亚洲棉CBL基因家族鉴定及生物信息学分析*
2021-04-27杨秀邓艳凤肖水平刘新稳王涛杨绍群
杨秀,邓艳凤,肖水平,刘新稳,王涛,杨绍群
(江西省棉花研究所/国家棉花产业技术体系鄱阳湖综合试验站,江西 九江 332105 )
土壤盐碱化会引起离子毒性、高渗透胁迫和氧化等次生胁迫从而对植物造成危害[1]。但植物为了应对恶劣的环境在高盐条件下进化出很多调节植物生长的机制[2]。其中一个关键机制涉及到植物细胞中的钙浓度升高和钙依赖信号通路[3]。研究发现,钙调磷酸酶B亚基蛋白(calcineurin B-like proteins)基因在植物非生物逆境应答以及钙信号转导过程发挥重要的作用。CBL蛋白包含EF-hand基序,与Ca2+结合时,它们发生构象变化,并与丝氨酸/苏氨酸蛋白激酶(CIPK)特异性相互作用,形成CBL-CIPK通路,被广泛认为是植物响应和适应不同外部压力触发Ca2+信号的主要机制[4-6]。此外,CBL-CIPK已被证明是调节植物对多种非生物胁迫(如高盐、K+缺乏、Mg2+过量和干旱)的重要信号网络[7-8]。它还调节植物的生长和发育,硝酸盐、铵和铁的吸收和运输,维持H+的稳态,以及活性氧信号的转导。
到目前为止,关于CBL蛋白功能的研究很多,拟南芥中的研究最为深入。在拟南芥中,已经发现了编码CBL蛋白的10个基因[4]。其中钙调磷酸酶B亚基蛋白4(CBL4/SOS3)、CBL相互作用蛋白激酶24(CIPK24/SOS2)和SOS1组成的SOS途径是通过将Na+挤压到细胞外空间,从而使植物具有耐盐性,控制细胞离子平衡[9-10]。