基于聚类分析的贺兰山东麓不同酿酒葡萄品种根系抗寒性综合评价
2021-05-06陈宁徐国前宋瑞李强证闻凯张军翔
陈宁 徐国前 宋瑞 李强证 闻凯 张军翔

摘要:以宁夏贺兰山东麓产区的17个酿酒葡萄品种一年生根系为试材,分别测定自由水/束缚水以及低温胁迫后根系电导率与丙二醛含量等抗寒性评价指标,计算各品种半致死温度(LT50)。结果表明,随著处理温度的降低,葡萄根系电导率与丙二醛含量呈递增趋势;根系抗寒性强的品种LT50低,自由水/束缚水值小,丙二醛增幅量小。对17个欧亚葡萄品种进行聚类分析,结合品种春季成活率,品种根系抗寒性可分为4类:一类为抗寒性强的品种威代尔;二类为抗寒性较强的品种小芒森;三类为抗寒性较弱的品种紫代夫、媚丽、雷司令、蛇龙珠、霞多丽、维欧尼、赤霞珠、品丽珠、泰娜特和西拉;四类为抗寒性弱的品种美乐、马瑟兰、小味儿多、贵人香和黑比诺。
关键词:贺兰山东麓;葡萄根系;抗寒性;聚类分析
中图分类号:S663.103.7文献标志码:A
文章编号:1002-1302(2021)04-0093-06
作者简介:陈宁(1995—),男,山东惠民人,硕士,研究方向为葡萄栽培与酿酒。E-mail:chen0605qx@163.com。
通信作者:徐国前,博士,研究方向为葡萄栽培与酿酒。E-mail:nxwine@yeah.net。
葡萄(VitisL.)是世界四大栽培果树之一,我国葡萄产量和栽培种植面积居世界前列[1],其中宁夏贺兰山东麓产区是我国酿酒葡萄栽培种植面积最大的产区,适宜的自然条件可以生产优质的葡萄酒。冬季严寒易发生冻害,葡萄根系是葡萄休眠期抗寒性最弱的器官,易遭受冻害的影响,因此根系抗寒性强弱成为葡萄越冬存活的关键因素。可以通过研究葡萄根系的抗寒性[2],了解品种间抗寒差异,制定相应的栽培技术措施提高越冬成活率。对根系抗寒性的鉴定方法有多种,从郭修武等和其他抗寒研究相关报道中选取了电导率[3-6]、半致死温度(LT50)[7-8]、自由水与束缚水的含量及比值[9-10]和丙二醛含量[4]作为鉴定根系抗寒性的重要指标,指标变化规律如下:王飞等和王文举等分别用电导率对杏和葡萄枝条测定抗寒性,发现电导率会随温度降低呈“S”曲线变化,植物抗寒性不同,电导率增加幅度不同[11-12];张倩等利用LT50测定不同葡萄种群抗寒能力,结果表明,美洲种抗寒性最强,欧亚种抗寒性最差[13];高振等以低温放热法研究葡萄根系抗寒性,结果表明,根系自由水含量越低,束缚水含量越高,自由水/束缚水比值越小,根系的LTS隶属函数度越大,其抗寒性越好,栽培品种中芳提娜抗寒性最强,西拉、佳美娜、紫代夫和泰娜特抗寒性最差[14-15];李桂荣等对不同品种枝条研究发现,可溶性糖、可溶性蛋白质以及丙二醛含量随着温度降低呈升高趋势,抗寒性不同其变化趋势不一样[16-17]。目前贺兰山东麓绝大部分葡萄园用自根苗建园,抗寒力差,和嫁接苗相比受冻害威胁较大,防止冻害发生则增加越冬埋土量,埋出土用工大,生产成本高。本研究选取17个主栽酿酒葡萄品种,测定其根系相关生理生化指标,对其抗寒性进行评价,比较品种根系抗寒性强弱,为确定葡萄抗寒能力、实现按品种埋土管理栽培、降低生产成本提供理论依据。
1材料与方法
1.1试验材料
