转录组的黄山栾树MYB基因家族鉴定及表达1)
2021-04-27吕运舟董筱昀梁珍海孙海楠黄利斌蒋泽平
吕运舟 董筱昀 梁珍海 孙海楠 黄利斌 蒋泽平
(江苏省林业科学研究院,南京,211153)
MYB转录因子家族广泛参与植物次生代谢、细胞分化、抗逆性等生理生化过程,其成员都具有高度保守的DNA结合结构域,由52个氨基酸构成的重复基序组成[1-3]。根据重复基序数目(R)可以分为1R-MYB、R2R3-MYB、3R-MYB、4R-MYB4种类型[3]。目前,MYB转录因子已在拟南芥(Arabidopsisthaliana)等模式物种中得到分离和功能鉴定[2-3],在毛果杨(Populustrichocarpa)[4]、杉木(Cunninghamialanceolata)[5]、楠木(Phoebezhennan)[6]、桑树(Morusnotabilis)[7]等乔木树种基因组或转录组水平得到鉴定。
植物叶片颜色是由叶片中色素的组分、含量及分布决定[8-9],包括叶绿素、类黄酮类和类胡萝卜素类色素。类黄酮类色素和类胡萝卜素的种类和含量决定了植物界的缤纷色彩,许多研究结果已经揭示了它们的生物合成过程[10]。MYB转录因子对黄酮类化合物和类胡萝卜素的生物合成起到重要的调控作用,如拟南芥MYB基因家族的MYB11[11]、MYB12[12-13]、MYB112[14]通过调控CHS、CHI、F3H和FLS1基因的表达实现对花青素等类黄酮类化合物生物合成的调控作用;彩艳龙头花(Mimuluslewisii)R2R3-MYB类基因RCP1可以激活所有类萝卜素合成基因而激活花瓣中类胡萝卜素的生物合成[15]。此外,Amponmah et al.[16]发现猕猴桃(Actinidiadeliciosa)MYB7基因在转录水平正向调控类胡萝卜素和叶绿素的生物合成。叶色变异植株是彩叶新品种和植物色素合成功能基因组学研究的重要材料[9,17]。‘金焰彩栾’(Koelreuteriabipinnata‘Jinyan’)是黄山栾树(Koelreuteriabipinnatavar.integrifoliola)的天然黄化突变体,枝干呈橙黄色,春季叶片橙红色,秋季全株叶片金黄,具有极高的观赏价值[18-19]。本研究以‘金焰彩栾’及其原种黄山栾树转录组数据入手,结合叶片发育不同时期高通量测序数据,挖掘鉴定黄山栾树MYB转录因子家族基因,并对其预测蛋白特征及表达特征进行比较分析,旨在鉴别与类黄酮类或类胡萝卜素调控相关MYB转录因子,探讨‘金焰彩栾’叶片呈色的分子机制。
