抚育间伐对针阔混交天然次生林生物量及碳密度的影响1)
2021-04-27王子纯李耀翔孟永斌王晨
王子纯 李耀翔 孟永斌 王晨
(东北林业大学,哈尔滨,150040)
抚育间伐是以促进森林生长、提高林木质量为目的,它是一种以培育目标树,对林木进行伐密留疏、伐坏留好的作业。抚育间伐必然影响森林种群的结构,从而影响林分生物量、碳密度及种群关系[1-3]。天然次生林是中国森林的重要组成部分,对维护生态平衡有着较大的贡献,其在林分环境、结构组成和生产力等方面与人工林和原始林显著不同[4],目前对天然林分别在土壤性质[5]、水文功能[6]、林下植被[7]、生境[8]等方面展开了较多研究,也有学者对间伐后林分碳密度变化展开研究,发现林分碳密度与树种、培育措施、间伐强度和间伐后持续时间等多种因素有关[9]。本研究选取2012年经3种间伐强度(15%、25%、35%)进行抚育间伐的小兴安岭试验样地及对照样地(间伐强度为0),于2019年测定抚育间伐8 a后试验样地的生物量、碳质量分数、碳密度,分析不同间伐强度对林分生物量、碳密度的影响,旨在为碳库管理及科学经营提供参考。
1 研究区概况
研究区域位于小兴安岭地区的伊春市带岭林业实验局东方红林场414林班,地理坐标为东经129°5.150′~129°5.205′、北纬46°52.511′~46°52.827′。地处中温带,属于大陆性湿润季风气候,夏季短且温热湿润,冬季长且干燥寒冷、多雪,全年平均气温1.4 ℃。降水主要集中在7—8月份,年平均降水量为661 mm。试验样地平均海拔493.36 m,位于山的中腹。研究区林分类型为针阔混交天然次生林,主要树种有冷杉(Abiesfabri(Mast.)Craib)、色木槭(AcermonoMaxim.)、云杉(PiceaasperataMast.)、青楷槭(AcertegmentosumMaxim.)、水曲柳(FraxinusmandshuricaRupr.)、黄波椤(PhellodendronamurenseRupr.)、红松(PinuskoraiensisSieb. et Zucc.)、紫椴(TiliaamurensisRupr.)、杨树(PopulusL.)、山桃(PrunusdavidianaFranch)、花楷槭(AcerukurunduenseTrautv. et Mey.)等。
2 研究方法
2.1 样地设置