CBL10-CIPK8复合物通过激活拟南芥幼苗中SOS1途径,使Na+外排,从而提高拟南芥的耐盐能力[11]。过表达拟南芥CBL5使植物在发育早期对高盐或高渗透胁迫具有更强的抵抗力,但没有改变它们对ABA的反应[12]。此外,CBL2和CBL3还参与调控植物种子产量和大小,并且与CIPK3/9/23/26构成了一个多价的相互作用网络,调控Mg2+的空泡隔离,从而保护植物免受Mg2+毒性[13-15]。AtCBL1还在盐和干旱环境下发挥正向调节作用,而在冷胁迫反应中发挥负向调节作用[16]。CBL9还参与植物对ABA、K+缺失的响应过程,并通过减少胞内游离钙浓度负调控耐寒性[17-19]。拟南芥CBL7还参与调控植物对低硝酸盐的反应[20]。目前,小麦[21]、水稻[22]、大白菜[23]、杜梨[24]、番茄[25]、陆地棉[26]等植物在全基因组水平上检测到多个CBL基因家族,且CBL蛋白含有EF-hands、myristoylation和palmitoylation sites等保守结构域[4, 27],并且研究了许多CBL基因在不同组织中的表达模式,以及在植物对各种非生物胁迫的响应[27, 28]。这些研究成果为人们进一步探索CBLs在植物中的作用机制奠定了基础。然而,关于亚洲棉CBLs的基因组学和进化研究还很少。因此,作者利用生物信息学的方法,在亚洲棉基因组序列的基础上,以CBL家族蛋白质结构域为搜索条件,鉴定亚洲棉中CBL基因,对CBL基因成员的理化性质、进化关系、基因结构、蛋白结构、染色体定位、顺式作用元件进行分析,为进一步探索棉花CBL基因的功能提供线索。