1 材料与方法
1.1 供试材料
供试黄山栾树及其黄叶变异株‘金焰彩栾’种植于江苏省林业科学研究院试验苗圃内。
1.2 RNA提取、cDNA文库建立及测序
分别从固定样株中采集春季新叶、夏季成熟叶及秋季变色期叶片,分别为T1(‘金焰彩栾’秋季叶片)、T2(实生对照秋季叶片)、T3(‘金焰彩栾’春季叶片)、T4(实生对照春季叶片)、T5(‘金焰彩栾’夏季叶片)及T6(实生对照夏季叶片),液氮速冻;总RNA按照Trizol试剂盒(Thermo Fisher,USA)提取,测序文库构建及测序由北京百迈客生物科技有限公司完成,使用Illumina HiSeq 2500高通量测序技术进行测序[17]。
1.3 MYB基因鉴定
利用pfam数据库PFAM(http://pfam.xfam.org/)的MYB基因家族结构域hmmer隐马尔科夫模型文件(PF00249)及hmmsearch软件对转录组数据中所有的蛋白序列进行搜索,筛选期望值<0.001为候选基因家族基因;手动确认候选基因中的结构域序列,利用关键“MYB”进行检索,结合NCBI CDD(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)工具,删除不含MYB结构域或重复序列。
1.4 蛋白序列及系统进化分析
从拟南芥数据库TAIR(https://www.arabidopsis.org/)下载模式植物拟南芥的MYB基因蛋白序列。提交MYB蛋白序列至ExPASy(http://web.expasy.org/protparam/),ProtParam在线分析蛋白序列性质。利用多序列比对软件muscle 3.8.31对97个黄山栾树和130个拟南芥MYB蛋白进行多序列比对分析,然后用软件MEGA7.0构建邻接法(neighbor joining,NJ)系统进化树,使用p-distance距离模型构建,选择成对删除选项,bootstrap值设置为1 000。
1.5 表达特征分析及验证
利用97个MYB候选基因在黄山栾树转录数据,包括‘金焰彩栾’和普通黄山栾树的3个时期(春季新叶、夏季成熟叶和秋季衰老叶)的基因表达数据,并对表达量数据FPKM[20]进行归一化转换,然后绘制热图。
