牡丹膜联蛋白基因PsANN1的克隆及表达分析
2021-04-27徐娟娟刘鑫贺丹逯久幸栗燕
徐娟娟,刘鑫,贺丹,逯久幸,栗燕
(河南农业大学风景园林与艺术学院,河南 郑州 450002)
牡丹(Paeoniasuffruticosa)属多年生落叶灌木,其花朵硕大、花型繁多、花色艳丽,具有较高的观赏价值,迄今已有1 650多年的栽培历史[1],实现牡丹连年盆栽种植,一直是家庭养花爱好者追求的目标。然而,牡丹盆栽种植后常常出现植株矮小、花蕾败育的现象,观赏价值大大降低,这已成为盆栽牡丹产业化发展中的瓶颈问题。已有研究表明,牡丹盆栽种植后,由于根域受到限制,叶片光合速率降低,蔗糖供应不足,从而导致盆栽植株生长发育不良[2]。蔗糖在植物体内运输有共质体与质外体2种途径,共质体途径通过胞间连丝运输蔗糖,质外体途径则需要借助于蔗糖转运蛋白[3]。目前已知的蔗糖转运蛋白有SUT(sucrose transporters)、SWEET(sugars will eventually be exported transporters)和ANN(Annexins)。SUT负责将蔗糖装载至源叶的韧皮部,以及将蔗糖卸载至如贮藏根、块茎、果实、发育中的组织等库组织的细胞中[4],SWEET将细胞内的蔗糖转运至质外体[5],而ANN是新近发现的与蔗糖转运相关的蛋白。WANG等[6]首次发现,拟南芥AtANN1和AtANN2参与蔗糖从韧皮部向根尖的运输过程。在无糖条件下培养的ANN突变体幼苗,与野生型相比,根冠中含有淀粉粒的柱状细胞层数减少,周围无边界状细胞,淀粉含量减少,但这种缺陷通过添加蔗糖可以修复,表明ANN1和ANN2可能参与蔗糖从韧皮部向根尖转运。GUILHERME等[7]在‘Beauregard’和‘Tanzania’甘薯中发现了包括ANN在内的5个与根系发育和蔗糖转运有关的基因,ANN基因在这2种甘薯的叶片、不同天数的根中均有表达,说明该基因可能参与了甘薯根部蔗糖的转运。基于牡丹栽培生理与分子生物学课题组前期对PsSUT1[8]、PsSUT2[9]、PsSWEETs[10]的研究,从‘洛阳红’牡丹盛花期花瓣的转录组测序数据,筛选出一个可能调控开花相关的蔗糖转运基因ANN[11],并对其生物信息学及表达特性进行分析,为后续验证该基因功能,探寻盆栽牡丹生长发育不良的机理,改善牡丹生长发育状况提供有益参考。
1 材料与方法
1.1 试验材料
试验材料取自河南农业大学第三生活区盆栽牡丹,品种为‘凤丹’砧‘洛阳红’。以盛花期的花瓣为材料进行PsANN1基因的克隆,以盛花期的根、茎、叶、花和小风铃期(2019年3月17日)、圆桃期(2019年3月27日)、盛花期(2019年4月9日)、谢花期(2019年4月15日)的花瓣为材料进行PsANN1基因的表达分析。
1.2 试验方法
1.2.1 总RNA的提取与cDNA的合成 参照李永华改良CTAB法[12]提取‘凤丹’砧‘洛阳红’盛花期根、茎、叶、花和小风铃期、圆桃期、盛花期、谢花期的花瓣总RNA。检测合格后使用PrimeScriptTMII 1st Strand cDNA Synthesis Kit反转录试剂盒(宝生物工程(大连)有限公司)合成cDNA第一条链,-20 ℃保存备用。
