干旱胁迫对黄山紫荆生长形态和生理特性的影响
2021-04-24田晓明陈新文颜立红蒋利媛向光锋李高飞谭知虎
田晓明,陈新文,颜立红,蒋利媛,向光锋,李高飞,谭知虎
(1.湖南省森林植物园,湖南 长沙 410116;2.江永县林业局,湖南 永州 431125)
植物所需养分来源于土壤。在不同的土壤类型上,植物的种类和生长状况也不尽相同。南方岩溶区是石漠化广为发生发展的生态脆弱地区,主要分布于云南、贵州、广西、湖南、广东、四川和湖北等9省[1]。石漠化地区由于土层瘠薄、水分流失严重、容易形成地表干旱,致使植株生长困难[2-3]。黄山紫荆Cercis chingii属豆科Leguminosae紫荆属Cercis落叶小乔木或丛生灌木,主要分布于安徽、浙江、广东北部、湖南南部等地[4-5],多发现在石漠化地区,是石漠化地区植被恢复的重要树种之一。目前,已有学者对紫荆C.chinensis[6-8]、高山杜鹃Rhododendron lapponicum[9]、杏Armeniaca vulgaris[10]、青冈Cyclobalanopsis glauca[11]和酸枣Ziziphus jujubavar.spinosa[12]等在干旱胁迫下植物的形态特征、抗氧化酶活性和渗透调节物质含量等方面进行了比较深入的研究。本研究采用盆栽控水法对持续干旱胁迫下黄山紫荆形态和生理生化指标的动态变化进行了研究,并对其抗旱能力进行评测,以期为石漠化地区植被恢复造林树种的选择提供科学的理论依据。
1 材料与方法
1.1 材料
供试材料选自湖南省森林植物园植物保育与利用研究所培育的黄山紫荆实生容器苗。于2015年3月,将1年生幼苗定植于规格一致的营养钵中(上口径为16 cm,下口径为12 cm,高为15 cm),每盆装混合基质(1黄土∶2珍珠岩∶1蛭石)0.8 kg,放置于湖南省森林植物园温室大棚中。试验开始前常规管理。
1.2 试验方法
自2015年8月26日开始,在温室大鹏内采用盆栽控水法进行干旱胁迫试验,室内外保持通风。设对照组(CK)和持续干旱组两个处理,每个处理90盆,CK处理每隔3 d正常浇灌至水分饱和;持续干旱组则在试验开始前充分浇灌,使土壤含水量基本保持一致,以后30 d内不再浇水,形成干旱胁迫。分别在第0天、第5天、第10天、第15天、第20天、第25天、第30天上午的同一时间,采集植株从下至上的第3至第7位叶片。每次采样时随机采5株植株的叶片,设3次重复,采样后的植株在下一个采样节点不再重复使用。将采集到的叶片装袋编号,迅速放置于冰桶中带回实验室进行生理生化指标测定。同时,在试验期间观察黄山紫荆叶片形态特征的变化。
1.3 生理生化指标测定