此外,选取10个与类黄酮类及苯丙氨酸类化合物代谢调控相关的MYB候选基因,选用看家基因KbGAPDH为内参[21]。定量PCR引物利用Premier 5在非保守域设计,引物信息参见表1;RNB100 RNA提取试剂盒(SIGMA)提取以上6个组织总RNA,逆转录产物cDNA稀释10倍,使用SYBR Premix Ex TaqTM(TAKARA)定量试剂盒,在CFX-96荧光定量PCR仪(Bio-Rad)上进行定量PCR。反应体系为20 μL,其中SYBR mix 10 μL,上下游引物(10 μmol·L-1)各0.8 μL,cDNA模板1.0 μL,并加6.4 μL ddH2O,每个样本3次重复。PCR扩增反应程序:95 ℃预变性30 s,95 ℃变性5 s,60 ℃退火20 s,共40个循环。
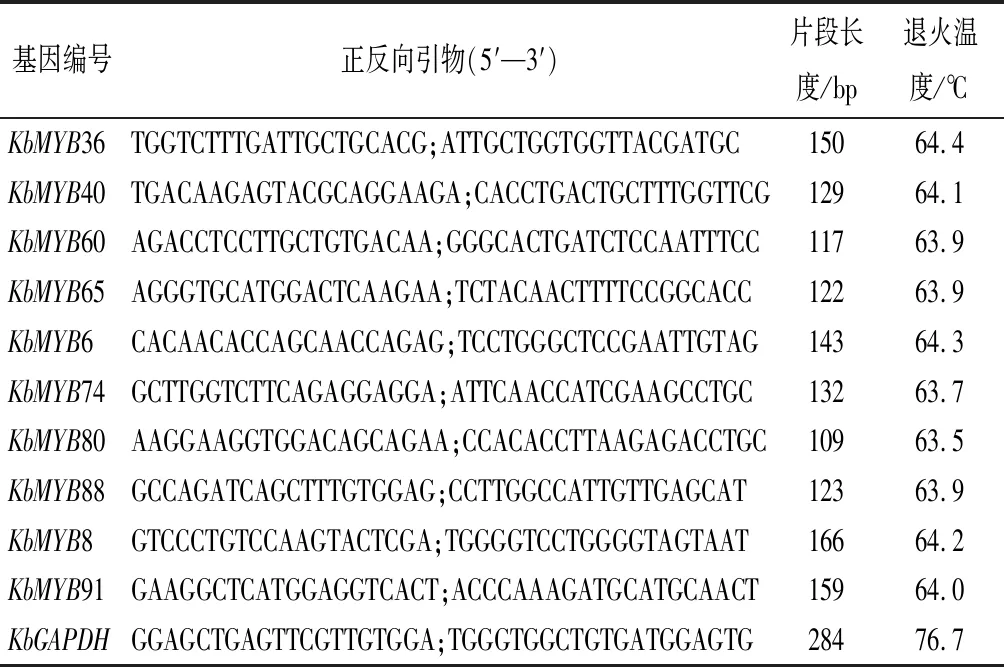
表1 用于定量分析的引物序列
2 结果与分析
2.1 测序结果
经过测序质量控制,共得到31.44 Gb有效数据,各样品Q30碱基百分比均不小于90.74%。经Trinity软件组装后,共得到175 881条转录本和94 125条单基因簇,转录本与单基因簇的N50分别为2 791和1 340,组装完整性较高。生物学分析比对发现,共有47 341条单基因簇在与NR、Swiss-Prot、GO、COG、KOG、KEGG数据库得到注释,这些序列信息为后续研究提供数据基础。
2.2 黄山栾树MYB基因家族的鉴定及编码蛋白
基于黄山栾树及其突变体叶片转录组数据的本地Blast比对检索,在NR、NT、Swiss-Prot等数据库进行注释,再经NCBI CDD工具检验,去除过短和重复冗余的序列后,共获得97个黄山栾树MYB转录因子,分别命名为KbMYB1~KbMYB97。从表2可以看出,包括48个1R-MYB类转录因子,47个R2R3-MYB类转录因子和2个3R-MYB类转录因子得到鉴定,预测编码蛋白长度介于52(KbMYB89)~1780KbMYB25)个氨基酸,理论分子质量大小介于52.0~193.35 ku,理论等电点范围在4.51(KbMYB16)~11.87(KbMYB51)。
2.3 黄山栾树MYB转录因子进化分析
利用MEGA7.0构建的黄山栾树与拟南芥MYB转录因子家族系统进化树,拓扑结构如图1所示,可以分为23亚组,亚组G23仅包含22个黄山栾树转录因子,亚组G3、G6和G21仅包含拟南芥MYB转录因子。19个亚组由黄山栾树和拟南芥MYB转录因子共同组成,其中亚家族G2、G9、G14中只有1个黄山栾树MYB基因家族成员。此外,在同一亚组中拟南芥基因和黄山栾树基因之间彼此相互分开,说明两者遗传关系较远。已有研究表明亲缘关系较近的MYB转录因子具有相似生物学功能[2-3],基于拟南芥中的研究,G1、G2、G4、G7、G8、G9和G13分别与次生生长、生长发育及胁迫应答等调控有关。