1.2.2PsANN1基因的克隆及鉴定 据实验室前期转录组测序获得的序列利用 Primer 5.0软件设计一对特异性扩增引物PsANN1-F:5’-ATGGCTACAATCATTGCCCCTC-3’和PsANN1-R:5’- TTAGTCCCCAAGCAAAGTGAGG-3’,送至郑州尚亚生物技术有限公司合成。以1.2.1方法反转录获得的盛花期花瓣cDNA为模板,使用1.1×T3 Super PCR Mix(擎科生物科技(郑州)有限公司)进行PCR扩增。扩增体系为25 μL:cDNA (100 ng·μL-1) 1 μL,1.1×T3 Super PCR Mix 22 μL,上、下游引物各1 μL。扩增程序:98 ℃预变性2 min;98 ℃变性10 s,58 ℃退火10 s,72 ℃延伸10 s,共 35个循环;72 ℃继续延伸2 min。PCR产物经体积分数为1%琼脂糖凝胶电泳后,使用DNA纯化回收试剂盒(天根生化科技(北京)有限公司)对目的片段进行回收与纯化,并将纯化后的目的片段与pMD18-T克隆载体连接(宝日医生物技术(北京)有限公司),并转化DH5α感受态细胞(宝日医生物技术(北京)有限公司),PCR筛选阳性菌株,送至郑州擎科生物公司测序。
1.2.3PsANN1基因生物信息学分析、进化树构建 利用DNAMAN软件把克隆得到的核苷酸序列通过推导得到氨基酸序列。使用NCBI基于氨基酸序列进行蛋白质的保守结构域分析(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi),并筛选同源序列(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome)。使用ProtScale在线分析PsANN1蛋白的理化性质,预测其亲水性和疏水性(https://web.expasy.org/protscale/)。利用SOPMA预测蛋白质二级结构(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html),利用SWISS-MODEL预测蛋白质的三级结构(https://swissmodel.expasy.org/interactive)。使用 DNAMAN对氨基酸序列进行多序列比较,利用在线软件MEME进行氨基酸保守基序分析(基序最大发现值设为5,其他参数均设为默认值)(http://meme.nbcr.net/meme/cgi-bin/meme.cgi),并利用 MEGA7.0 (邻接法)构建系统进化树。
1.2.4PsANN1基因表达 根据克隆测得的PsANN1序列设计qRT-PCR特异性引物(qPCR-PsANN1-F:5’-GGAGCCTTGATTACCGAGTG-3’,qPCR-PsANN1-R:5’-CAATGCCTTGTCCTTGATAG-3’),并以牡丹泛素延伸蛋白Ubiquitin(Ubiq)基因为内参[13](Ubiq-F:5’-AGCCCATATTCAGGAGGTGT-3’,Ubiq-R:5’-GCAATCTCAGGTACAAGGGG-3’),以1.2.1中得到的‘凤丹’砧‘洛阳红’盛花期的根、茎、叶、花,以及小风铃期、圆桃期、盛花期、谢花期的花瓣的cDNA为模板,使用TB GreenTMPremix Ex TaqTMII (Tli RNaseH Plus)(宝生物工程(大连)有限公司),在荧光定量分析仪QuantStudio 5(美国Thermo Fisher Scientific公司)上进行PsANN1基因在组织特异性和不同花期花瓣中表达水平的测定。扩增体系20 μL:cDNA (100 ng·μL-1) 2 μL,TB Green Premix Ex Taq II(Tli RNaseH Plus)(2×)10 μL,ROX Reference Dye II(50×)0.4 μL,上、下游引物各0.8 μL,ddH2O补足至20 μL。所有处理均设3次重复。PCR扩增程序为:95 ℃预变性10 min;95 ℃变性15 s,60 ℃延伸1 min,共35个循环;95 ℃ 变性15 s,60 ℃ 1 min,95 ℃变性1 s。使用 2-ΔΔCt方法和SPSS 19.0软件对试验结果进行分析。