天然林试验区于2012年进行带状抚育间伐。本次试验选择小兴安岭天然林试验区内3个抚育间伐强度(以林分密度计),分别近似为15%(样地编号T15)、25%(样地编号T25)、35%(样地编号T35),应用下层抚育法,采伐濒死木、被压木及干型不良的林木,同时在间伐区进行了补植作业,补植树种包括红松、云杉、落叶松[10]。2019年8月进行样地复查,在3种间伐强度的样地内,分别设置20 m×20 m的标准试验样地各3块,在保留带未间伐林分选取与间伐林分条件相似的林分设置3块20 m×20 m标准试验样地作为对照样地(CK),共计12块样地。试验样地均在同一坡面,坡向西北,坡度14°左右,平均林龄70 a,GPS记录海拔坐标。间伐林分基本状况见表1。
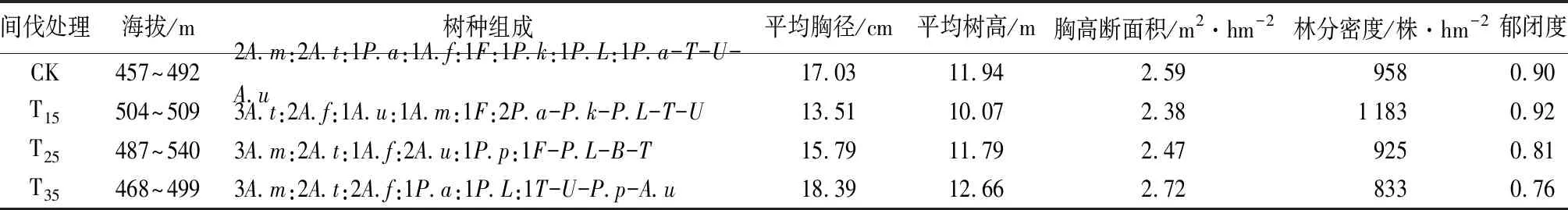
表1 间伐林分基本状况
2.2 优势树种的选择
对样地内林木进行每木检尺。检尺内容包括树种、胸径、树高、冠幅、郁闭度、林分生长状况等。本次试验样地树种组成较复杂,每木检尺数据结合通用二元材积模型[11]可得各树种蓄积量。由表2可知,蓄积量较大的树种,依次为云杉、冷杉、色木槭、青楷槭,累计蓄积量高达83.8%。因此,选取以上4个树种作为该林分的优势树种进行生物量估算和碳质量分数测定。
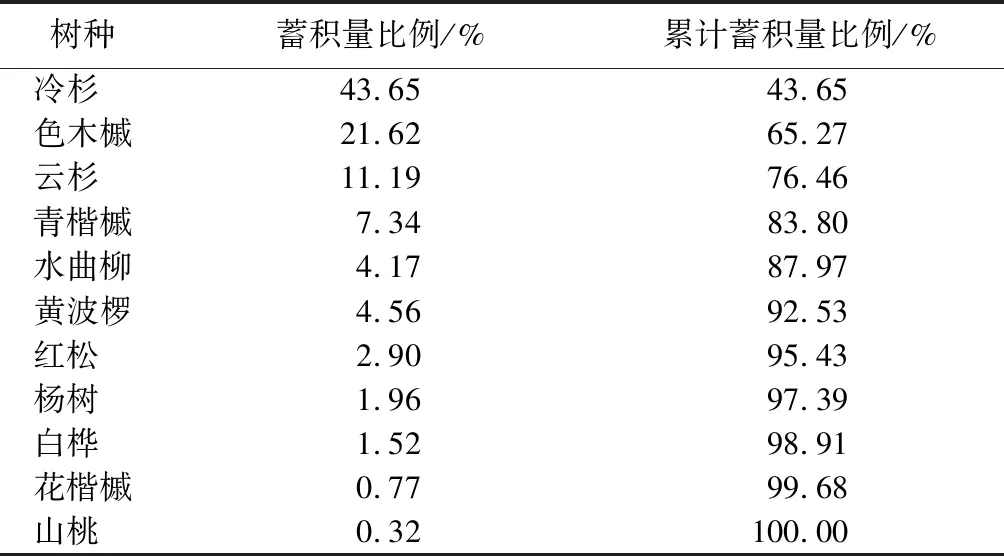
表2 不同树种的蓄积量分布
2.3 样品的采集与测定
样品采集:2019年8月份,在每个样地内按径阶在每个优势树种里分别选取2~3株标准木,对其干、枝、叶、皮进行取样。树干、树皮,分别在树干基径处、树干胸径处、树梢头处进行取样。树干用生长锥(产于瑞典,长度350 mm)进行取样;树枝、树叶使用枝剪从不同高度取样,其中树枝按照粗细的比例截取,树叶从枝条的不同部位取样。采回的样品进行野外称量,用密封袋密封并做好标记,用冰袋保鲜带回用于后续实验测定分析。
生物量的估算:利用东北林区现有的林木生物量方程,结合每木检尺调查数据,计算出各标准试验样地内优势树种的生物量。云杉、冷杉生物量模型采用胡海清[12]建立的小兴安岭典型林分生物量模型,色木槭、青楷槭生物量采用王传宽[13]建立的东北林区10种单木生物量模型进行估算。
碳质量分数的测定:国际通用的林木碳质量分数为50%,但不同森林类型及不同树种间碳质量分数存在差异[14]。本研究在小尺度上进行了树木碳质量分数的测定。将采回的样品带回实验室,放入烘箱80 ℃烘至恒质量,将已烘干的试验样品用粉碎机粉碎,为保证取样均匀,采用三次粉碎法制样,过60目筛(筛孔尺寸0.25 mm)。样品碳质量分数的测定,采用重铬酸钾氧化-外加热法,试验时用石英砂做空白对照,每个样品做3组平行试验以减小误差。
2.4 林分碳密度计算方法
样品碳质量分数为样品碳质量与样品干质量比值的百分数[12],单木各器官生物量乘以该树种在相应器官的碳质量分数合计即为单木碳储量[15],累计1个样地的单木碳储量则为该样地总碳储量,单位面积的总碳储量则为碳密度[16],单位为t/hm2。碳密度计算公式为ρ=(W/S)w,ρ为所求器官的碳密度(单位为t/hm2)、W为该器官的总生物量(单位为t)、S为样地面积(单位为hm2)、w为该器官的碳质量分数。
