光照和氮磷供应比对木荷生长及化学计量特征的影响
2021-04-25虞木奎成向荣邹汉鲁
熊 静,虞木奎,成向荣,*,汪 成,邹汉鲁
1 中国林业科学研究院亚热带林业研究所华东沿海防护林生态系统国家定位观测研究站, 杭州 311400 2 南京林业大学, 南京 210037 3 江西省永丰县官山林场, 吉安 331500
亚热带低山丘陵区是我国生态安全战略格局“长江经济带建设”和“两屏三带”的重要组成部分,是中国木材战略储备林和经济林产品的重要基地[1]。该区域人工林以纯林为主,普遍存在林分结构单一、生产效益低、林分生态功能和系统稳定性较差等问题[2]。研究表明,构建异龄复层林是改善人工纯林林分结构,提高林分综合效益的有效措施[3-4]。然而,现存人工林内,由于林内光照条件不足、土壤质地酸化和养分失衡等因素严重影响了林下阔叶树种的天然更新和生长发育[5]。首先,光照异质是制约植物生长和分布的首要因子,在林缘、林窗和林下光照强度具有显著差异[6],植物对异质光生境的适应是物种定植、生长的基础,良好的适应能力对森林物种的竞争力非常重要[7]。其次,南方典型红壤丘陵区土壤普遍缺N少P[8]。在全球N、P沉降和农林施肥大背景下,外源N、P输入带来的元素比例失衡对生态系统的影响越来越大。N、P元素作为关键限制元素,其供应量或供应比例的改变都会严重影响植物的生长发育[9]。
生态化学计量学是研究生态系统中多重化学元素(主要是C、N、P等)平衡的科学[10],也是研究生态系统植物营养元素分配情况、利用效率和限制性元素判断的一种方法[11],能够反映植物器官不同元素的内稳性及相互关系[12],有助于解决植物和生态系统养分供应与需求平衡等方面的问题[13]。此外,植物较强的C、N、P元素可塑性对环境变化的响应可直接影响植物的生长和适应能力[14],与植物生长和功能发挥等紧密相关[15]。研究表明光照变化和树种差异均对耐荫植物在异质光环境中的N、P利用策略和分配格局产生调节,进而影响总生物量和养分积累[16-17]。此外,养分添加可直接影响植物对养分的吸收和利用,改变植物化学计量甚至植物整体生物量分配和生态策略[18-19]。然而,目前研究多集中光照或养分添加的研究,光照和养分互作对林木化学计量及生长的影响认识较为有限。因此,针对南方典型红壤丘陵区异龄复层林的发展存在的典型光照和养分失衡问题,探究伴生树种在不同光照和养分等因素调控下的化学计量和生长性状变化及适应特征,对促进异龄复层林高效培育具有重要意义。
木荷(Schimasuperba)属山茶科木荷属常绿大乔木,是我国重要的珍贵阔叶用材、生物防火和生态防护树种,常见于华南及东南沿海各省区域,是亚热带常绿林阔叶的建群种,具有速生、丰产、材质优良和适应性强等特点[20],并被广泛用于营造混交林且效益良好[21-22]。因此,探讨木荷在变化光生境和异质养分环境中的化学计量特征和生长性状的响应规律和适应性,这不仅有助于揭示变化环境中木荷生存策略,也有利于木荷幼苗更新及混交林的科学经营管理,为亚热带低山丘陵区人工纯林树种结构优化与调控提供理论参考。
1 材料与方法
1.1 试验地概况
试验在浙江省杭州市中国林业科学研究院亚热带林业研究所虎山试验基地(119°57′E,30°05′N)进行。该地区所处海拔高度为30.5 m,属亚热带季风气候,温暖湿润,光照充足,年平均气温16.4℃,年日照时数1816 h。
1.2 试验材料