可溶性糖采用蒽酮比色法测定[13];可溶性蛋白采用考马斯亮蓝法测定[13];超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)还原法测定[13];过氧化物酶(POD)活性采用愈创木酚显色法测定[13];过氧化氢酶(CAT)活性采用紫外线分光光度计吸收法测定[13];丙二醛(MDA)含量采用硫代巴比妥酸法测定[13];游离脯氨酸含量采用用酸性茚三酮显色法测定[13];质膜相对透性采用相对电导率法[14]。
1.4 数据分析
采用Microsoft Excel 2010进行数据管理,Sigmaplot 10软件绘制图表;SPSS 19.0软件的独立样本T检验进行差异显著性分析。
2 结果与分析
2.1 干旱胁迫对黄山紫荆形态特征的影响
在持续干旱胁迫期间,黄山紫荆嫩梢和老叶最先做出反应。随着干旱胁迫时间的增加,叶片逐渐出现萎蔫卷曲、下垂、直至枯黄掉落,持续干旱30 d时,老叶颜色变淡、嫩叶枯落、嫩枝干枯(图1)。试验结束后,将持续干旱胁迫组植株恢复正常水肥管理,所有处理组植株经过一段时间恢复后均能正常生长。
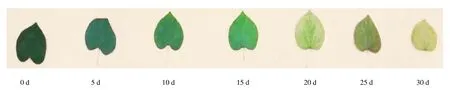
图1 黄山紫荆叶片在持续干旱胁迫下不同时段的颜色变化Figure 1 Color changes of C.chingii leaf at different stage of drought stress
2.2 干旱胁迫对黄山紫荆叶片内渗透调节物质含量的影响
如图2所示,随着干旱胁迫时间的增加,黄山紫荆叶片中可溶性糖的含量逐渐升高。在持续干旱胁迫≤15 d时,持续干旱组与CK间差异不显著,但干旱胁迫>15 d后,植株叶片中的可溶性糖含量持续升高且与CK间达均显著差异水平(P<0.05),说明可溶性糖对抵御干旱胁迫有重要的调节作用,可溶性糖含量的变化可以在一定程度上反映黄山紫荆对外界环境的响应情况。
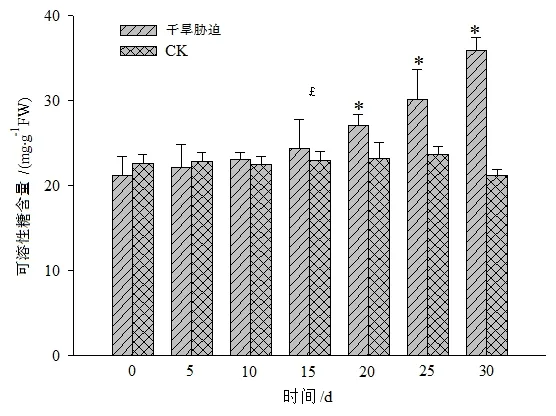
图2 干旱胁迫对黄山紫荆叶片可溶性糖含量的影响Figure 2 Effect of drought stress on soluble sugar content in leaves of C.chingii
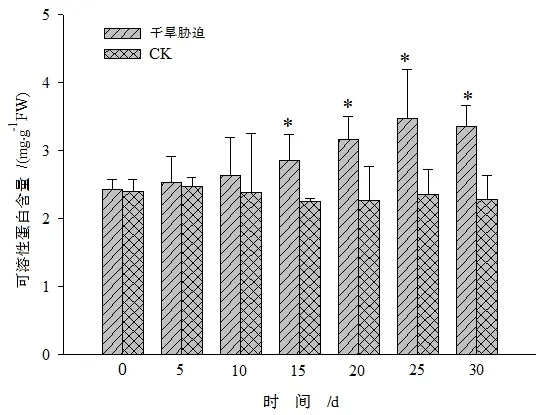
图3 干旱胁迫对黄山紫荆叶片可溶性蛋白含量的影响Figure 3 Effect of drought stress on soluble protein content in leaves of C.chingii