2.5 数据处理
采用Excel 2010、Origin85进行数据整理、分析,用SPSS 17.0进行单因素方差分析及邓肯(DunCan)多重比较差异显著性检验(α=0.05)。
3 结果与分析
3.1 不同间伐强度对林木各器官生物量的影响
由表3可知,8 a后不同间伐强度抚育间伐的样地,林木各器官生物量分配一致,由大到小均依次为树干、树枝、树叶、树皮,树干生物量占总生物量比例为70.62%~77.03%、树枝生物量占总生物量比例为18.88%~22.63%、树叶生物量占总生物量比例为2.71%~3.72%、树皮生物量占总生物量比例为2.31%~3.72%。树叶、树皮在生物量中所占比例相差很小,但树皮生物量所占比例最小,这与刘之洲[17]、徐美玲[18]得出的结果基本一致。
间伐可以改变生物量在林木各器官的分配。对照样地中,树干生物量占总生物量的70.62%;间伐整体可以使树干生物量所占比例增大,当间伐强度为15%、25%、35%时,树干生物量所占比例先增大再减小,分别为74.15%、77.03%、73.85%,依次比对照样地增加了3.53%、6.41%、3.23%。树枝生物量则相反,对照样地中,树枝生物量占总生物量的22.63%;间伐整体使树枝生物量所占比例减小,当间伐强度为15%、25%、35%时,树枝生物量先减小再增大,分别为19.41%、18.88%、19.74%,依次比对照样地减小了3.22%、3.75%、2.89%。树叶、树皮的生物量所占比例几乎不变。
树干生物量随间伐强度增加呈“J”型变化,其中,当间伐强度为15%时树干生物量最小,为59.63 t·hm-2,比对照样地减少了38.36%;间伐强度为25%时,树干生物量比对照样地减少了24.33%;当间伐强度为35%时,树干生物量比对照样地增加了13.12%。
对照样地树枝生物量为31.00 t·hm-2,大于各间伐样地的树枝生物量。随间伐强度增加,树枝生物量变化呈先减小再增大的“V”型趋势,其中,间伐强度为15%时,树枝生物量为15.61 t·hm-2,减小程度最大,比对照样地减少了49.65%;间伐强度为25%时,减小程度稍小,减少42.13%;间伐强度为35%时,树枝生物量最接近对照样地,为29.24 t·hm-2,仅减少了5.68%。
随间伐强度增大,树叶生物量变化呈“J”型趋势。其中,间伐强度为15%时,树叶生物量最小,为3.13 t·hm-2,与对照样地相比减少程度最大,减少32.54%;间伐强度为35%时,树叶生物量最大,为5.75 t·hm-2,比对照样地增加了23.92%。间伐处理对树皮生物量无显著影响。