1 材料与方法
1.1 亚洲棉CBL家族成员鉴定与理化性质分析
从phytozom (https://phytozome.jgi.doe.gov/pz/)下载水稻的CBL蛋白,而拟南芥的CBL蛋白从TAIR(http://www.arabidopsis.org/)下载。利用拟南芥和水稻CBL家族蛋白序列,在CottonGen数据库(https://www.cottongen.org/)中比对亚洲棉CBL蛋白序列,人工去除冗余蛋白序列,利用SMART (http://smart.embl-heidelberg.de/)、CDD(http://www.ncbi.nlm.nih.gov/cdd)和PFAM(http://pfam.sanger.ac.uk/) 预测并分析上述候选蛋白序列的保守结构域,剔除不含完整保守结构域的序列,同时,利用COTTON FGD (http://www.cottonfgd.org/)数据库对亚洲棉CBL基因的染色体位置及其编码产物的相对分子质量和理论等电点等基本信息进行分析。
1.2 亚洲棉CBL基因家族系统进化、基因结构和染色体定位分析
利用Clustal X 2.0(http://www.clustal.org/clustal2/)程序对获得的拟南芥、水稻、亚洲棉的CBL蛋白序列进行多序列比对[29],利用MEGA 6.0软件生成.meg格式文件,然后采用Neighbor-Joining法构建系统进化树,其Bootstrap method值设为1000,其余参数设为默认值。