2019年2月中旬,于富阳周边灌丛地挖取试验所需生黄土,土壤在大棚内摊开经自然风干一个月后过筛,筛子孔径为1 cm × 1 cm。栽培基质中加入适量蛭石(直径3—6 mm),主要配比为生黄土∶蛭石=4∶1,栽培基质中全N、P含量为0.24 g/kg,0.11 g/kg,有效N、P、K和有机质含量分别为20.53 mg/kg,0.51 mg/kg,53.60 mg/kg,2.86 g/kg,土壤pH为4.87。栽培花盆规格为高35 cm,上直径25 cm,下直径15 cm。试验苗木为浙江省开化县林场提供的木荷一年生实生容器苗。
1.3 试验设计
本试验包括2种光照和4种施肥处理,共8个处理水平,每个处理12次重复。于2019年3月17日进行苗木栽植,栽植时仔细清洗容器苗的栽培基质后移栽至花盆中(栽培基质总重3.5 kg)。于4月中旬进行光照处理,分别设置全光照和遮阴处理(约45%全光照)。采用遮阴度为50%的黑色遮阴尼龙网遮盖,遮阴网顶部离地面高度约3 m,四周距地面约30 cm,用以通风。光照处理后,在晴朗天时随机选取3 d,采用TES-1335光照计(泰仕电子工业股份有限公司,台北)对大棚内光照组合遮阴组的光照强度进行测定,每次测定时间均为晴天中午,每次测定时重复5次,最后取平均值作为测定结果[23]。
于5月初对苗木进行施肥处理,在全光照、遮阴处理下均设置4个施肥处理:不施肥(CK),低N、P供应比例(N∶P=5),中N、P供应比例(N∶P=15),高N、P供应比例(N∶P=45)[9,17]。除不施肥处理外,其余每个处理N施入量均为100 mg/kg,通过改变P肥施用量(40 、13.4、4.5 mg/kg),从而形成4个施肥水平(NPCK、NP5、NP15、NP45)。试验氮肥为硝酸铵(NH4NO3),磷肥为磷酸二氢钾(K2HPO4,其中P2O5含量约为50%)。施肥处理时,将两种肥料混合成水溶液,共分5次施肥,20 mL/次,于一个月内施完成施肥试验,肥料水溶液现配现用,对照组施等量清水。除此之外,试验期间不定期挪动位置,防止边缘效应,所有苗木采用统一无差别管理。
1.4 样品采集和测定
2019年4月中旬,对缓苗后的木荷分别随机选取10样株,测定初始生长性状,苗木初始株高和地径分别为12.67 cm、0.14 mm,初始根、茎、叶和单株生物量分别为0.56 g、0.53 g、0.66 g、1.90 g。于当年10月中旬,每个处理随机选取6株苗木,对选定的苗木进行整株收获。将每个处理收获的样株分根系、枝干和叶片三部分,采集的样品编号后及时放入装有冰袋的保温箱内带回实验室。
将样品置于65℃烘干至恒重后称量根系、枝干和叶片的生物量。单株生物量=根干重+ 枝干重+叶干重;光合器官生物量=叶干重;地上生物量=叶干重+枝干重;非光合器官生物量=枝干重+根干重;地下生物量/地上生物量(根冠比)=根干重/(枝干重+叶干重);叶质比=叶干重/单株生物量;茎质比=枝干重/单株生物量;根质比=根干重/单株生物量。将烘干的根系、枝干、叶片经研磨后过0.1 mm筛,用于化学计量等相关指标的测定。C和N含量利用元素分析仪测定;P含量采用HClO4-H2SO4消煮—钼锑抗比色法测定;植物总C、总N和总P含量为叶片、枝干和根系C、N和P含量各自的加权平均值,以各部分生物量为加权系数。
1.5 数据分析
采用双因素方差分析(Two-Way ANOVA)检验光照和养分处理及其交互作用对生长性状的影响。采用单因素方差分析(One-Way ANOVA)和Duncan多重比较方法分析化学计量和生长性状在不同光照和养分处理下的差异。对不满足正态分布的指标在进行双因素或单因素方差分析前采用log对数转换,数据统计分析均在SPSS 22中进行,图表均在Excel 2007中绘制。