2 结果与分析
2.1 牡丹PsANN1基因的克隆及推导的氨基酸序列
根据转录组数据库中已知ANN基因序列设计特异引物,以盛花期花瓣cDNA为模板进行PCR扩增。PCR产物经体积分数1%琼脂糖凝胶电泳,检测到特异扩增条带(图1)。测序结果显示,PCR产物长度为939 bp,编码312个氨基酸(图2)。DNAMAN软件分析结果表明,此扩增序列包含完整开放阅读框(open reading frame,ORF),将该基因命名为PsANN1,GeneBank登录号为MK550694。

注:M:DL 2 000标记;泳道1:无样品;泳道2:为PsANN1基因扩增产物。Note:M:DL 2 000 DNA Marker;Lane 1:No sample;Lane 2:PsANN1 gene amplification product.图1 PsANN1基因RT-PCR扩增Fig.1 RT-PCR amplification of PsANN1 gene

图2 PsANN1基因的ORF序列与推导的氨基酸序列Fig.2 ORF sequence and deduced amino acid sequence of PsANN1 gene
2.2 牡丹PsANN1基因编码蛋白的理化性质
利用NCBI中的ExPASy ProtPra在线软件对PsANN1基因编码的蛋白质进行理化性质分析。分析表明,该蛋白分子式为C1566H2550N456O474S6,相对分子质量35.54 kD,等电点8.20。该蛋白中Ala(10.6%)和Leu(10.3%)含量相对较多,Met(0.6%)和Trp(0.6%)含量相对较少。总的带负电荷残基Asp+Glu数目为48,带正电荷残基Arg+Lys 数目为50。由图3可知,分值为负的氨基酸明显多于分值为正的氨基酸,且平均亲水系数-0.395,因此可推测该蛋白为亲水性蛋白。
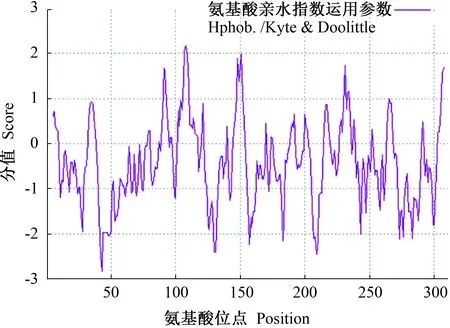
图3 PsANN1蛋白的疏水性预测Fig.3 Hydrophobicity prediction of PsANN1 protein
2.3 牡丹PsANN1蛋白二级结构、三级结构及功能结构域的预测
利用SOPMA对PsANN1蛋白进行二级结构的预测,通过预测结果可知,该蛋白主要由 α-螺旋(70.83%)、无规卷曲(20.19%)、扩展链(6.73%)、β-转角(2.24%)组成(图4)。使用SWISS-MODEL对PsANN1蛋白的三级结构进行预测(图5),PsANN1蛋白与黄麻(Corchoruscapsularis)、野草莓、月季的ANN蛋白三级结构高度相似。通过NCBI软件分析该蛋白的功能结构域(图6),结果表明,该蛋白包含4个重复的 Annexin 保守结构域,其中第2和第3个基本结构单位有一定的缺失和突变。

图4 PsANN1蛋白二级结构预测Fig.4 Prediction of the secondary structure of PsANN1 protein
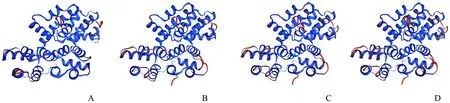
注:A.牡丹;B.黄麻;C.野草莓;D.月季。Note:A.Paeonia suffruticosa;B.Corchorus capsularis;C.Fragaria vesca subsp.vesca;D.Rosa chinensis.图5 PsANN1蛋白及其同源蛋白三级结构预测Fig.5 Tertiary structure analysis of PsANN1 and other homologous proteins