试材选取自贺兰山东麓产区宁夏大学葡萄酒学院葡萄种植基地,该地区属温寒带干旱地区,灌溉条件为滴灌,土壤为沙壤土。共选择17个欧亚葡萄品种(表1)作为试验材料,冬季埋土前取各品种地表下20~30cm土层内粗细均匀的1年生根系,试验测定于2018年11—12月在宁夏大学葡萄与葡萄酒研究院实验室内进行。
1.2处理方法
将采集的根系去除须根,用自来水清洗2~3遍,再用去离子水清洗1~2遍,放入密封袋中,抽真空,置于4℃恒温冰箱中待测;在尽量保持根系不受破坏的情况下称取每个品种根系12g,分3份,密封,抽真空,放入上海蓝豹高低温(交变)湿热试验箱处理,处理温度分别为-12、-9、-6、-3℃,及4℃(对照组)。以4℃/h速度进行程序降温,至目的温度后持续12h,之后以同样速度回升至4℃取出样品,自封袋密封,测定抗寒指标,重复3次。
1.3抗寒性指标与测定方法
1.3.1相对电导率与半致死温度
参照电导仪法[18]进行测定,称取0.5g根系放入离心管中,加10mL蒸馏水静置12h,用雷磁DDS-307型电导仪测值记为R1,煮沸30min冷却室温测值记为R2,3次重复。计算:相对电导率=R1/R2×100%,结合相对电导率拟合Logistic方程y=k/(1+ae-bx),求方程二阶导数并令其等于零,方程曲线拐点x=lna/b即为LT50。
1.3.2生理指标测定
自由水/束缚水比值和丙二醛含量测定参照王学奎《植物生理生化实验原理与技术》中测定方法,自由水和束缚水含量在取样后立即测定,丙二醛(MDA)含量经过不同温度处理后用硫代巴比妥酸法(TBA)测定,3次重复[18]。
1.4统计分析方法
基本数据采用MicrosoftExcel进行处理编辑,统计分析用SPSS25.0软件进行处理。
1.4.1方差分析
对3次重复试验数据进行方差分析,比较每个温度处理或每个指标下17个品种之间的差异性,最终结果用平均值±标准差表示。
1.4.2聚类分析
选取-12℃温度处理下的相对电导率和MDA数据作为初始数据,对数据进行标准化=(初始值-对照值)/对照值。用标准化后的相对电导率、MDA含量、LT50、自由水/束缚水比值等4个指标对17个葡萄根系进行系统聚类,得到谱系图,对葡萄根系抗寒性进行分类。
2结果与分析
2.1低温胁迫对葡萄根系电导率和LT50的影响
表2中是所有品种低温处理后根系电导率的变化。随着处理温度的降低,所有品种根系电导率值均增大。对比4℃对照组数据发现,除美乐和黑比诺外,其余品种电导率均低于50%,威代尔低于40%,其中马瑟拉、品丽珠、霞多丽、维欧尼和雷司令之间差异性不显著,泰娜特、西拉、蛇龙珠、小芒森和贵人香之间差异性不显著;在-3℃下,所有品种电导率均高于对照,赤霞珠、紫代夫、西拉、品丽珠、媚丽、霞多丽、维欧尼和雷司令之间差异性不显著;在-6℃下,所有品种电导率均高于-3℃处理,马瑟兰、小味儿多和霞多丽之间差异性不显著;在-9℃下,所有品种电导率均高于-6℃处理,除小芒森、霞多丽和威代尔外,其余品种电导率均高于70%,美乐、马瑟兰、紫代夫、泰娜特和黑比诺之间差异性不显著;在-12℃下,所有品种电导率均高于-9℃处理,除品丽珠、小芒森和威代尔外,其余品种电导率均高于80%,赤霞珠、黑比诺、媚丽、贵人香、霞多丽和维欧尼之间差异性不显著。对比所有品种不同处理,威代尔与其他品种差异性均显著。温度下降时,对冷敏感度高的品种细胞质膜透性会增加幅度较大,若温度下降到植物所能抵抗的最低温度以下时,细胞质膜透性产生不可逆改变[19-20],膜内可溶性物质外渗,使电导率逐渐升高。