由图3可知,黄山紫荆叶片中可溶性蛋白含量在干旱胁迫期间整体呈现先升高后降低的趋势。在干旱胁迫初期(0~10 d),持续干旱组和CK间的差异不显著,从干旱胁迫第10天开始,可溶性蛋白含量急剧升高,至第25天时达到最大值,为CK的1.48倍。这可能是持续干旱促使一些与逆境胁迫相关的蛋白的合成和植物生理代谢的酶表达量的增加[15],而这些蛋白含量的增加是植株受干旱胁迫时的一种应激反应。
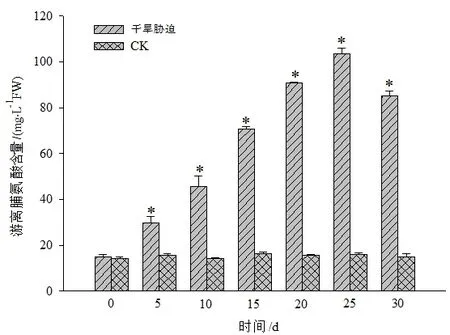
图4 干旱胁迫对黄山紫荆叶片游离脯氨酸含量的影响Figure 4 Effect of drought stress on free proline content in leaves of C.chingii
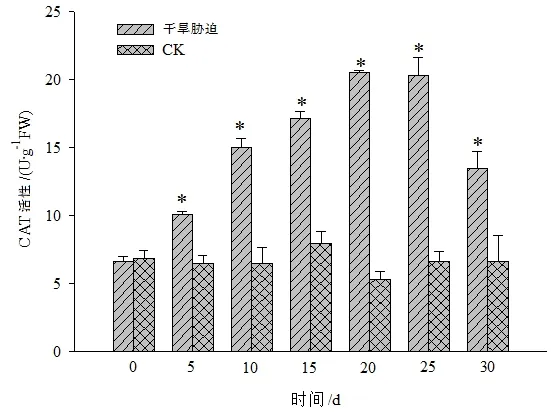
图5 干旱胁迫对黄山紫荆叶片CAT活性的影响Figure 5 Effect of drought stress on CAT in leaves of C.chingii
从图4可知,随着干旱胁迫时间的增加,黄山紫荆叶片中的游离脯氨酸含量有显著升高的过程,在干旱胁迫第25天时达到峰值(103.52 mg·g-1FW),随后呈下降趋势。黄山紫荆受干旱胁迫时,通过增加游离脯氨酸的含量来抵抗逆境环境,但这种抵抗能力有限,因此,当干旱胁迫强度超过植物所能承受的限度时,游离脯氨酸含量呈下降趋势。
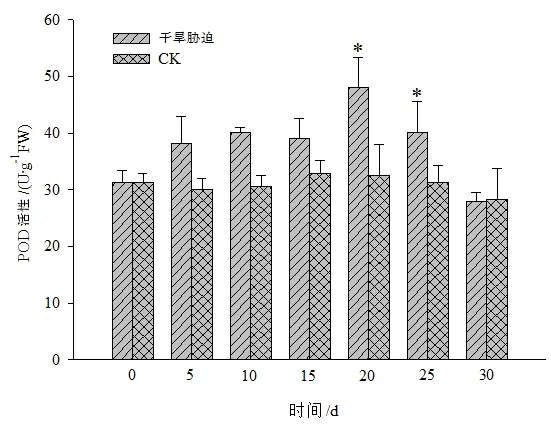
图6 干旱胁迫对黄山紫荆叶片POD活性的影响Figure 6 Effect of drought stress on POD in leaves of C.chingii
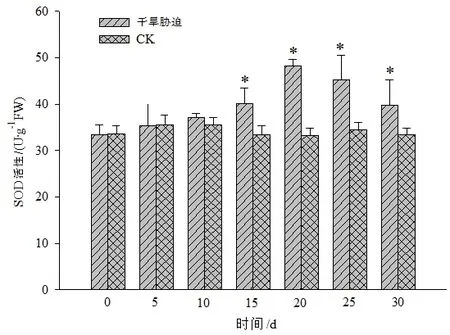
图7 干旱胁迫对黄山紫荆叶片SOD活性的影响Figure 7 Effect of drought stress on SOD in leaves of C.chingii
2.3 干旱胁迫对黄山紫荆叶片内抗氧化酶活性的影响
黄山紫荆CK处理叶片内的CAT、POD和SOD等抗氧化酶活性在不同时间的变化不显著,干旱胁迫组则随着干旱胁迫时间的增加整体呈先升后降的变化趋势,且在干旱胁迫第20天时3种酶的活性均达到峰值(图5至图7)。其中,CAT活性在干旱胁迫第20天时达到最大值后缓慢下降,在干旱胁迫25 d后,急剧下降,但仍显著高于CK。POD活性在干旱胁迫第20天时达到最大值后急剧下降,持续干旱30 d时,POD活性与CK之间差异不显著。SOD活性在干旱胁迫0~10 d时与CK之间差异不显著,在干旱胁迫15 d后急剧上升,干旱胁迫第20天时达到最大值后急剧下降。这说明黄山紫荆在干旱胁迫时通过增加抗氧化酶活性来去除逆境胁迫所产生的活性氧,因而在整个干旱胁迫期间黄山紫荆抗氧化酶活性均保持较高水平,以维持植株的正常生长;随着干旱胁迫时间的增加,逆境环境超出了黄山紫荆可自我调控的阈值,抗氧化酶活性下降,植株受到伤害。