利用MapChart 2.2[30]软件对亚洲棉CBL家族基因进行染色体定位。GaCBL基因家族成员的外显子-内含子结构通过基因结构分析软件分析(Gene Structure Display Server, GSDS 2.0)[31]。
1.3 亚洲棉CBL蛋白基序和保守结构域分析
CBL蛋白序列的保守基序分析采用MEME在线软件预测,参数设置如下:“Maximum number of motifs”为15,“Occurrences of a single motif”为“zero or one per sequence”,采用CDD(https://www. ncbi.nlm.nih.gov/cdd/)数据库查找CBL的保守结构域信息,利用TBtool作图工具绘制保守结构域。
1.4 亚洲棉CBL家族基因的顺式作用元件分析
利用棉花基因组数据库Cotton FGD(http://www.cottonfgd.org/)提取陆地棉CBL基因序列转录起始位点ATG上游1500 bp序列,然后将获得的序列提交到Plant CARE(http://www.dna.affrc.go.jp/PLACE/signalscan.html)数据库中,鉴定启动子区域可能存在的顺式作用元件。
2 结果与分析
2.1 亚洲棉CBL家族成员的鉴定与理化性质
利用BLASTP和HMMER搜索获得候选氨基酸序列,并用Pfam、SMART和CDD工具筛选候选氨基酸序列,剔除不含EF-hand结构域的冗余序列,最终获得20个亚洲棉CBL基因。根据CBL基因所在的染色体位置和前人的研究,并以GaCBL1-1形式将其命名,所有基因家族成员的基因名字和理化性质如表1。研究分析发现亚洲棉CBL基因家族成员的编码基因组序列差异较大(241~11123 bp),而编码序列长度差异较小(199~786 bp),这可能与内含子-外显子结构的不同有紧密的关系。从表1中还可以看出,基因编码产物的理论等电点(pI)、相对分子质量大小差异不大,表明棉花CBL蛋白的理化性质差异不大;除GaCBL4-3(等电点为8.241)和GaCBL2-2(等电点为6.241)以外,其余CBL蛋白的等电点为4~5.5,表明CBL蛋白中的氨基酸大部分为酸性。在亚洲棉CBL的家族成员中,除GaCBL2-3、GaCBL2-4、GaCBL2-5、GaCBL2-6、GaCBL3-2、GaCBL4-2和GaCBL9的总平均疏水性得分为阳性外,其余GaCBL得分为阴性,说明大多数GaCBL蛋白为亲水性蛋白。
2.2 亚洲棉CBL基因家族的系统进化分析
为了确定CBL蛋白的系统发育关系,采用邻近法(NJ),将完整的CBL蛋白序列用MEGA6.0进行比对,构建系统进化树(图1)。系统进化树将亚洲棉的CBL蛋白与拟南芥、水稻的CBL蛋白分成了两个组,其中Group II包含的成员最多,Group I中仅有GaCBL4-1、GaCBL4-2、GaCBL4-3和GaCBL8共4个成员。Group I包含的成员数量虽少,但在不同的物种中均存在,说明该组成员可能参与了植物体的某种生物学过程。
