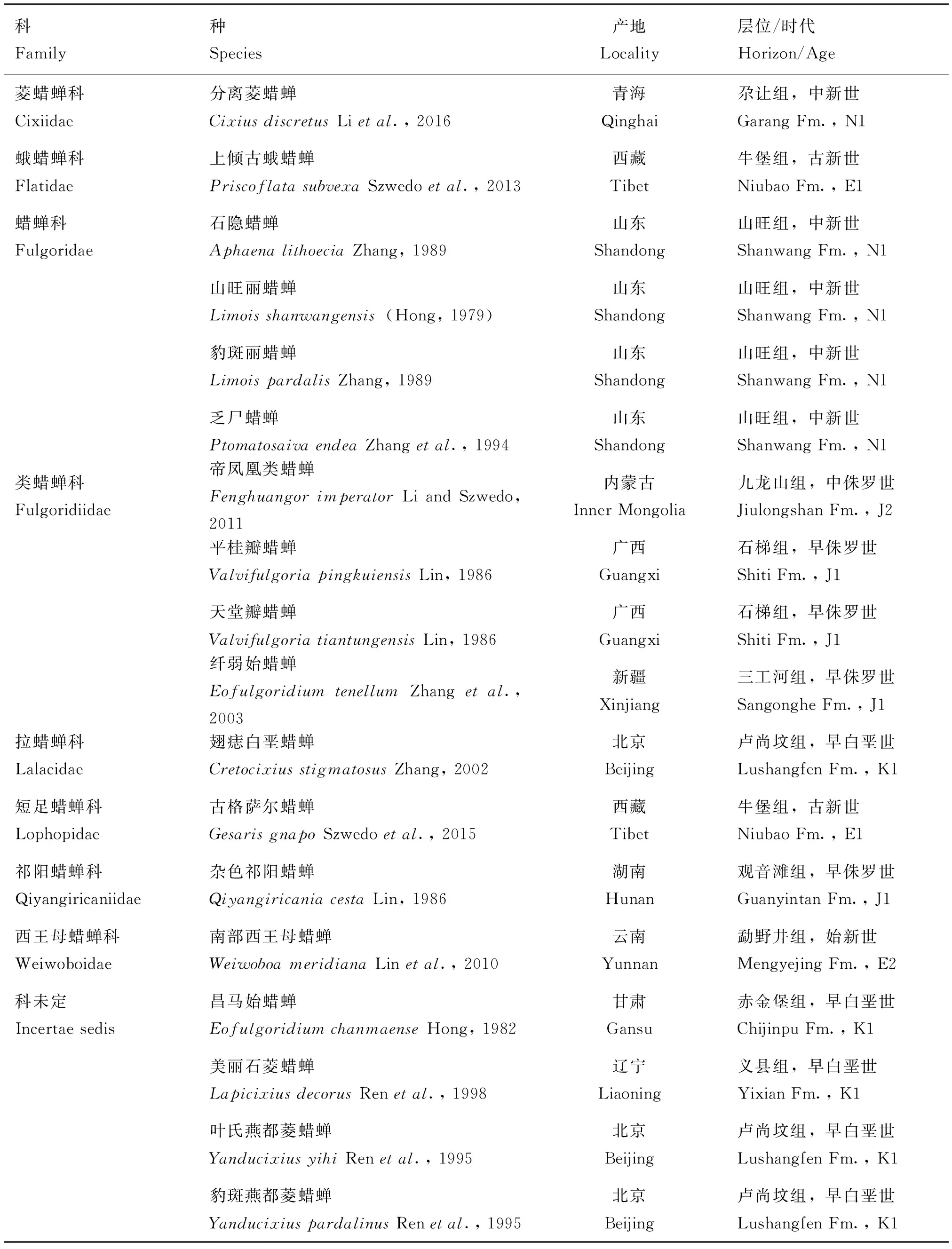
表1 中国蜡蝉次目化石名录
2 蜡蝉次目化石昆虫地质历史
古革蝉科Archescytinidae是公认的头喙亚目的祖先类群,体型较小,大约只有5 mm左右,在二叠纪中期和晚期空前繁盛。蜡蝉次目是头喙亚目中一类古老的类群,有着漫长的地质历史,最早的化石记录产自晚二叠世罗德阶(273~269 Ma)俄罗斯阿尔汉格尔斯克和克麦罗沃地区,属于革蜡蝉科Coleoscytidae(Shcherbakov and Popov, 2002;Brysz and Szwedo, 2019)。蜡蝉次目下共有两个灭绝总科革蜡蝉总科Coleoscytoidea和苏菱蜡蝉总科Surijokocixioidea,和一个现生总科蜡蝉总科Fulgoroidea(Wangetal., 2019)。已知蜡蝉总科的化石数量最大,物种最为丰富,共涉及11个灭绝科和15个现生科(Brysz and Szwedo, 2019)。
2.1 古生代
二叠纪蜡蝉次目化石记录较少,仅有8属16种被报道,它们分属于两个灭绝的总科革蜡蝉总科和苏菱蜡蝉总科,分布范围局限于俄罗斯地区(Szwedoetal., 2004)。
2.2 中生代
三叠纪蜡蝉次目的化石极为稀少,仅苏菱蜡蝉总科下的苏菱蜡蝉科Surijokocixiidae有化石记录,发现3属3种,来自三叠纪中期和晚期的澳大利亚昆士兰地区(Evans, 1971; Lambkin, 2020);此外,另有一种(科级地位未定)分布于三叠纪中期的德国(Brauckmann and Schlüter, 1993)。
侏罗纪时期,蜡蝉次目昆虫物种丰富度显著增高。蜡蝉总科中古老的祖先类群开始出现,类蜡蝉科在侏罗纪极为繁盛,已发现16属140种,大多发现于早侏罗世的德国,侏罗纪大部分蜡蝉化石均被归于类蜡蝉科,这个科一直被认为是并系,需要全面修订(Szwedoetal., 2011)。一些类蜡蝉具有极其长的喙,延伸到达腹部末端,可能表明早期蜡蝉通常和木本植物关系密切(Shcherbakov, 2002);许多类蜡蝉还具有明显的色斑,比如眼斑,这种迷惑性色斑被认为具有分散捕食者注意力的作用(Shcherbakov, 2002)。同时期存在的还有翅型和翅脉均和高等蜡蝉“higher Fulgoroidea”非常相似的祁阳蜡蝉科Qiyangiricaniidae,目前仅在早侏罗世中国湖南发现1属1种(Szwedoetal., 2011)。