2 结果与分析
2.1 不同光照和N、P供应比例下木荷化学计量特征
由表1可知,绝大多数性状受光照、养分及其交互作用的显著影响(P<0.05)。除生物量分配外,其余性状受光照的影响大于养分及养分与光照间的交互作用。
据表2,NPCK组中,遮阴显著增加了根、茎和植物总N、P含量,但显著降低了根、茎和植物总C/N和C/P比(P<0.05)。遮阴显著增加了叶C、N、P含量,显著降低了叶C/N和C/P比,显著增加了叶N/P比。遮阴仅显著降低了根C含量(P<0.05),但对根N/P比、茎C含量、茎N/P比、植物总C含量和植物总N/P比无显著影响(P>0.05)。

表1 木荷化学计量和生长性状的双因素方差分析(F值)
在全光照组中,与NPCK相比,施肥显著增加了根C、N含量,显著降低了根C/N比,且随养分供应比例的变化而显著变化(P<0.05);NP45时显著降低了根C/P比。施肥显著增加了茎N含量,显著降低了茎C/N比,且随养分供应比例的变化而显著变化(P<0.05);NP5时显著增加了茎C含量(P<0.05);NP45时显著增加了茎P含量,但显著降低了茎C/P比(P<0.05)。施肥显著增加了叶N含量,显著降低了叶C/N比,且随养分供应比例的变化而显著变化(P<0.05);叶N/P随养分供应比例的增加呈上升趋势。施肥显著增加了植物总C含量(P<0.05);施肥显著增加了植物总N含量,显著降低了植物总C/N比,且随养分供应比例的变化而显著变化(P<0.05);NP45时显著增加了植物总P含量,但显著降低了植物总C/P比;在NP15和45时植物总N/P比显著增加(P<0.05)。
在遮阴组中,与NPCK相比,NP5时显著降低了根C含量、C/N比,显著增加了根N含量,NP45时显著降低了根C含量(P<0.05)。施肥显著降低了茎P含量,NP5和15时显著促进了茎C/P和N/P比,NP15和45时显著降低了茎C含量(P<0.05)。施肥显著增加了叶N含量(P<0.05);NP5时显著促进了植物总N含量(P<0.05);NP15时显著降低了植物总C、P含量,显著增加了植物总N/P比(P<0.05);NP45时显著降低了植物总C、P含量,显著增加了植物总N含量(P<0.05)。
2.2 不同光照和N、P供应比例下木荷各器官化学计量分配特征
据图1,当全光照转为遮阴时仅显著降低了根和茎C含量、叶P含量、根C/P比的分配。全光照处理中,N、P添加显著降低了根C含量(P<0.05),中低N、P供应比例时根系中P含量显著增加(P<0.05);而遮阴处理中,仅在高N、P供应比例为45时根C含量显著增加(P<0.05),N、P添加显著提高了叶片和根系中P含量(P<0.05)。无论是全光照和遮阴处理中,N、P添加一定程度上增加了茎C/P比,而N含量和C/N、N/P比在不同器官中变化趋势一致,N含量和N/P比为叶>根>茎,C/N比为根>茎>叶。
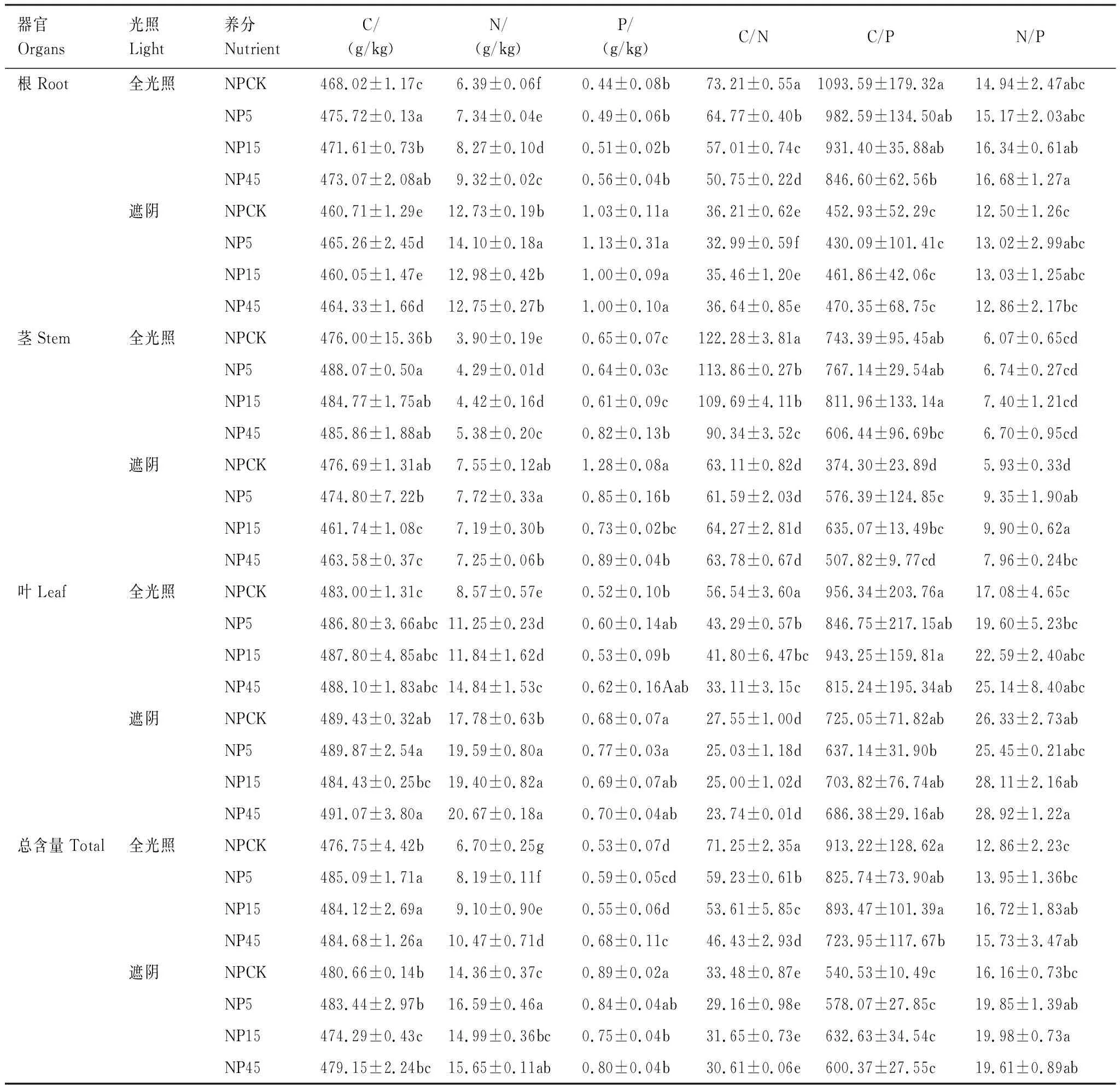
表2 不同光照和施肥下各器官化学计量性状的影响