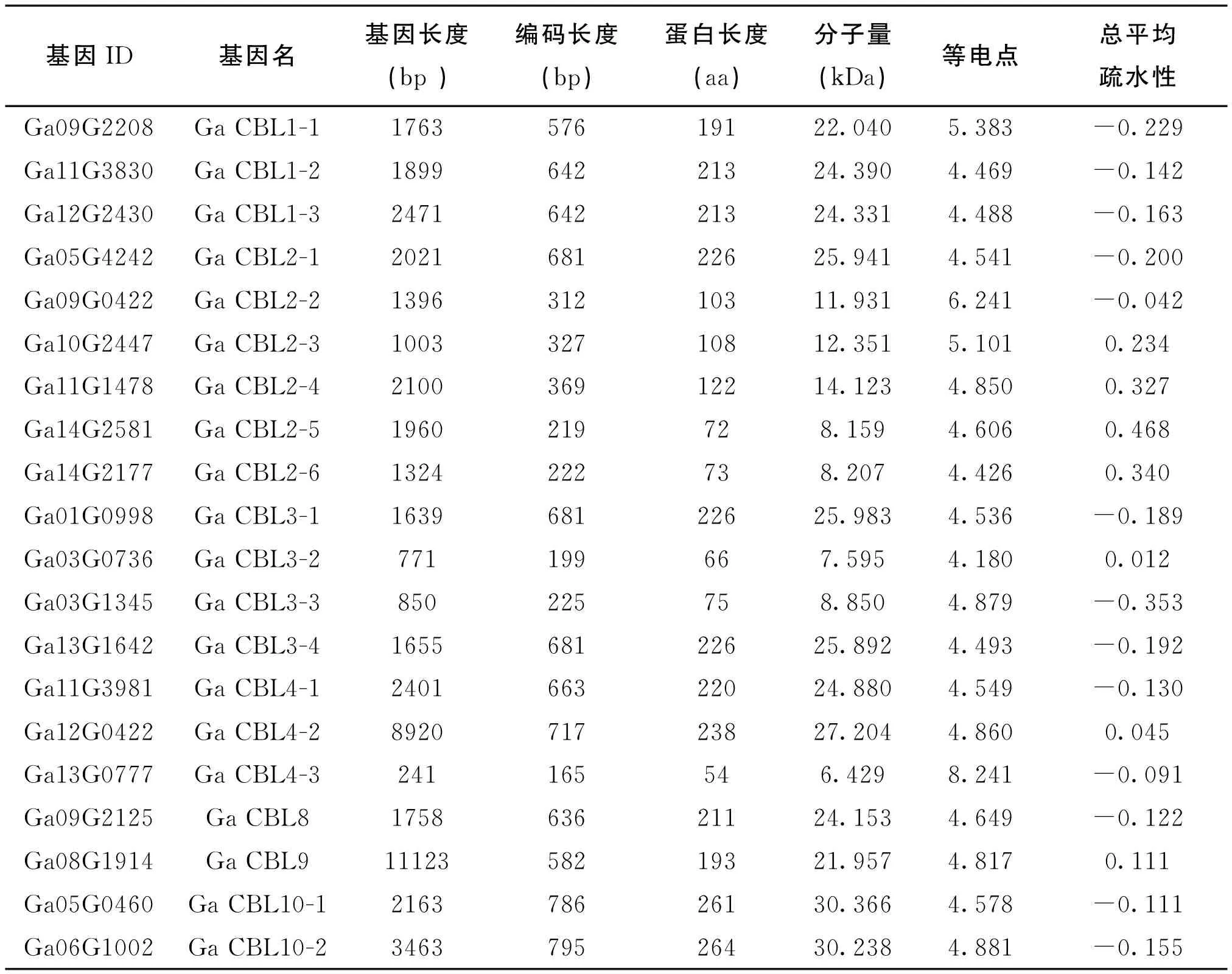
表1 亚洲棉CBL家族基因的理化性质
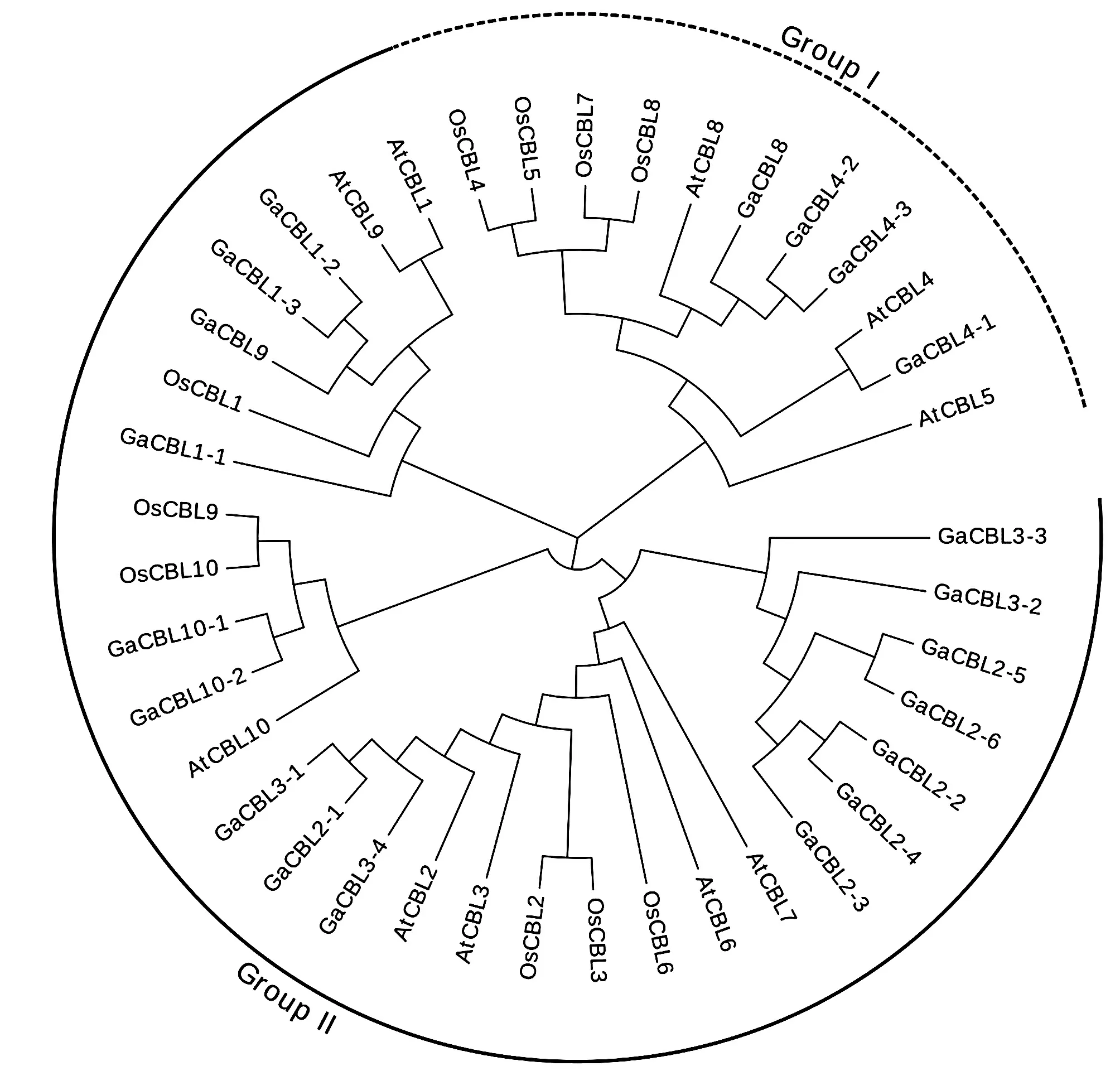
图1 拟南芥、水稻、亚洲棉CBL基因家族系统进化树
2.3 亚洲棉CBL基因家族成员基因和蛋白结构分析
在基因结构分析中,大部分亚洲棉CBL基因结构被内含子破坏。在不同的基因中被破坏程度不同,被内含子高度打断的基因包括GaCBL8、GaCBL4-1、GaCBL4-2、GaCBL10-1、GaCBL10-2、GaCBL1-2、GaCBL1-3、GaCBL9、GaCBL1-1、GaCBL3-1、GaCBL2-1和GaCBL1-1,占超过55%的亚洲棉CBL基因。其余的CBL基因最多有4个内含子。此外,根据基因结构将CBL基因家族成员分为2组,这与系统进化树的分组高度一致。同一组内各基因中的外显子-内含子结构比较相似,而不同组的基因结构差异较大。这表明基因结构与它们的遗传进化关系密切。
通过结构域分析发现所有的CBL基因均含有至少一个EF-hand结构域,且除了该结构域以外,不含有其他结构域。保守基序分析发现,同一类群中的大多数成员具有相似的motif,说明同一类群中的基因存在着功能上的相似性。
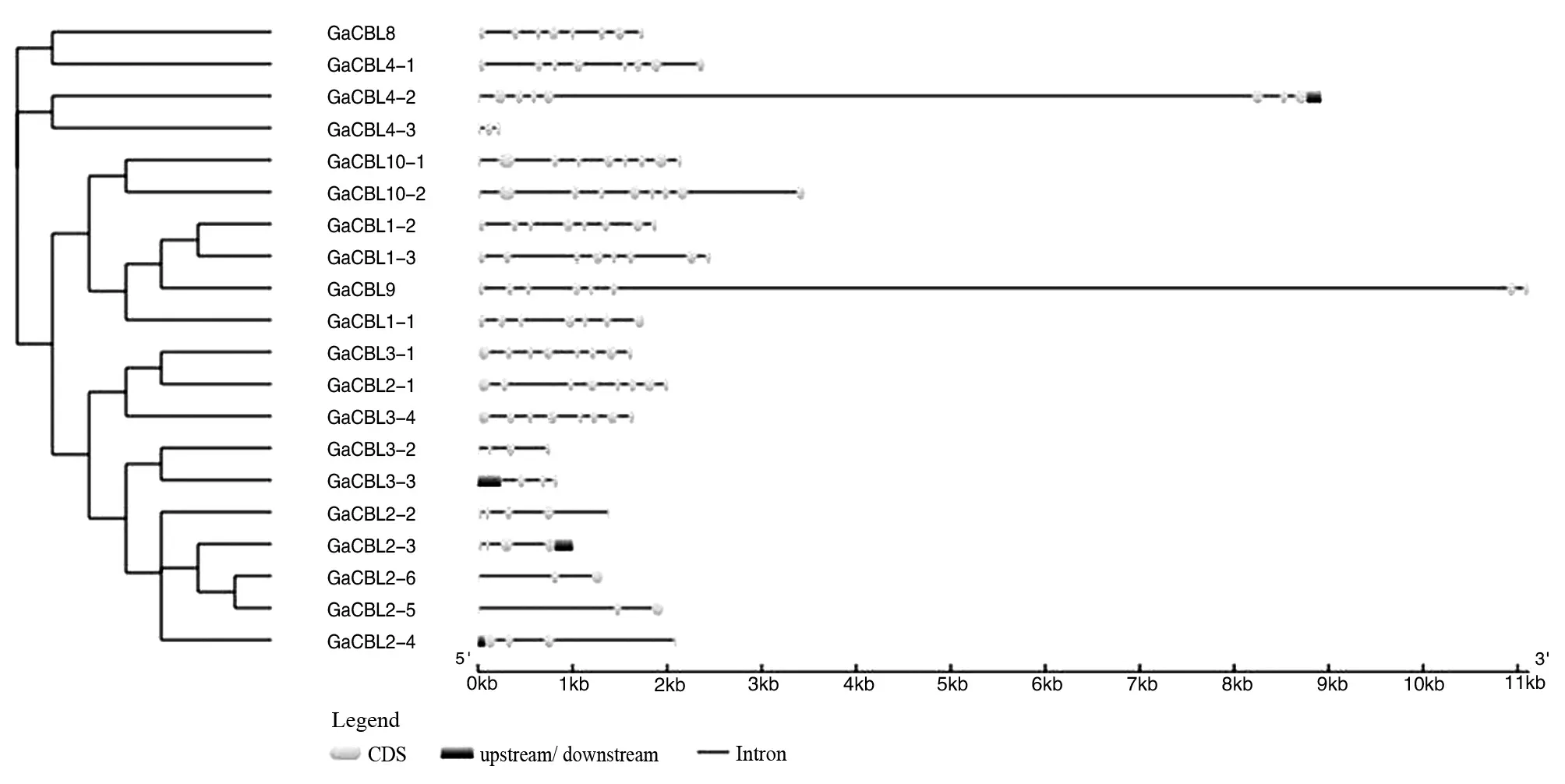
图2 亚洲棉CBL家族基因结构分析

图3 亚洲棉CBL家族基因蛋白结构分析
2.4 亚洲棉CBL基因家族染色体定位