图6 PsANN1基因ORF序列的功能结构域Fig.6 The functional domain of the ORF sequence of PsANN1 gene
2.4 牡丹PsANN1基因的氨基酸序列比对及系统进化树构建
使用DNAMAN软件将PsANN1编码氨基酸的序列,与其他植物ANN基因的氨基酸序列进行同源性比对。结果显示,牡丹PsANN1编码的氨基酸序列与黄麻(OMP03638.1)、野草莓(XP_004307132.1)、可可(Theobromacacao.XP_017984838.1)、月季(XP_024176879.1)、石榴(Punicagranatum.OWM90035.1)、欧洲栓皮栎(Quercussuber.XP_023879336.1)、榴莲(Duriozibethinus.XP_022764810.1)的同源性分别为78.14%、77.17%、76.85%、75.88%、75.56%、74.92%、74.60%。
通过MEME在线软件对以上植物ANN基因的氨基酸序列进行分析,结果表明,共鉴定出5个相同的保守区域(图7),其中保守区域1内存在annexin repeat Ⅰ,保守区域3内存在annexin repeat Ⅱ,保守区域4内存在annexin repeat Ⅲ,保守区域5内存在annexin repeat Ⅳ,annexin repeat Ⅰ~Ⅳ是膜联蛋白基因典型的4个重复保守结构域,由此可确定,从‘凤丹’砧‘洛阳红’牡丹中克隆获得的序列为ANN基因。
为了进一步研究PsANN1蛋白与其他ANN类蛋白的亲缘关系,构建了系统进化树(图8-A),结果表明,在这8个ANN的进化关系中,牡丹与月季和野草莓亲缘关系较近(图8-B)。锦葵科(Malvaceae)的榴莲(Duriozibethinus)与可可(Theobromacacao)的ANN基因聚为一束,蔷薇科(Rosaceae)的月季与野草莓的ANN基因聚为一束,这与他们之间的亲缘关系较近有关。
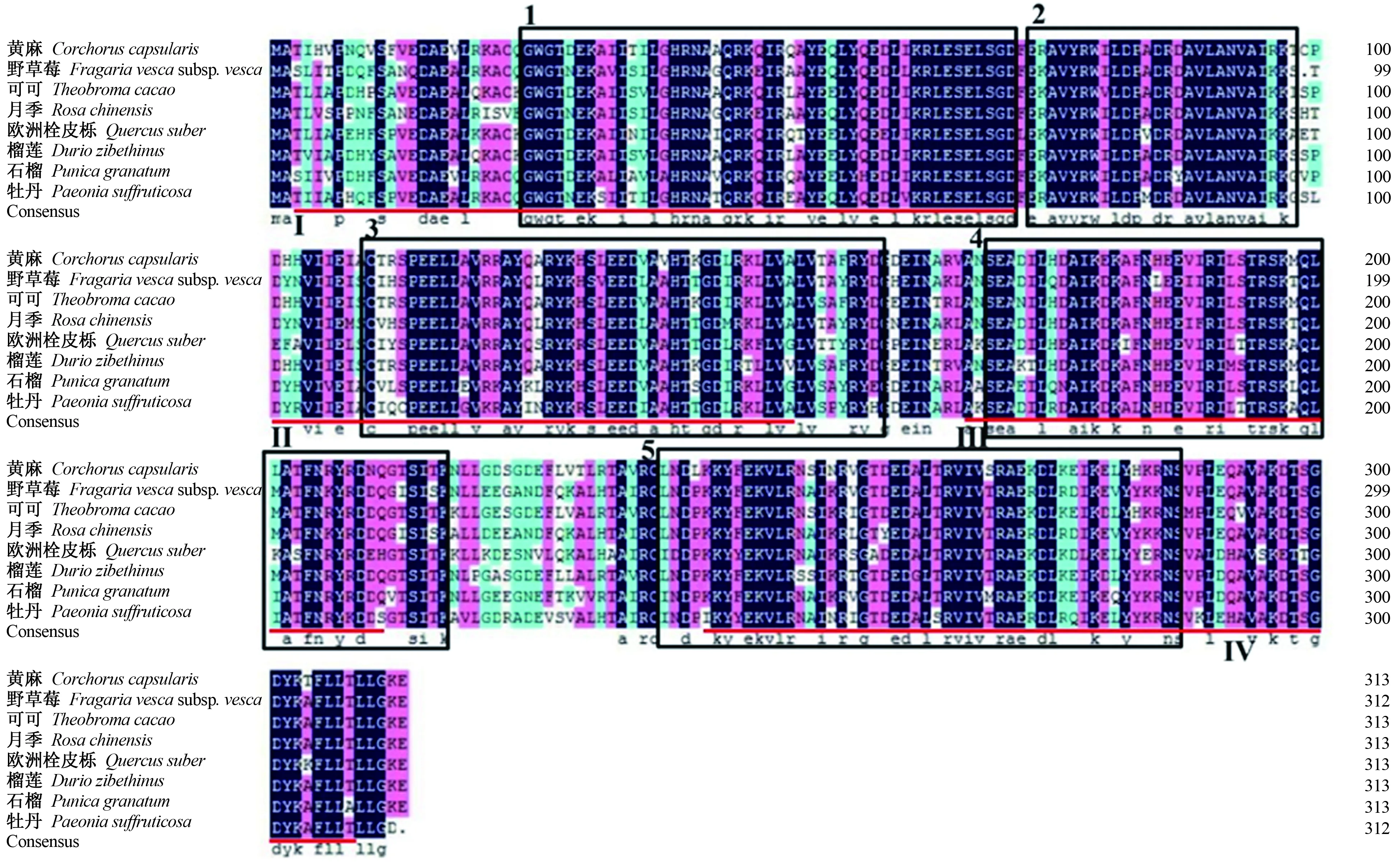
注:方框1、2、3、4、5为保守区域;红色下划线部分Ⅰ、Ⅱ、Ⅲ、Ⅳ为4个annexin repeats。Note:The box1,2,3,4,and 5 are the conserved domain;the red underlined parts Ⅰ,Ⅱ,Ⅲ,and Ⅳ are 4 annexin repeats.图7 牡丹PsANN1氨基酸与其他植物ANN氨基酸序列比对及保守区域。Fig.7 Amino acid sequence alignment and conserved regions of PsANN1 amino acid of peony.and ANN amino acid of other plants
2.5 牡丹PsANN1基因在不同组织及不同花期花瓣中的相对表达量分析
对PsANN1在盛花期‘洛阳红’牡丹不同组织部位样品进行qRT-PCR检测,结果表明,以根部的表达量为参照,PsANN1在花瓣中的表达量最高,是根部表达量的24.95倍,差异达显著水平,在茎中的表达量为根的3.46倍,在叶片的表达量极低,仅为根的0.003倍,差异达显著水平(图9-A)。以PsANN1基因在‘洛阳红’牡丹小风铃期花瓣中的表达量为参照,圆桃期、盛花期和谢花期的PsANN1基因表达量分别是小风铃期的26.66倍、149.47倍和9.26倍,差异均达显著水平,且表达量从小风铃期至盛花期迅速增加,盛花期的表达量最高(图9-B)。