图1 不同光照和施肥下各器官的化学计量分配特征Fig.1 Effects of different light and fertilization on stoichiometric allocation of each organs不同小写字母表示同一处理不同器官间差异显著(P < 0.05);NPCK:不施肥对照,Unfertilized control;NP5:氮磷供应比例为5 N-P supply ratio was 5;NP15:氮磷供应比例为15 N-P supply ratio was 15;NP45:氮磷供应比例为45 N-P supply ratio was 45
2.3 不同光照和氮磷供应比例下木荷生长及生物量分配特征
在NPCK组中,遮阴显著降低了木荷的单株生物量、叶干重、枝干重、地上生物量、根干重、根冠比和根质比(图2)(P<0.05),但显著增加了叶质比(图2)(P<0.05),对茎质比无显著影响(图2)。
在全光照组中,与NPCK相比,不同养分供应比例增加了单株生物量(图2)(P>0.05),但均显著降低了根干重、根冠比和根质比(图2)(P<0.05)。NP5时枝干重显著增加(图2)(P<0.05),NP15和NP45时茎质比显著增加(图2)(P<0.05)。NP5和NP15时叶干重和地上生物量显著增加(图2)(P<0.05),叶质比仅在NP15时显著增加(图2)(P<0.05)。
在遮阴组中,与NPCK相比,不同养分供应比例增加了单株生物量、叶干重、枝干重、地上生物量、根干重(图2)(P<0.05)。NP5时叶干重和叶质比在显著增加(图2)(P<0.05),茎质比显著降低(图2)(P<0.05)。NP45时茎质比显著增加(图2)(P<0.05)。

图2 不同光照和施肥对木荷生物量积累及其分配的影响Fig.2 Effects of different light and fertilization on biomass accumulation and allocation of S. superba不同小写字母表示同一器官不同处理间差异显著(P<0.05)
2.4 不同性状间的相关关系
据表3,各生长性状与各器官和总C含量、C/N及C/P比呈极显著的正相关关系(P<0.01),而与各器官和总N、P含量呈极显著的负相关关系(P<0.01)。根N/P比与各生长性状呈极显著正相关关系(P<0.01),而叶N/P比与各生长性状呈极显著负相关关系(P<0.05),茎N/P比与根干重、单株生物量呈负相关关系(P<0.05),总N/P比仅与根干重呈显著负相关(P<0.05)。茎质比、根冠比和根质比与单株生物量呈显著正相关关系(P<0.05)。叶质比与各生长性状呈极显著负相关关系(P<0.01)。
3 讨论
3.1 光照和N、P供应对木荷化学计量特征的影响
N、P是植物体内必需的大量元素,其含量和比例反映了植物的养分吸收、利用和分配等适应策略[24],作为功能性物质,N、P元素受环境影响较大,光照改变和养分添加均会对木本植物N、P分配格局产生影响[25],且在植物器官间分布根据器官结构功能的不同往往存在较大差异[26]。本研究中随光照降低,尽管木荷各器官和总N、P含量显著增加,但C/N、C/P比显著降低,这表明低光下木荷加强了养分吸收,但养分利用效率也随之降低。这是因为大多数植物在低光强下都倾向于加强N、P养分吸收,这有利于植物提高光合和资源竞争能力[27]。其次,遮阴下植物光合速率降低、生长减慢则导致N、P养分利用效率也随之下降,进而促使N、P在植物体内富集[17]。此外,植物根、茎、叶中的养分含量还取决于土壤养分供应和植被养分需求间的动态平衡[28],同一植物不同器官之间的C、N、P及其比值不仅受植物的基本生理过程需求的影响,还受相应器官组织结构和功能分化的影响[29]。