地上部分的林木总生物量,随间伐强度的增大呈“J”型变化,其中,15%、25%间伐强度样地生物量,分别比对照样地减少了41.30%、30.63%,差异显著;35%间伐强度样地林木生物量,比对照样地增加了8.15%,但无显著差异。由此可见,小兴安岭针阔混交天然次生林间伐8a后,35%间伐强度最有利于林分恢复,可以使林分生物量增加。

表3 不同间伐强度的林分生物量 t·hm-2
3.2 不同间伐强度对林木各器官碳质量分数的影响
在林木各器官中,树干碳质量分数几乎均高于其它组分,并且树干是林木各器官中生物量占比最大的部分,说明树干是树木碳的主要载体。云、冷杉、青楷槭的树干碳质量分数,均在间伐强度为25%时最大,分别为52.27%、49.50%、46.55%;在所有样地中,色木槭树干碳质量分数随间伐强度的增加呈倒“J”型变化趋势,在间伐强度为15%时达到最大,为45.12%。林木的枝、叶、皮碳质量分数,随间伐强度的增大无明显规律变化。

表4 不同间伐强度的林木各器官碳质量分数
间伐会直接改变林分密度,从而间接改变林内光照、土壤条件、水分条件等因子,进而影响林木碳质量分数。林木碳质量分数还由树种本身遗传特性决定,因此,在同一地区相同间伐强度处理时不同树种碳质量分数亦有所差异(见表5)。在各间伐处理样地中,不同树种碳质量分数(所有器官碳质量分数均值)不同,其中:对照样地、间伐强度为15%的样地,不同树种碳质量分数由大到小依次为冷杉、云杉、色木槭、青楷槭;间伐强度为25%、35%的样地,不同树种碳质量分数由大到小依次为冷杉、云杉、青楷槭、色木槭。
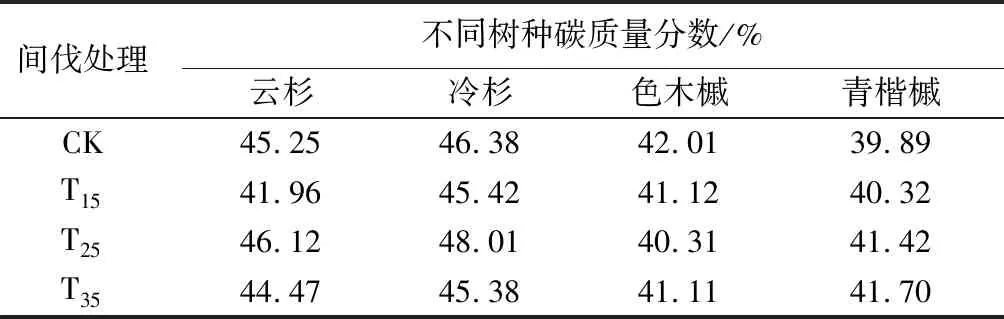
表5 不同间伐强度样地的各树种碳质量分数
3.3 不同间伐强度的林木各器官碳密度的变化
抚育间伐虽能改善林内结构,促进林木个体生长,但在间伐8 a后,因林木个体加速生长,导致的乔木层碳密度增加量难以补偿因抚育间伐移出的乔木层碳密度损失量,最终也会导致间伐处理样地碳密度减少。
由表6可见:在对照样地中的树干碳密度最大,随间伐强度增加,间伐样地与对照样地之间树干碳密度的差异逐渐缩小,15%、25%、35%间伐强度的树干碳密度,分别比对照样地减少了18.48%、9.88%、7.51%,彼此间无显著差异;树枝碳密度在对照样地最大,且显著大于15%、25%、35%间伐强度的样地;树叶碳密度在15%间伐强度样地最大,树皮碳密度则在35%间伐强度样地最大,但树叶和树皮的碳密度与其它间伐处理样地间无显著差异。