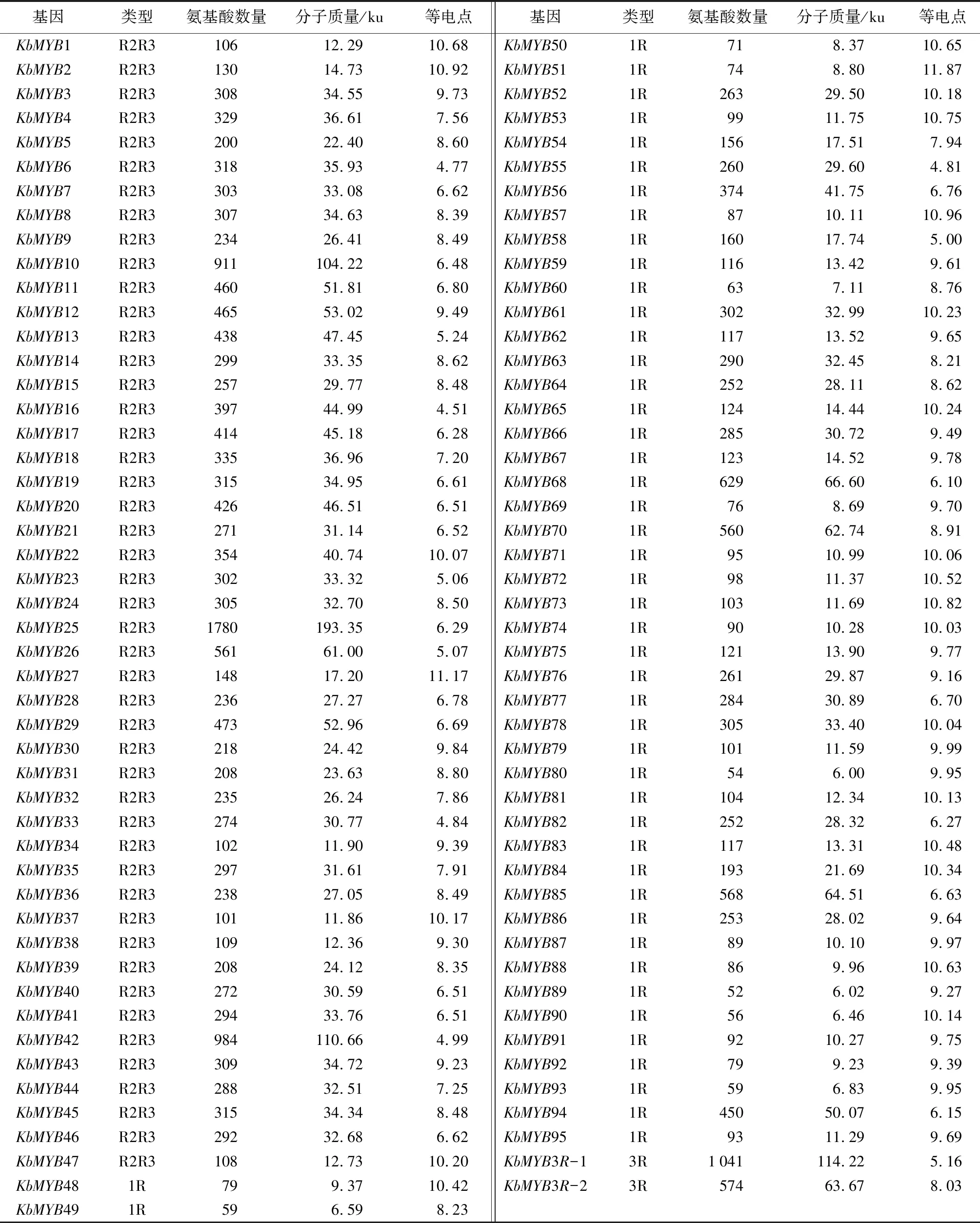
表2 黄山栾树MYB基因家族编码蛋白信息

图1 黄山栾树和拟南芥MYB类蛋白家族的进化分析
2.4 MYB转录因子在‘金焰彩栾’叶片呈色过程中的表达模式
利用黄山栾树及其黄叶突变体‘金焰彩栾’春、夏、秋3个时期的叶片转录组数据库,提取97个MYB基因对应的RPKM值(代表每百万片段中来自于某基因每千碱基长度的片段数),并经归一化处理制作Heml热图(如图2所示)。结果表明,MYB基因在‘金焰彩栾’及普通黄山栾树不同时期叶片中表达差异明显。根据表达模式特征可以将黄山栾树MYB基因家族成员分为4类,I~III类在‘金焰彩栾’和普通栾树中表达模式基本一致,其中I组为春季叶片中表达丰度较少,在夏季上调表达,秋季又下调表达;II组为春季和夏季叶片中表达量较少,在秋季上调表达;III组为春季叶片中表达丰度较高,在夏季和秋季叶片中表达下调。而IV组中的MYB基因在‘金焰彩栾’的3个时期叶片中表达量均较高,而在对照中呈现春季叶片表达丰度较高,其它两个时期较低,这些MYB基因可能参与调控突变体叶片呈色过程。
2.5 类黄酮类等次盛化合物合成调控候选MYB基因表达
根据MYB转录因子表达特征及聚类分析结果,挑选与拟南芥类黄酮类、苯丙烷类等次生代谢合成调控相关MYB转录因子蛋白同源性较近的10个黄山栾树MYB候选基因。实时定量分析结果表明(表3),春季叶片相对高表达的MYB转录因子包括KbMYB40,KbMYB65,KbMYB80和KbMYB91,其中KbMYB80在‘金焰彩栾’表达量相对较高,而KbMYB40相反。候选基因KbMYB6和KbMYB8,预测与花青素等类黄酮类化合物合成正向调控相关,表达模式一致,在普通栾树夏季成熟叶中表达较高,秋季表达下降。花青素合成负向调控候选基因KbMYB60在供试样本中表达量较为一致,呈高水平状态。而苯丙烷类合成调控候选基因KbMYB88在‘金焰彩栾’和普通栾树的春、秋时期叶片中表达基本一致,在夏季叶片中表达量增加,且在普通栾树中表达量最高。
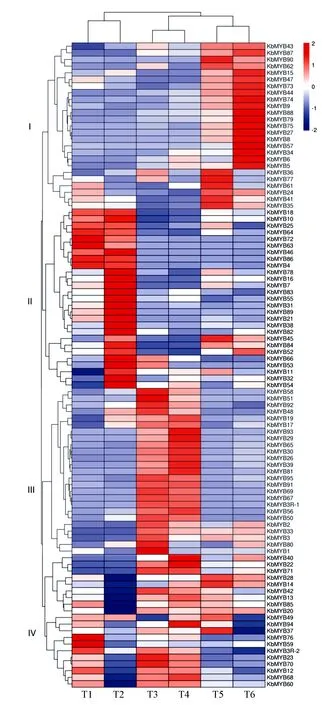
T1、T3、T5与T2、T4、T6分别为‘金焰彩栾’与对照的秋季、春季及夏季叶片。
3 结论与讨论