为了确定CBL家族基因在染色体上的分布,对CBL基因进行染色体定位(图4)。结果表明18个CBL基因被定位在10条染色体上,而GaCBL2-5和GaCBL2-6显然不能定位到任何染色体上。且基因在染色体上的位置相对不均匀,在9号和11号染色体的下端处分布较密。
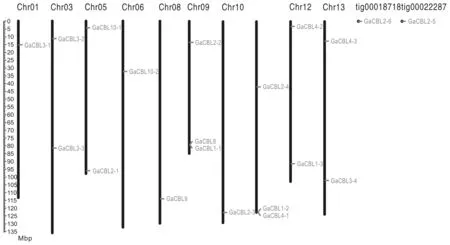
图4 亚洲棉CBL家族基因在染色体上的位置
2.5 顺式作用元件分析
对GaCBL基因的启动子区域上游的1500 bp序列进行顺式作用元件分析,我们发现GaCBL家族基因成员启动子区域中均含有多个能够应答逆境和植物激素的顺式作用元件,包括ABA反应元件(ABRES)、低温应答元件(LTRS)、病原物防御和应激反应元件(TC-rich repeats)、茉莉酸甲酯应答元件(CGTCA-motif 和TGACG-motif)、水杨酸应答元件(TCA-element)、干旱诱导元件(MBS)(表2),这说明亚洲棉CBL基因可能参与多种非生物胁迫和激素应答过程,具有复杂的生物学功能。其中GaCBL1-2不仅含有脱落酸应答元件(ABRES)、干旱应答元件(MBS)和低温应答元件(LTRS)等非生物逆境应答元件,还含有病原物防御和应激元件(TC-rich repeats)等,推测该基因可能参与ABA、低温及各种生物胁迫应答。从表2中还可以发现茉莉酸甲酯应答元件(TGACG-motif)是CBL基因启动子中最丰富的顺式调节激素应答元件,其中15个基因成员包含TGACG-motif,说明大多数的亚洲棉CBL基因参与茉莉酸甲酯的应答过程。