地上部分碳密度在对照样地最大,15%、25%、35%间伐强度样地与对照样地的碳密度差异逐渐缩小,依次比对照样地减少了21.43%、12.12%、8.63%;其中25%、35%间伐强度样地和对照样地三者间的碳密度无显著差异。不同间伐强度林木碳密度,在林木各器官的分配特征基本一致,由大到小依次为树干、树枝、树叶、树皮,树干碳密度占总碳密度的73.57%~76.34%、树枝碳密度占总碳密度的16.48%~20.96%、树叶碳密度占总碳密度的3.72%~4.78%、树皮碳密度占总碳密度的2.04%~2.56%。

表6 不同间伐强度样地的林木碳密度
3.4 不同树种组成林分的生物量及碳密度
受自然及人为因素影响,各样地生物量、碳密度也存在差异。自然因素包括海拔、气候、降水、土壤等因素。本研究中的人为因素,主要指因间伐造成的树种组成的改变、径级分布的变化。
由表7可见,优势树种中的阔叶树种与针叶树种在所有样地中的组成,均为6阔4针。但不同样地中的色木槭、青楷槭、云杉、冷杉组成比例不完全相同,在对照样地、间伐强度为15%和25%样地中,云杉和冷杉组成均为2云2冷,但色木槭和青楷槭的组成有所差异,分别为3色3青、1色5青、4色2青。
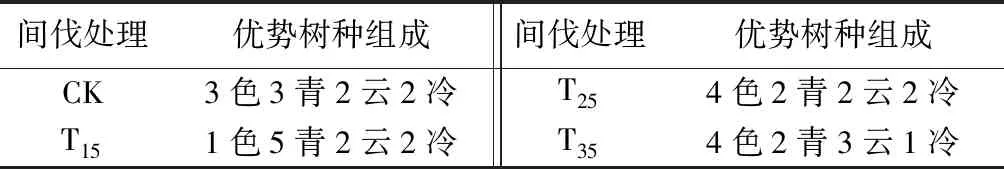
表7 不同间伐强度样地的优势树种组成
由表8可见,间伐强度为15%的样地,阔叶树种生物量为38.61 t·hm-2,小于对照样地。说明间伐强度15%样地的1色5青2云2冷的树种组成,不利于样地生物量累积;在间伐强度为25%和35%的样地,青楷槭和色木槭的组成均为4色2青,但云杉和冷杉的组成有所差异,分别为2云2冷、3云1冷。在间伐强度为35%的样地,针叶树种生物量最大,此时阔叶树种和针叶树种的生物量分别占总生物量的52.25%、47.75%,说明间伐强度为35%样地的4色2青3云1冷树种组成,优于间伐强度为25%样地的4色2青2云2冷树种组成,有利于生物量增加。
随间伐强度的增大,阔叶树种和针叶树种的生物量均呈“V”字型变化,且阔叶树种的生物量在间伐强度为15%样地的最小,针叶树种生物量在间伐强度为25%样地的最小。在对照样地、间伐强度为25%和35%样地中,阔叶树种生物量比针叶树种大36.23%、27.84%、8.63%,而在间伐强度为15%样地,阔叶树种生物量比针叶树种生物量减小了8.26%。
因不同树种固碳特性不同,碳密度也有所差异。在对照样地、间伐强度为25%及35%的样地中,阔叶树种的碳密度比针叶树种增加21.67%、17.05%、32.17%;在间伐强度为15%的样地中,阔叶树种的碳密度仅为10.07 t·hm-2,比针叶树种碳密度小39.01%。
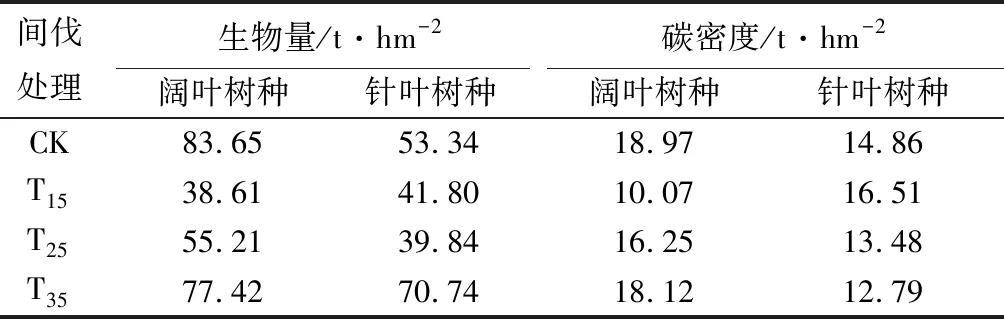