无论全光照或遮阴处理下,木荷各器官和总N含量均随N、P供应比例的增加而显著增加,这是因为随着N、P供应比例增加,土壤中N素的相对有效可利用性增加,木本植物加速N获取促进蛋白质等有机物合成[30]。然而,相对C、N元素,木荷叶和根P含量对养分添加的响应较迟钝,在不同N、P供应比例间无显著差异,这可能主要与植物生活型及自身特性相关。研究表明草本植物根和叶P含量随N、P养分供应比例增加而显著降低,但根和叶N含量仅在低供应比例时变化显著[31],而木本植物幼苗N、P含量不仅受养分含量的影响,更与树种自身特性相关,不同树种的N、P响应规律甚至相反[32]。其次,还可能与植物限制元素相关,Han等[33]研究认为元素含量的变异范围还与该元素的限制情况有关,越是限制性元素,其含量变化越小,而本研究中木荷普遍受P限制,其较强的P素稳定性也提高了自身对低P环境的适应性。不同光照处理下木荷根和叶P含量无显著变化,仅低光照下不同N、P供应比例显著降低了茎P和总P含量增加,这是因为对木本植物而言,叶片和根作为植物生理代谢活跃的场所,在养分添加下需最大限度的维持P稳定性[34]。而植物茎主要起支撑和运输作用,作为木本植物重要的P养分库,在土壤有效P充足时“奢侈吸收”并储存P养分,而在土壤有效P不足时则向叶、根等其它器官输送P养分[34]。尽管木荷N含量对N、P供应比例的响应大于P含量,但其P分配格局的响应大于N分配格局。本研究中各器官间C、N含量基本遵循叶大于根或茎,这与其它木本植物C、N分配较为一致[35],保证叶C、N含量的分配有利于有机物的合成从而促进植物生长或防御机制生成[36]。然而,木荷各器官P元素的分配比例发生显著改变。首先,遮阴显著降低了叶P和根C/P比的分配,这与弱光下植物加大地上部分投入不同[37],木荷主要通过高P利用效率来慢速地上部分的生长发育,将更多的营养物质累积到根部,以促进根部发育,进而促进根系吸收能力[17]。全光照且N、P添加后,木荷根P分配显著增加,而遮阴且N、P添加后,木荷叶和根P含量的分配又显著增加,这主要与叶片和根系作为生长和养分吸收器官在养分增加下的代谢活跃相关[32]。

表3 化学计量与生长性状间的相关性
植物N/P比变化表征了植物维持体内化学计量内稳性的能力,通常被用来判断植物的N、P受限状况[38],反映了植物对环境变化的适应性。本研究中不同处理中各器官N/P表现均为叶>根>茎,相对根和茎而言,不同处理下叶N/P比相对稳定,这辅证了木荷幼苗叶N/P比来判断植物生长受限的合理性[16]。本研究中叶N/P随养分供应比例增加[9],仅全光照下不施肥和低供应比例时,木荷叶N/P小于20,其它处理的叶N/P均大于22,说明当前试验条件下木荷幼苗P限制随养分供应比例的增加而加剧,但这可能是过量的N吸收造成了植物体内N、P失衡及生长P受限性增强[39]。从N、P利用效率角度,植物在营养元素缺乏的情况下往往具有较高的养分利用效率,而在营养元素供应充足时,元素利用效率较低,这是植物对土壤养分状态的一种适应策略[31,40]。N、P添加后木荷各器官和总C/N、C/P基本与N、P响应规律相反,表明由N、P供应比例造成的N素可利用性增加反而降低了木荷的N利用效率[41]。与木荷N含量、C/N比不同,尽管高养分供应比例时木荷可能受P限制,木荷P含量与C/P比均无显著变化,表明木荷具有较高的P素稳定性且不受N、P供应比例的影响。
3.2 光照和N、P供应对木荷生长的影响