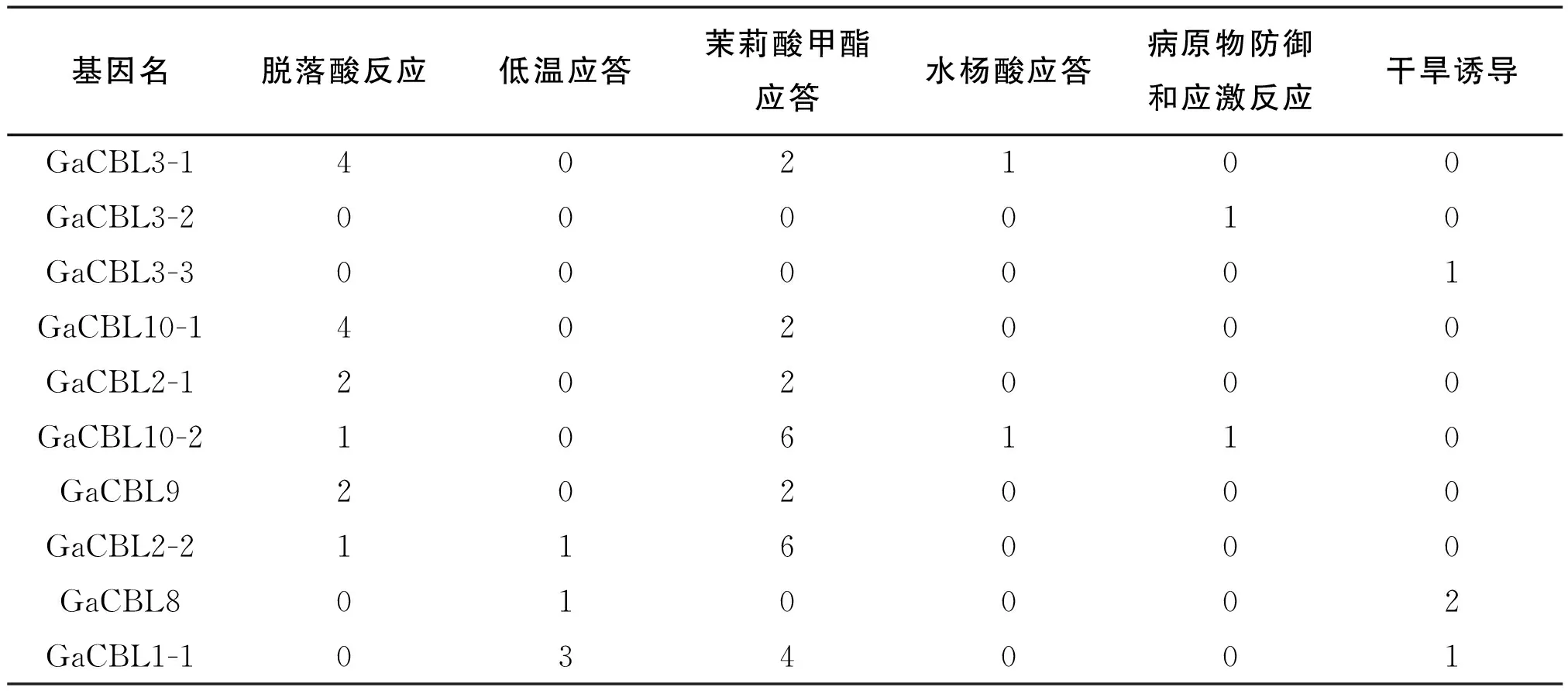
表2 亚洲棉CBL基因顺式作用元件分析
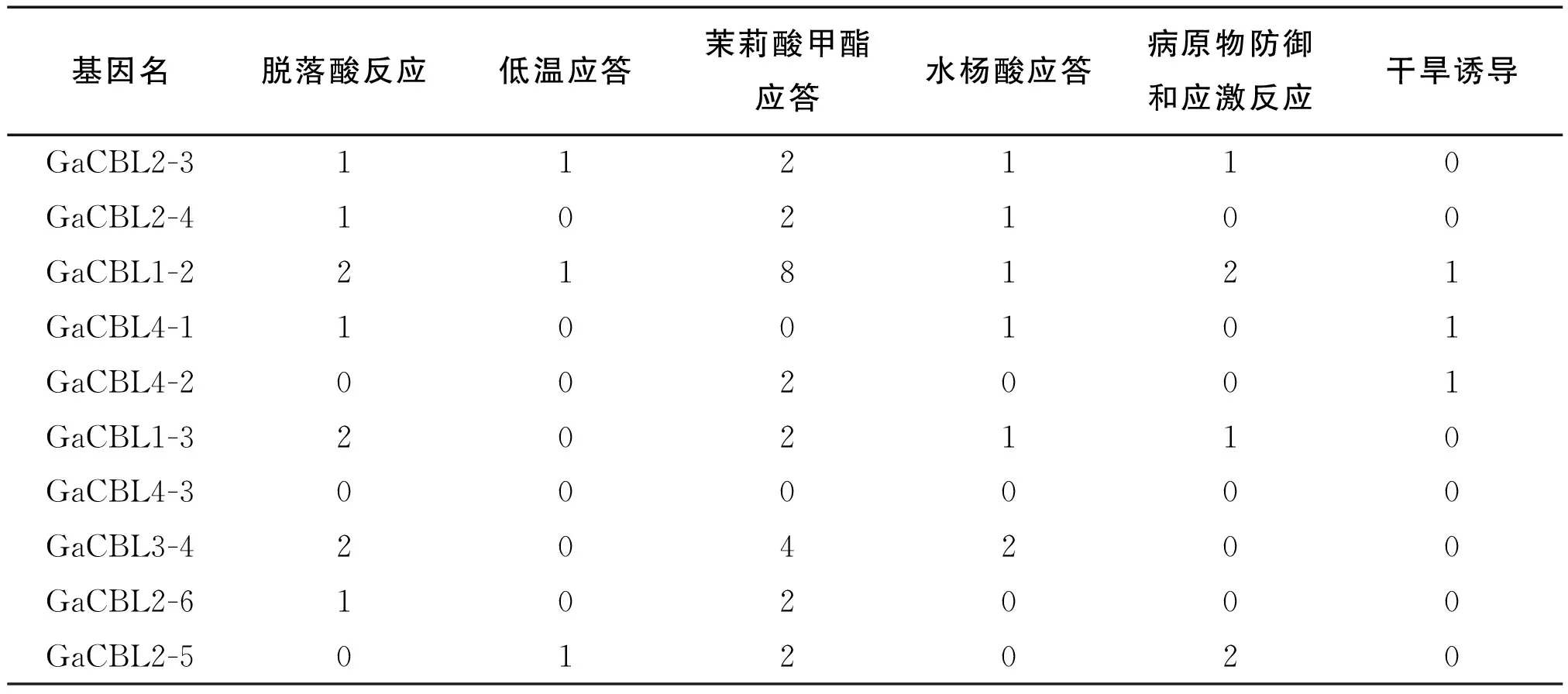
续表2 亚洲棉CBL基因顺式作用元件分析
3 讨论与结论
目前关于棉花CBL基因的研究较少,关于亚洲棉CBL家族基因的研究鲜见报道。本研究通过生物信息学分析,从亚洲棉基因组中鉴定出20个CBL基因,并对这些基因的系统进化关系、基因和蛋白结构、染色体分布和顺式作用元件进行分析。结果表明,亚洲棉CBL家族基因的理化性质差异不大,且大多数GaCBL蛋白为亲水性蛋白;系统进化树分析得出两个组,Group II包含的成员最多,Group I中仅有GaCBL4-1、GaCBL4-2、GaCBL4-3和GaCBL8共4个成员。外显子-内含子结构分析表明,大部分CBL基因有8~9个外显子,且在同一组的基因中,外显子-内含子结构比较相似,且组之间的基因结构差异较大。编码区分析表明CBL基因均含有至少一个EF-hand结构域,且不含有其他结构域。染色体定位分析表明,20个CBL基因在基因组中的分布是不均匀的,9号和11号染色体上分布最多,在2号、4号、7号染色体上未发现。顺式作用元件分析表明,所有的亚洲棉CBL基因都含有几个或者多个应答不同逆境和植物激素的顺式作用元件,且含有的顺式作用元件的类型和数量各不相同,这说明亚洲棉不同CBL成员可能参与不同的生物学过程并发挥着不同的功能。这些结果可以为进一步克隆和研究亚洲棉CBL的功能提供参考。