表8 不同间伐强度样地阔叶树种和针叶树种生物量及碳密度
3.5 不同径级结构林分的生物量及碳密度
3.5.1应用韦伯(Weibull)分布函数对不同间伐强度样地径级分布进行拟合
针对针阔混交林直径分布曲线类型较多、变化复杂的特点,选择适应性强、灵活性大的韦伯分布函数对直径分布进行拟合描述。用式Pn=A{1-exp[-(k(D-Dc))d]}(D为对应径阶直径,Pn为各径阶株数累计百分数)分别求出对照样地、间伐强度为15%、25%、35%样地直径分布的分布参数A、Dc、d、k及相关系数R2(见表9)。拟合结果D对Pn的相关系数R2很高,均在0.99以上,表明用韦伯分布函数拟合小兴安岭针阔混交天然次生林直径分布,效果较好。
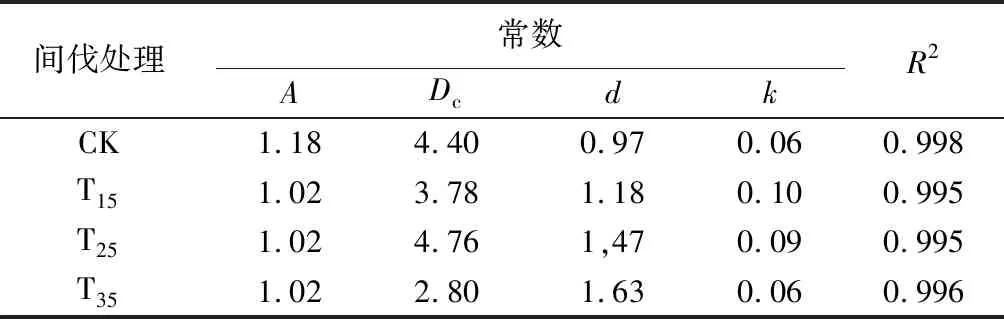
表9 不同间伐强度样地径级分布用韦伯分布函数拟合结果
3.5.2 径级结构对林分生物量及碳密度的影响
依据实际情况将各样地林木按胸径(D)大小分为小径级(5 cm≤D<16 cm)、中径级(16 cm≤D<26 cm)、大径级(D>26 cm),按径级分布计算各样地生物量及碳密度分布。
由表10可见,间伐改变了林分的径级分布。与对照样地相比,间伐强度为15%的样地多出11.37%的小径级林木,结合树种组成可知,该样地存在大量小径级青楷槭,十分符合次生林的特点。间伐强度为25%的样地中,大径级林木所占比例最少,仅为10.48%,比对照样地减少了5.49%。间伐强度为35%的样地中,中径级、大径级林木所占比例增加,共占57.93%,是4块样地中唯一超过小径级林木比例的样地。

表10 不同间伐强度样地林木径级结构分布
由表11可见,间伐强度为25%样地的小径级林木生物量,显著大于其余样地小径级林木生物量,分别比对照样地、间伐强度为15%的样地、间伐强度为35%的样地增加了60.24%、44.7%、43.74%,其中间伐强度为15%的样地与间伐强度为35%的样地,小径级林木生物量无显著差异;间伐强度为35%样地的中径级林木生物量最大,且显著大于其余样地中径级林木生物量,与对照样地、间伐强度为15%的样地、间伐强度为25%的样地相比,增加了25.72%、25.68%、29.64%。对照样地大径级林木生物量最大(72.57 t·hm-2),分别比间伐强度为15%的样地、间伐强度为25%的样地、间伐强度为35%的样地显著增加了79.33%、76.95%、8.53%,其中,间伐强度为35%的样地,与间伐强度为15%的样地、间伐强度为25%的样地间大径级林木生物量存在显著差异,而间伐强度为15%的样地与间伐强度为25%的样地间大径级林木生物量无显著差异。