根据所得品种电导率值(不包括对照组)进行Logistic方程拟合,求得各品种LT50。由表3可知,拟合度R2均大于0.900,拟合度好,均达到显著水平。比较发现根系抗寒性強的品种是威代尔和小芒森,其LT50分别为-6.92和-5.09;抗寒性弱的品种是小味儿多和黑比诺,其LT50分别为-2.70和-2.40。对品种LT50进行品种根系抗寒性大小排序为威代尔>小芒森>蛇龙珠>紫代夫>媚丽>雷司令>维欧尼>霞多丽>泰娜特>赤霞珠>品丽珠>西拉>贵人香>美乐>马瑟兰>小味儿多>黑比诺。
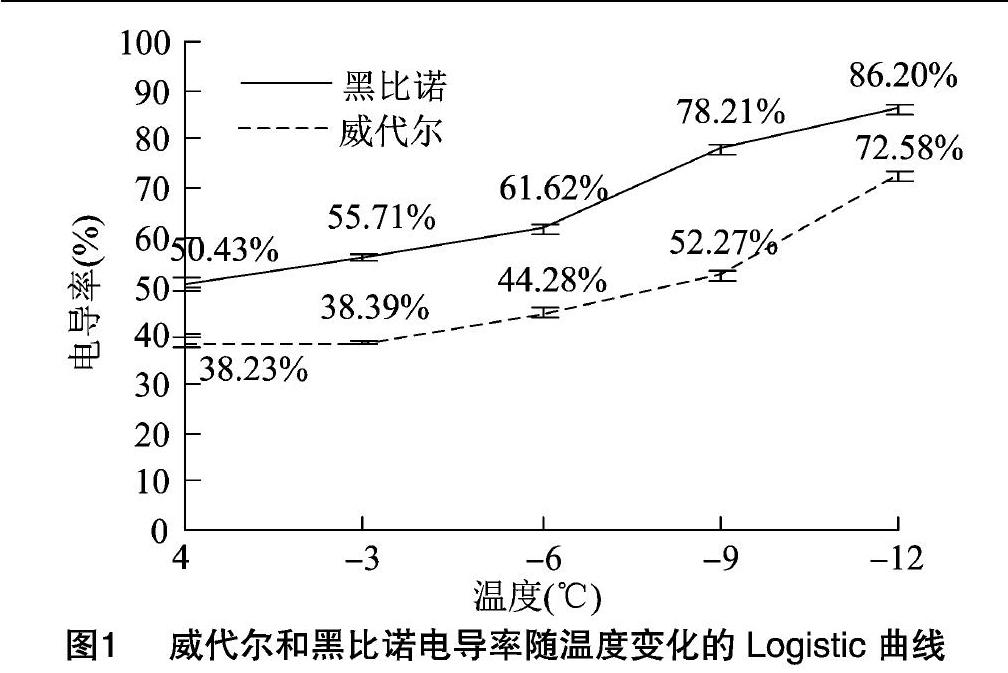
17个葡萄品种的相对电导率随着处理温度的下降而上升,且呈明显的“S”形曲线。品种根系抗寒能力不同,低温处理下电导率增加幅度不同;品种间拟合度不同,“S”形有所差异。图1显示的是抗寒性强的威代尔和抗寒性弱的黑比诺相对电导率变化规律。
2.217个品种葡萄根系自由水/束缚水含量的比较
由表4可知,自由水含量最高的是美乐,含量最低的是品丽珠,其中美乐和黑比诺差异性不显著,紫代夫、西拉、蛇龙珠、品丽珠、媚丽、小芒森、维欧尼、雷司令和威代尔差异性不显著;束缚水含量最高的是小芒森和威代尔,最低的是品丽珠,其中泰娜特、蛇龙珠、小芒森、维欧尼和威代尔差异性不显著,其余差异性显著;自由水/束缚水比值最大的美乐,最小的是威代尔,马瑟兰、小味儿多和黑比诺比值较大且差异性不显著,蛇龙珠、小芒森、维欧尼和威代尔比值小且差异性不显著。自由水/束缚水比值越大的其自由水含量越大、束缚水含量越小,抗寒性越差,比值越小的束缚水含量较高抗寒性较强。
2.3低温胁迫对葡萄根系MDA含量的影响
由表5可知,随着处理温度的降低,所有品种MDA含量逐渐增加。17个供试品种在4℃下,葡萄根系产生了少量的MDA,威代尔含量最高,泰娜特、蛇龙珠和维欧尼含量较高且差异性不显著,美乐、紫代夫和霞多丽含量较少且差异性不显著;在-3℃下,MDA含量开始增加,黑比诺和威代尔含量较高且差异性不显著,马瑟兰含量最低与其他品种差异性显著;在-6℃下,品丽珠和维欧尼含量较高且差异性不显著,美乐和霞多丽含量较低且差异性不显著;在-9℃下,除美乐外,其余品种含量均高于0.200μmol/g,泰娜特、品丽珠和黑比诺含量较高且差异性不显著;在-12℃下,MDA含量达到最大值,小味儿多、泰娜特、西拉、品丽珠、黑比诺和小芒森含量较高且差异性不显著,紫代夫含量最低。
随处理温度的降低,膜脂过氧化作用增强,MDA的含量不断增加,对葡萄根系造成的伤害逐渐增强,但不同品种间含量增加幅度不同,说明各品种间抗寒性差异显著。增幅量与根系抗寒性有密切关系,丙二醛含量增加小的品种,受损伤程度较轻,抵抗低温能力较强。威代尔、小芒森、蛇龙珠增幅量较小,分别为0.044、0.054、0.056μmol/g,抗寒性相对较强;黑比诺、小味儿多、马瑟兰增幅量较大,分别为0.083、0.091、0.097μmol/g,抗寒性相对较差。