到白垩纪时期,侏罗纪的代表类群类蜡蝉科和祁阳蜡蝉科已经绝灭,同时出现了许多仅存在于白垩纪的灭绝科。白垩纪早期蜡蝉次目化石昆虫种类较为丰富,新出现的类群有青蜡蝉科Neazoniidae(仅报道了若虫)(Szwedo, 2007);类似于菱蜡蝉Cixiidae的拉蜡蝉科Lalacidae(Hamilton, 1990),覆翅前缘强烈增厚;成虫保留有感觉窝的孔瓢蜡蝉科Perforissidae、拟蛛蜡蝉科Mimarachnidae(Zhangetal., 2017, 2018);以及现生菱蜡蝉科和颖蜡蝉科的最早记录(Fennah, 1987; Martins-Neto, 1988; Hamilton, 1990),其中青蜡蝉科和拉蜡蝉科仅发现于早白垩世,孔瓢蜡蝉科和拟蛛蜡蝉科的代表一直延续到晚白垩世。白垩纪中期蜡蝉次目昆虫在缅甸琥珀中被大量发现,头部强烈延伸的矛蜡蝉科Dorytocidae(Emeljanov and Shcherbakov, 2018),全身被覆浓密长毛,隐藏生活的鬃蜡蝉科Jubisentidae(Zhangetal., 2019),基室三角形的神蜡蝉科Yetkhatidae(Songetal., 2019),翅脉多分支的密脉蜡蝉科Katlasidae(Luoetal., 2020)均是缅甸琥珀特有类群。白垩纪晚期,象蜡蝉科Dictyopharidae+蜡蝉科支系最古老的成员在俄罗斯泰梅尔琥珀中被发现(Emeljanov, 1983; Shcherbakov, 2004)。由此可见,白垩纪蜡蝉类群明显发生了改变,这些新兴类群多为过渡类群,到白垩纪末期基本消失,只有现生菱蜡蝉科、颖蜡蝉科和象蜡蝉科的代表延续至新生代。
2.3 新生代
新生代所发现的蜡蝉次目化石绝大部分可以归入到现生科,仅有一灭绝科——西王母蜡蝉Weiwoboidae发现于始新世早期的中国云南(Linetal., 2010)。
古近纪是蜡蝉次目昆虫繁盛的时期。白垩纪-古近纪危机之后,古新世蜡蝉次目化石数量明显减少,此时短足蜡蝉科Lophopidae最早的化石记录在欧亚大陆出现(Stroiński and Szwedo, 2012; Szwedoetal., 2015),同时高等蜡蝉“higher Fulgoroidea”(蜡蝉总科内非正式的分组,包括该类群中一些较为进化的科)也从古新世开始出现,其中蛾蜡蝉科Flatidae、脊唇蜡蝉科Nogodinidae和瓢蜡蝉科Issidae,分别发现于中国、美国、法国、阿根廷等地(Piton, 1940; Fennah, 1968; Petrulevicius, 2005; Szwedoetal., 2013; Bourgoinetal., 2020)。