间伐强度为25%样地的小径级林木碳密度,分别比对照样地、间伐强度为15%的样地、间伐强度为35%的样地增加64.86%、35.03%、64%,而对照样地与间伐强度为35%的样地间小径级林木碳密度无显著差异;间伐强度为15%样地的中径级林木碳密度最大(16.77 t·hm-2),分别比对照样地、间伐强度为25%的样地、间伐强度为35%的样地增加了23.91%、18.01%、16.16%;4个样地的大径级林木碳密度间均存在显著差异,间伐强度为15%样地的大径级林木碳密度最小(3.80 t·hm-2);对照样地的大径级林木碳密度最大(17.81 t·hm-2),分别比间伐强度为15%的样地、间伐强度为25%的样地、间伐强度为35%的样地增加78.66%、62.21%、24.14%。
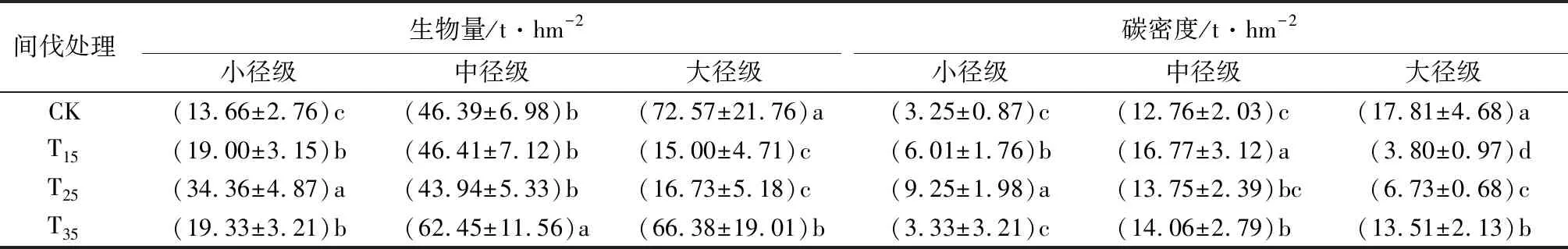
表11 不同间伐强度样地各径级生物量及碳密度
4 结论与讨论
抚育间伐移去了部分林木,直接降低了林分密度,导致林分生物量减小。间伐强度的大小直接影响被移除林木个体数量,从而影响林分密度的大小,最终导致林分生物量及碳储量的变化。林分密度大的样地,林木个体生长受限冠幅的限制,使得供给林木生长的光合同化物减少,林分生物量较小。林分密度较小的样地,林木个体生长显著增加,林分生物量随之增大。李春明等[19]、Campbell et al.[20]、Kariuki[21]认为,间伐虽能增加单木生物量,但随间伐强度增加林分生产力会降低。殷博等[2]、武朋辉等[22]、徐金良等[23]的研究结果显示,抚育间伐对乔木层的影响是在一定程度上降低碳储量;而成向荣等[24]、游伟斌等[25]研究则表明,合理的抚育间伐强度能够增加乔木层碳密度。
小兴安岭针阔混交天然次生林间伐8 a后,间伐强度为15%、25%的样地,生物量小于对照样地,说明间伐会一定程度上减少林分生物量,从而减少相应碳储量,这与殷博等[2]、武朋辉等[22]、徐金良等[23]研究结果一致。林木生物量,随间伐强度的增加呈“J”型变化,直到间伐强度为35%时林木生物量超过对照样地生物量。说明合理的间伐强度可以增加林分生物量,这与成向荣等[24]、游伟斌等[25]研究成果一致。间伐可以改变生物量在林木各器官的分配,对于树干间伐可以整体使树干生物量占比增加,对于树枝则相反。间伐虽移除部分乔木,使林分生物量降低,经过8 a时间恢复,间伐强度较大的样地,林分密度适中,林木个体生长显著增加。而间伐强度较低的样地,林分密度大,林木个体生长受有限冠幅的限制,使得供给林木生长的光合同化物减少[26],林木个体生长缓慢,最终导致生物量增加缓慢。碳密度在林木各器官的分布排列一致,由大到小依次均为树干、树枝、树叶、树皮。乔木层树干的碳密度,在间伐强度为15%、25%、35%时,随间伐强度的增大而变大。表明合理的间伐强度有利于树干累积碳储量,其主要原因是间伐改善林木生长空间,促进了保留木树高、胸径的快速生长。