黄山栾树是我国优良的乡土树种,具有多种经济价值和园林观赏价值,是园林绿化、庭院配植的理想树种[18]。MYB转录因子在类黄酮类、类萝卜素及苯丙烷类化合物代谢调控关系密切,在植物叶片、花瓣及果实呈色调控中发挥至关重要的作用[22-26]。通过对黄山栾树叶片转录组数据挖掘,共鉴定出97个MYB基因,预测编码蛋白长度介于52~1 780个氨基酸。通过与130个拟南芥MYB(AtMYB)蛋白聚类分析,预测到多个MYB基因可能参与类黄酮类、类胡萝卜素类和苯丙烷类化合物代谢通路功能基因表达调控。Dubos et al.[3]根据结构及功能将拟南芥MYB基因家族分为27个亚家族,其中AtMYB11、AtMYB12与AtMYB112[11-14];AtMYB75[27]、AtMYB90[3]与AtMYB123[28]分别参与黄酮和花青素生物合成过程。共有13个黄山栾树MYB基因与它们归为4个亚组(G1、G2与G4),涉及KbMYB5、KbMYB9、KbMYB80、KbMYB91,KbMYB6、KbMYB8、KbMYB44、KbMYB79、KbMYB93与KbMYB74、KbMYB40、KbMYB65,它们可能参与黄酮和花青素的生物合成调控。AtMYB58、AtMYB63[29],AtMYB46[30]和AtMYB72[30]参与苯丙烷类化合物代谢而影响植物次生细胞壁合成,间接影响黄酮类代谢通路。由此推测,KbMYB16和KbMYB88参与黄山栾树叶片细胞次生壁合成过程。
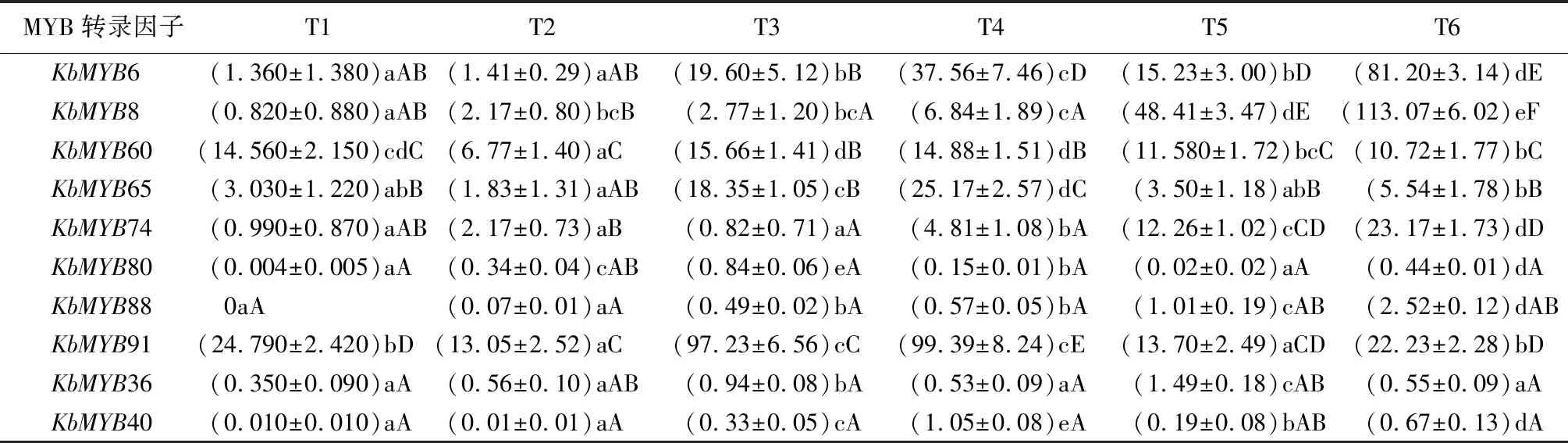
表3 候选MYB基因实时荧光定量PCR分析
‘金焰彩栾’是黄山栾树实生变异的黄化突变体,观赏价值极高[18]。研究发现,金焰彩栾’整个生长期中的叶绿素a、叶绿素b、总叶绿素含量和类胡萝卜素、花色素苷含量均显著低于原种,其光合能力减弱,生长较原种受到部分影响[19]。此外,研究发现脱落醛氧化酶基因的下调表达导致了类胡萝卜素含量降低,叶绿素a酸酯氧化酶基因的下调表达与叶绿体合成受阻有关[17],而在功能酶转录水平调控上也存在差异。表达模式分析发现,有25个MYB基因在春季叶片中表达较高,有21个在‘金焰彩栾’春、夏、秋叶片中表达量均高于普通栾树,这些基因可能参与了‘金焰彩栾’叶片呈色调控。过表达AtMYB60可以抑制植物花青素生物合成[31-32],KbMYB60与其进化关系最近,且在‘金焰彩栾’各时期叶片中表达均较高,预测其参与了花青素生物合成的负调控。进一步验证此类MYB基因功能,有助于揭示‘金焰彩栾’叶片变异的调控机理,为后续彩叶植物新品种开发提供分子基础。