化学计量和生物量分配特征能直接或间接影响植物生长及其对异质环境的适应性及适应策略[42],在不同环境条件(光照和养分)下植物有不同的生长特性和物质分配规律[43]。多数研究认为光照是制约植物生长的首要因子,是植物进行光合作用所必需的资源,而对耐荫植物而言,适度遮荫有利于植物幼苗生长[44]。本研究中遮阴显著降低了木荷的株高、地径、各器官和单株生物量,说明45%全光照严重限制了木荷幼苗生长,尽管木荷具有一定耐荫性,其幼苗仍对光照有较高需求。低光强下,木荷的叶质比、茎质比、叶N和P含量显著增加,根冠比和根质比显著降低,这与其它木本植物对光照的响应类似,当光照不足时,植物往往分配更多资源到地上部分以捕获光资源,从而提高植物的光竞争能力和生存适合度[23,27]。
N、P作为植物生长必需大量元素,在个体发育和碳同化过程中具有重要作用,大多数研究认为N、P添加有利于促进植物生长[40,45]。本研究中木荷生长性状与各器官N、P基本呈极显著负相关,而与各器官C/N和C/P比呈极显著正相关,表明木荷生长主要与N、P利用效率相关。研究表明施N显著促进地上生物量积累、显著降低地下生物量,而施P显著促进根系分配,但对茎叶分配无显著影响[40]。类似地,本研究中木荷地上生物量、叶质比或茎质比在N、P添加处理中显著增加,根干重和根质比降低,表明木荷生长对添加的P含量不敏感,但对N的可利用性较为敏感。也有研究表明较低或较高的N、P供应比例往往造成营养元素之间相对关系的失衡,进而引发元素限制,将对植物的生长发育产生负面效应[27,46]。随N、P供应比例的增加,N素有效可利用性增加,但木荷单株生物量却与各器官N含量、叶质比呈极显著负相关关系,这表明过量N吸收导致了木荷叶生长过快从而导致自遮阴,影响了植物生长[46]。当遮阴和N、P添加共同作用时,木荷生物量及其分配的影响不显著,这是因为相对养分而言,光照是制约木荷幼苗生长的首要因子,生物量分配的无显著变化符合“李比希最小因子定律”。
本研究中木荷N/P比与生长性状基本呈极显著负相关,并暗示了木荷P限制随养分供应比例增加而增加,但单株生物量在各N、P供应比例间无显著变化,这表明木荷在不同N、P供应比例下的生长策略较为保守,这主要与木本植物自身的生长策略导致对环境中的养分供应比例的敏感性较弱有关[47]。相对全光照处理,遮阴处理下N、P添加对木荷生长的促进作用更为有限,说明低光照可能也部分掩盖了养分对木荷生长的全部效益[48]。尽管如此,与木荷生物量仅在高N高P或低N低P配施时显著增加存在相似之处[49],本研究中木荷幼苗生长亦偏好N、P供应量较高的生境。在N、P供应比例为5时,全光照处理下木荷地上生物量和叶C/N比显著高于其它处理,遮阴处理下叶干重、叶质比显著大于其它处理,这均有利于木荷的后期生长[50],表明木荷幼苗生长偏好较低的土壤N、P供应比例,且适量施肥在一定程度上有利于缓解低光胁迫。
4 结论
光照和N、P养分都是影响木荷生长发育的重要环境因子,其交互作用对木荷幼苗的影响也极为复杂,对木荷幼苗生长和化学计量特征产生显著影响。首先,光照不足导致木荷幼苗生长受到严重制约,木荷的生长策略较为保守,高N、P供应比例和遮阴环境均可能导致或加剧P限制。其次,尽管木荷化学计量和生长特征对N、P供应比例的响应较弱,但不同光照处理下,木荷幼苗生长均偏好低N、P供应比例,后续应加强更低N、P供应比例的研究。综上,木荷幼苗生长需要较高的光照和较低的土壤N/P条件,提高光照强度、合理调控土壤N、P供应比例将有利于木荷幼龄林生长,这对木荷幼林更新和人工林结构优化调控具有重要意义。