进入始新世之后,蜡蝉次目昆虫发生了大规模辐射演化,近14个科的100余种被发现,主要分布于波罗的海琥珀,其中现生飞虱科Delphacidae、袖蜡蝉科Derbidae、颜蜡蝉科Eurybrachidae、蜡蝉科、广翅蜡蝉科、扁蜡蝉科Tropiduchidae均从始新世开始出现。此外,颖蜡蝉科、菱蜡蝉科、脊唇蜡蝉科在始新世属种发生了明显分化。
新近纪蜡蝉次目化石昆虫记录只发现于中新世。此时期报道的化石丰富度较低,短翅蜡蝉科Caliscelidae和阉蜡蝉科Kinnaridae均首次发现于多米尼加琥珀中(Emeljanov and Shcherbakov, 2000; Bourgoinetal., 2015),至此,现生蜡蝉次目科级格局基本形成。
第四纪蜡蝉次目昆虫化石发现较少,只有零星袖蜡蝉科和广翅蜡蝉科报道于更新世东非柯巴脂中(Synave, 1973; Stroiński and Szwedo, 2002)。

图1 蜡蝉次目化石昆虫代表Fig.1 Fossil representatives of Fulgoromorpha注:A, 中新世菱蜡蝉科, 分离菱蜡蝉; B, 中白垩世孔瓢蜡蝉科, 异形窝孔瓢蜡蝉; C, 中白垩世鬃蜡蝉科, 胡氏鬃蜡蝉; D, 中侏罗世类蜡蝉科, 帝凤凰类蜡蝉; E, 中白垩世拟蛛蜡蝉科, 无毛延拟蛛蜡蝉。比例尺A=B=C=1 mm; D=E=5 mm。Note: A, Miocene Cixiidae, Cixius discretus Li et al., 2016; B, Mid-Cretaceous Perforissidae, Foveopsis heteroidea Zhang et al., 2017; C, Mid-Cretaceous Jubisentidae, Jubisentis hui Zhang et al., 2019; D, Middle Jurassic Fulgoridiidae, Fenghuangor imperator Li and Szwedo, 2011; E, Mid-Cretaceous Mimarachnidae, Jaculistilus oligotrichus Zhang et al., 2018. Scale bars A=B=C=1 mm; D=E=5 mm.