间伐会直接改变林分密度,间接改变林内光照、土壤条件、水分条件等因子,从而影响林木碳质量分数[27]。以合适的强度间伐后,小兴安岭林木充分利用光照、降水等资源,使保留木生长活力得到释放,进而促进其生长,从而更好地实现固碳。不同树种碳质量分数有差异,小兴安岭中的云冷杉碳质量分数大于色木槭和青楷槭碳质量分数,是因为云冷杉稍耐荫,能耐干燥及寒冷的环境条件[28],在较适宜的生境下能固定更多的碳;而色木槭和青楷槭属阔叶落叶乔木,不耐干燥,对气候变化的适应性稍差。同一树种不同器官受主要组成成分、器官功能、器官生长年限的影响,碳质量分数也存在差异。
择伐周期、更新措施、立地条件等因素,均能影响树种组成[28]。树种组成能够影响林木更新生长,进而影响林分生物量、碳储量、碳密度。对人工林的树种组成优化比例研究成果很多[29-30],而对天然次生林的树种组成研究较少,主要原因是该问题的复杂性。目前天然林多为复层混交林,树种组成由几个到十几个不等。稳定的天然林中,这些树种的存在是它们在自然过程中相互选择、相互适应的结果,因而具有一定的规律性。而间伐会改变林分的树种组成,从而影响生物量、碳储量的分配。本研究中,间伐强度为35%样地的4色2青3云1冷树种组成,会使样地生物量增加,有利于林分更新生长。
林分生长中,径级分布是遵从一定规律变化的[31]。针阔混交林直径结构比较复杂,其分布曲线类型受林分结构、更新方式、树种特征及组成、采伐方式及强度等因素影响。本研究分析了抚育间伐8 a后的小兴安岭针阔混交天然次生林直径结构,用韦伯(Weibull)分布函数对现实林分直径结构进行了静态拟合,R2均在0.99以上,可准确描述林分直径分布。在间伐强度为35%样地中,存在大量中大径级优势树种,使得样地生物量增加,且碳密度与对照样地无显著差异。
分析结果表明,合理的抚育间伐不会导致生态系统碳储量降低。在间伐8 a后的小兴安岭针阔混交天然次生林中,间伐强度为15%会显著降低林木碳密度,但间伐强度为25%的样地、间伐强度为35%的样地,碳密度与对照样地无显著差异,且间伐强度为35%的样地生物量超过对照样地生物量,因此间伐强度35%为合理间伐强度。除了因抚育间伐移去了乔木层部分林木,使林分密度降低,碳密度因此变小这一直接原因外,还由于间伐改变了林分结构,从而改变林内光照、土壤条件、水分条件等间接因素,间接改变林分碳密度。林分碳密度也会因所选样地地理位置不同、海拔不同而有所差异。因此,仍需加强间伐对天然林碳密度长期影响的研究。