2.4品种春季出土成活率比较
西北地区葡萄越冬需埋土防寒,越冬期间埋土厚度为30~40cm,埋土宽度为60~80cm。如表6所示,威代尔成活率最高,达90%;其次泰娜特、黑比诺、小芒森、雷司令和小味儿多成活率也达到80%以上;成活率较低的是西拉和马瑟兰,在70%以下。品种成活率普遍不高,推测可能埋土厚度不够,以及越冬期间管理不够,使得春季出土后成活率较低。
2.5聚类分析对葡萄根系抗寒性的综合评价
用标准化后的相对电导率、MDA含量、LT50、自由水/束缚水比值等4个指标数据对17个葡萄根系抗寒性进行系统聚类,得到谱系图。如图2所示,可以将葡萄根系抗寒性分为以下4类:紫代夫、媚丽、雷司令、蛇龙珠、霞多丽、维欧尼、赤霞珠、品丽珠、泰娜特和西拉特性相近为一类;美乐、马瑟兰、小味儿多、贵人香和黑比诺特性相近为一类;小芒森为一类;威代尔为一类。
3讨论与结论
低温胁迫会使细胞渗透液含量增加,随着低温的不断增大,电解质渗出率增加。本试验中,随处理温度的下降,根系电导率呈上升趋势,整体变化趋势呈“S”形,与张倩等的研究结果[13-21]一致。其中LT50反映了温度与抗寒性的数量关系[22],LT50越低,抗寒性越好;反之,LT50越高,抗寒性越差[23-24]。本研究中威代尔、小芒森、蛇龙珠和维欧尼自由水与束缚水比值小;美乐自由水/束缚水比值大。研究证明自由水/束缚水比值与抗寒性呈负相关[8-9],自由水/束缚水比值越小,抗寒性越强。在低温下细胞质膜透性减弱,丙二醛含量升高[25],其含量可以反映植物遭受逆境伤害程度大小。本研究中葡萄根系在低温处理后MDA含量及变化规律表明,随处理温度降低,各品种根系中MDA含量增大且增加幅度不同。MDA含量低且增加幅度小,抗寒性强;含量高且增加幅度大,抗寒性弱[26]。如威代尔、小芒森、蛇龙珠增幅量较小,抗寒性相对较强,而黑比诺、小味儿多、马瑟兰增幅量较大,抗寒性相对较弱。
对17个欧亚葡萄品种进行聚类分析,将品种根系抗寒性分为4类:一类抗寒性强的品种威代尔;二类抗寒性较强的品种小芒森;三类抗寒性较弱的品种紫代夫、媚丽、雷司令、蛇龙珠、霞多丽、维欧尼、赤霞珠、品丽珠、泰娜特和西拉;四类抗寒性弱的品种美乐、马瑟兰、小味儿多、贵人香和黑比诺。将结果与春季成活率对比,威代尔抗寒性最强,成活率最高;小芒森抗寒性较强,成活率较高;紫代夫、媚丽、蛇龙珠、霞多丽、维欧尼、赤霞珠和品丽珠等品种成活率在70%~80%,品种抗寒性可归为一类;马瑟兰抗寒性最弱,成活率也最低;其余品种成活率和抗寒性大致符合,与其他同类品种有差异可能是因为越冬期间防护有限导致越冬受损。葡萄根系抗寒性是受多因素综合影响,除生理指标外,还须从其他方面进行研究。此次研究通过将品种根系抗寒性进行分类,可使得越冬管理上对抗寒性相近的不同品种进行统一管理,通过抗寒性强弱来制定不同的管理措施,减少生产成本。选育种质资源还应结合其他因素来研究分析抗寒机制,为抗寒育种提供理论支持。
参考文献:
[1]2019statisticalreportonworldvitiviniculture[S].InternationalOrganisationofVineandWine,2019.