3 世界蜡蝉次目化石昆虫地理分布和年代
世界已知的蜡蝉次目昆虫化石广布于23个国家:俄罗斯、德国、澳大利亚、吉尔吉斯斯坦、中国、英国、哈萨克斯坦、巴西、黎巴嫩、西班牙、日本、法国、蒙古国、缅甸、阿根廷、美国、波罗的海、乌克兰、丹麦、加拿大、多米尼加、墨西哥、坦桑尼亚。其中德国发现化石种类最为丰富,共145种,占已发现总数的近三分之一。
根据化石记录情况,二叠纪蜡蝉次目昆虫均发现于俄罗斯地区,推测该地区可能为蜡蝉次目昆虫重要起源中心;三叠纪蜡蝉次目种类急剧下降可能和二叠纪末期的大灭绝事件有关,分布范围开始扩大到南半球冈瓦纳大陆;侏罗纪早期蜡蝉次目昆虫多样性迅速增长,分布地向亚洲东部扩散;白垩纪时期蜡蝉次目昆虫科级单元快速分异,属种多样性明显增加,化石产地增多,分布范围进一步扩散到南北美洲和非洲;第五次生物大灭绝事件之后,蜡蝉次目化石昆虫种类明显减少,在始新世,蜡蝉次目经历了一次快速的辐射演化,许多现生科的最早化石记录发现于这一时期,这一时期化石集中分布于欧亚大陆和美洲大陆;新近纪之后蜡蝉次目昆虫逐渐演化成现今全球分布格局。
4 世界蜡蝉次目化石昆虫属种组成特征
当前已记载的蜡蝉次目化石昆虫共涉及28科、216属、407种。其中类蜡蝉科种类最多,共16属140种,占全部已描述命名类群总数的34.6%;其次是菱蜡蝉科,29属43种,占已描述命名总数的10.6%;之后是脊唇蜡蝉科21属33种,占8.1%;拉蜡蝉科9属17种,占4.2%;扁蜡蝉科15属17种,占4.2%;颖蜡蝉科12属16种,占4.0%;拟蛛蜡蝉科11属15种,占3.7%;蜡蝉科9属14种,占3.5%;除此之外,其他科化石记录只有零星报道。
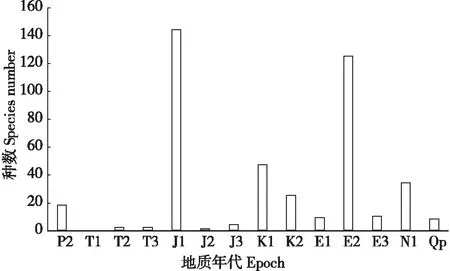
图2 世界蜡蝉次目化石各地质时期种级分异度Fig.2 The species number of the world’s Fulgoromorpha fossils in different geological ages注:P2,晚二叠世; T1,早三叠世; T2,中三叠世; T3,晚三叠世; J1,早侏罗世; J2,中侏罗世; J3,晚侏罗世; K1,早白垩世; K2,晚白垩世; E1,古新世; E2,始新世; E3,渐新世; N1,中新世; Qp,更新世。Note:P2, Late Permian; T1, Early Triassic; T2, Middle Triassic; T3, Late Triassic; J1, Early Jurassic; J2, Middle Jurassic; J3, Late Jurassic; K1, Early Cretaceous; K2, Late Cretaceous; E1, Paleocene; E2, Eocene; E3, Oligocene; N1, Miocene; Qp, Pleistocene.

图3 世界蜡蝉次目化石各地质时期地理分布Fig.3 The distribution of the world’s Fulgoromorpha fossils in different geological ages注:A, 二叠纪; B, 三叠纪; C, 侏罗纪; D, 白垩纪; E, 古近纪; F, 新近纪和第四纪。Note: A, Permian; B, Triassic; C, Jurassic; D, Cretaceous; E, Paleogene; F, Neogene and Quaternary.