注:A:系统进化树;B:氨基酸序列保守结构域。Note:A:Phylogenetic tree;B:Conserved domain of amino acid sequence.图8 植物PsANN1基因系统进化树Fig.8 Phylogenetic tree of plant PsANN1 gene

图9 PsANN1基因在‘洛阳红’牡丹不同组织部位和不同花期花瓣中的相对表达量Fig.9 Relative expression of PsANN1 gene in different tissue parts of ‘Luoyanghong’ tree peony and petals of different flowering stages
3 结论与讨论
自1989年首次从番茄(Lycopersiconesculentum)中克隆出ANN基因以来[14],陆续从蒺藜苜蓿(Medicagotruncatula)[15]、普通小麦(Triticumaestivum)[16]、玉米(Zeamays)[17]等多种植物中分离出来。根据FENG等[18]、JAMI等[19]、HOSHINO等[20]的研究,植物膜联蛋白的相对分子质量通常在32~42 kD之间。本试验从‘凤丹’砧盛花期‘洛阳红’牡丹花瓣中克隆得到了膜联蛋白基因PsANN1,其CDS区长939 bp,编码312个氨基酸,相对分子质量为35.54 kD。将PsANN1编码的氨基酸序列与其他植物ANN基因的氨基酸序列进行同源性比对以及保守结构域分析,发现PsANN1基因编码的氨基酸序列与黄麻同源性最高,达78.14%,膜联蛋白4个重复保守结构域存在于鉴定出的5个相同保守区域中。对PsANN1蛋白进行二级结构、三级结构和功能结构域的预测,可知该蛋白主要由 α-螺旋组成,与黄麻、野草莓、月季的ANN蛋白的三级结构高度相似,包含4个重复的 Annexin 保守结构域,其中第2和第3个基本结构单位有一定的缺失和突变,这一结果与花生(Arachishypogaea)[21]、芥菜(Brassicajuncea)[22]中克隆得到的膜联蛋白结构域一致,符合植物膜联蛋白的进化特点。
已有研究表明,ANN基因在不同植物的不同组织均有表达,但表达情况不尽相同。花生AhAnn1在根、茎、叶中均有表达,茎的表达量最高,根的表达量最低[21];苦荞(Fagopyrumtataricum)FtANX1在根、茎、叶、花及未成熟的果实中均有表达,在未成熟的果实中表达量最多,在叶中的表达量最少[23];拟南芥(Arabidopsisthaliana)AtANN1在所有部位均有表达且在茎中表达量最高[24]。本研究中qRT-PCR结果显示,PsANN1基因在‘洛阳红’牡丹根、茎、叶、花中均有表达,但在花中表达量最多,茎中次之,叶片中最少,表明PsANN1基因主要在‘洛阳红’牡丹的花器官(库器官)中发挥重要作用,这与苦荞中ANN基因的表达量结果相似,与花生ANN基因的表达量结果不一致,可能与物种间的遗传差异有关。ANN基因在同一植物组织部位的不同发育时期的也有表达,棉花(Gossypiumhirsutum)GhAnn3、GhAnn4和GhAnn5基因主要在棉纤维伸长期表达[25];GhFAnn1和GhFAnnx在纤维发育的不同时期均有表达,且表达量均很高[26];草莓(Fragaria×ananassa)ANN基因随着果实发育成熟,表达量逐渐增加[27]。PsANN1基因在开花的不同时期中,随着花瓣的伸长与增大,自小风铃期至盛花期表达量迅速增加,盛花期表达量最多,推测PsANN1基因在‘洛阳红’牡丹从小风铃期到盛花期的开花转变中具有重要调节作用。为进一步验证该推测,还需采用基因敲除或构建超表达载体等手段,转化模式植物,进一步验证该基因的功能。