[2]宋瑞.贺兰山东麓酿酒葡萄根系抗寒性研究[D].银川:宁夏大学,2017.
[3]郭修武.葡萄根系抗寒性鉴定方法研究[J].葡萄栽培与酿酒,1994(4):26-29.
[4]付晓伟,张倩,刘崇怀,等.评价葡萄根系抗寒性指标的确定[J].果树学报,2014,31(1):52-59,封2.
[5]MittlerR.Oxidativestress,antioxidantsandstresstolerance[J].TrendsinPlantScience,2002,7(9):405-410.
[6]解越,张敏,梁飞侠,等.电导法配合Logistic方程鉴定西葫芦和黄瓜果实抗寒性[J].农业工程学报,2015,31(7):292-298.
[7]SteponkusPL.Roleofplasmamembraneinfreezinginjuryandcoldacclimation[J].PlantPhysiology,1984,35(1):543-584.
[8]刘艳萍,朱延林,康向阳,等.电导法协同Logistic方程确定不同类型广玉兰的抗寒性[J].中南林业科技大学学报,2012,32(10):69-71,78.
[9]徐伟慧,周兰娟,王志刚.外源水杨酸缓解西葫芦幼苗低温胁迫的效应[J].浙江农业学报,2013,25(4):764-767.
[10]陈佰鸿,张彪,毛娟,等.葡萄枝条水分含量变化与抗寒性关系[J].植物生理学报,2014(4):535-541.
[11]王飞,陈登文,高爱琴,等.杏品种一年生休眠枝、花、幼果抗寒的相关分析[J].西北植物学报,1999,19(4):618-622.
[12]王文举,张亚红,牛锦凤,等.电导法测定鲜食葡萄的抗寒性[J].果树学报,2007,24(1):34-37.
[13]张倩,刘崇怀,郭大龙,等.5个葡萄种群的低温半致死温度与其抗寒适应性的关系[J].西北农林科技大学学报(自然科学版),2013,41(5):149-154.
[14]GaoZ,LiJ,ZhuH,etal.Usingdifferentialthermalanalysistoanalyzecoldhardinessintherootsofgrapevarieties[J].ScientiaHorticulturae,2014,174:155-163.
[15]张文娥,王飞,潘学军.应用隶属函数法综合评价葡萄种间抗寒性[J].果树学报,2007,24(6):849-853.
[16]李桂荣,连艳会,程珊珊,等.低温胁迫下不同无核葡萄品种抗寒性的分析[J].西南农业学报,2018,31(11):2399-2406.
[17]何伟,艾军,范书田,等.葡萄品种及砧木抗寒性评价方法研究[J].果树学报,2015,32(6):1135-1142.
[18]王学奎,黄见良.植物生理生化实验原理和技术[M].3版.北京:高等教育出版社,2015.
[19]LyonsJM,RaisonJK.Oxidativeactivityofmitochondriaisolatedfromplanttissuessensitiveandresistanttochillinginjury[J].PlantPhysiology,1970,45(4):386-389.
[20]高振.基于温度-伤害度关系分析葡萄抗寒性及其影响因素[D].泰安:山东农业大学,2014.[HJ2mm]
[21]牛锦凤,平吉成,王振平,等.几个鲜食葡萄品种抗寒性的比较研究[J].北方园艺,2005(6):63-65.
[22]苏维埃,宓容钦,王文英,等.植物抗性指标的数量化研究——Ⅰ.胁强、时间与胁变三者在植物抗冻性中的数量关系[J].中国科学(化学):1987,17(10):1058-1067.
[23]曹建东.9个葡萄砧木和品种的抗寒性及耐鹽性鉴定[D].兰州:甘肃农业大学,2010.
[24]周香云,李爽,李连国,等.低温胁迫对8种葡萄砧木根系细胞膜透性影响的研究[J].内蒙古农业大学学报(自然科学版),2012,33(4):51-53.
[25]GaoSM,ChengPJ,GuoHH,etal.Studyoncoldacclimationandfreezing-tolerancemechanismofAucubajaponicacv.Variegata[J].ActaBotanicaBoreali-OccidentaliaSinica,2003,23(12):2113-2119.
[26]曹建东,陈佰鸿,王利军,等.葡萄抗寒性生理指标筛选及其评价[J].西北植物学报,2010,30(11):2232-2239.