5 当前蜡蝉次目化石昆虫研究存在问题与展望
5.1 蜡蝉次目化石研究中存在的问题
蜡蝉次目化石昆虫的研究已接近200年,但是随着研究的不断深入,存在的问题也逐渐凸显出来,主要体现在:
(1)化石保存不完整,分类位置存疑。由于受到标本数量和保存状态的限制,蜡蝉次目昆虫化石中新的分类单元的建立许多基于单一标本,甚至数块残破标本,可信度较低;并且不同分类学家对特征认识和把握也存在一定差异,对同一化石标本的分类位置把握常常不统一;同时,翅通常为化石材料中保存完好的结构,一些支脉的分支数量、纵脉的连接方式等细微差异往往被视为重要的鉴别特征,而忽略了脉序在种内甚至同一个体左右翅之间的差异(图4)。因此造成了大量的同物异名或高级分类阶元无法确定的情况。例如:任东等(1995)建立一属两种,叶氏燕都菱蜡蝉Yanducixiusyihi和豹斑燕都菱蜡蝉Yanducixiuspardalinus,两种均只有前翅的保存,而科级阶元的划分通常还需要虫体特征,因此根据保留的特征只能将其归入蜡蝉总科,但无法定科。

图4 蜡蝉次目昆虫翅脉变异Fig.4 Venation variation in Fulgoromorpha注:缅甸琥珀中科未定CNU-HOM-MA2018004。A, 左翅; B, 右翅。比例尺A=B=0.5 mm。Note: Family incertae sedis from Burmese amber CNU-HOM-MA2018004. A, Left wing; B, Right wing. Scale bars A=B=0.5 mm.
(2)研究基础薄弱,缺少专业人员。蜡蝉次目化石昆虫研究相对零散,目前世界专门从事蜡蝉次目化石昆虫研究的专家仅几人,使得该类群化石系统研究工作进展缓慢。
(3)研究领域单一,技术手段落后。现今的蜡蝉次目化石昆虫研究领域仍主要集中在传统分类方面,分支系统学的研究方法在化石研究中还从未有过实质性的应用,有关重要特征的功能、与伴生生物之间相互关系、古生态以及古气候延展性领域的研究罕有报道,化石自身所具备的科学信息挖掘还远远不够。当前化石研究中所用仪器设备仍主要是传统光学显微镜和光学成像系统,CT、扫描电镜、能谱仪等仪器在蜡蝉次目化石昆虫研究中运用较少。
5.2 蜡蝉次目化石研究展望
蜡蝉次目昆虫化石十分丰富,其中已研究的化石标本数量仅占少数,仍然有大量研究工作有待进行,尤其在以下几个方面可深入开展:
(1)继续深入进行经典分类学研究。蜡蝉次目化石昆虫分类研究工作依然薄弱,大量化石样本有待研究。在今后的工作中,仍需广泛收集化石材料,不仅要发现新单元,更要回顾前人工作,厘定分类位置存疑的属种,同时寻找科学稳定的分类标准。
(2)全证据对蜡蝉次目昆虫系统发育关系进行研究。探究蜡蝉次目系统发育关系意义重大,当前蜡蝉次目在半翅目中系统位置仍存有争议,即蜡蝉次目和蝉次目组成的头喙亚目是否为单系(Songetal., 2012),近年来形态和分子系统学研究均未能得到一致结果(Campbelletal., 1995; Liang, 2005; Cryan and Urban, 2012);此外,蜡蝉次目至今没有稳定的分类系统,高级阶元间系统发育关系混乱不清(Urban and Cryan, 2007; Song and Liang, 2013; Bartlettetal., 2018)。因此,利用分子、形态和化石三者的证据,宏观和微观、古代和现代相结合,对它们进行系统学研究是解决问题的关键。
(3)重要特征演化的研究。蜡蝉次目昆虫重要结构特征(口器、触角、翅等)的演化能够体现它们的生物学行为的演化(如取食、拟态等),折射出其与环境之间的演化关系,今后,蜡蝉次目昆虫重要特征的功能形态和适应性演化机制、与伴生生物及生态环境的协同演化是研究的一个重要方向。
(4)多学科交叉、多技术手段探讨起源问题。在蜡蝉次目化石昆虫的研究中,需要具备宏观和微观、古生和现生相结合的研究思路,同时需要综合运用古生物学、分子生物学、生物地理学、功能形态学等学科的知识和理论,掌握扫描电镜、Macro-CT、能谱分析等多种技术手段。只有如此,才能解答蜡蝉次目昆虫的起源演化中的部分问题,为重建古环境、古地理、古气候以及验证大陆漂移等方面提供新的化石证据,进而将这一类群的研究引向深入。