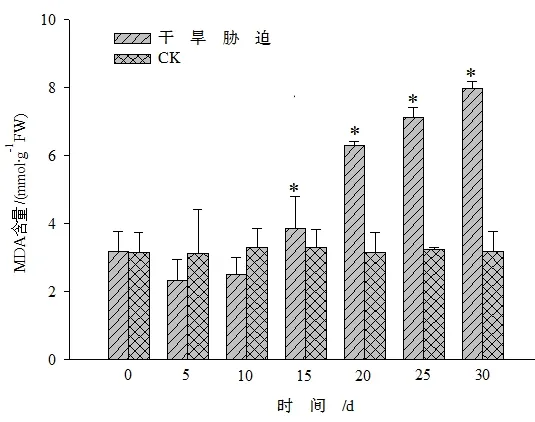
图8 干旱胁迫对黄山紫荆叶片MDA含量的影响Figure 8 Effect of drought stress on MDA content in leaves of C.chingii
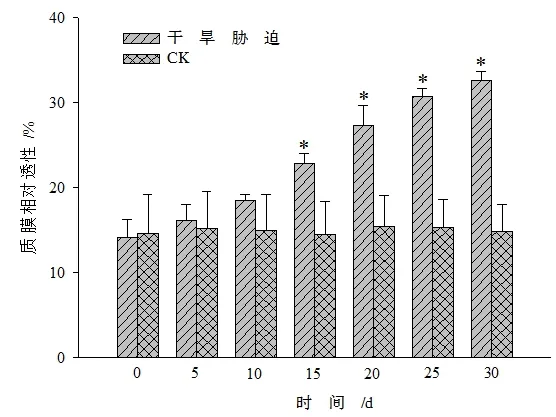
图9 干旱胁迫对黄山紫荆质膜相对透性的影响Figure 9 Effect of drought stress on relative membrane permeability in leaves of C.chingii
2.4 干旱胁迫对黄山紫荆叶片内MDA含量和质膜相对透性的影响
由图8表明,与CK相比,随着胁迫时间的增加黄山紫荆叶片内的MDA含量呈上升趋势。在干旱胁迫0~15 d时,MDA含量与CK之间的差异不显著,胁迫>10 d后,胁迫处理的MDA含量呈持续上升趋势,且与CK间达显著差异水平(P<0.05)。这可能是因为黄山紫荆对逆境胁迫有一定的适应过程,在胁迫初期当抗氧化酶系统产生防御作用时,MDA含量变化不显著。从图9中可以发现,受到干旱胁迫时,黄山紫荆叶片的质膜相对透性随着胁迫时间的延长而增大,到15 d以后(≥15 d),干旱胁迫处理与CK之间达显著差异水平(P<0.05),说明黄山紫荆叶片受到逆境伤害时脂膜过氧化程度逐渐升高。
3 结论与讨论
3.1 讨论
形态特征的改变是植物受逆境胁迫时产生应急响应最明显的表征[16]。通过试验观测发现,随着干旱胁迫时间的增加,黄山紫荆叶片、嫩枝逐渐出现萎蔫、枯落,干旱胁迫对黄山紫荆的生长产生一定程度的影响,但在极度干旱胁迫下(持续干旱胁迫30 d)黄山紫荆并没有出现死亡的现象,复水后也能正常生长。可见黄山紫荆可通过调整自身生物量来适应逆境胁迫。
可溶性糖、可溶性蛋白和游离脯氨酸是植物体内重要的渗透调节物质[17],当植物遭受逆境胁迫时,诱导植物体内多种生理反应加速,渗透调节物质逐渐积累,细胞保水能力得到加强,从而维持细胞结构的稳定和平衡[18]。在干旱胁迫伊始,黄山紫荆叶片中游离脯氨酸的含量明显增加,说明黄山紫荆通过增加游离脯氨酸来抵抗逆境,提高渗透调节能力,防止细胞内水分过多的丢失,王毅敏等[19]在巴东木莲Manglietia pat ungensis上的研究结果与此相同。可溶性糖和可溶性蛋白含量在胁迫处理初期与CK之间差异不显著,随着胁迫时间的延长,二者的含量逐渐升高,这与吴玲等[9]、黄燕等[20]和林伟通等[21]的研究结果相一致。胁迫处理后期,可溶性蛋白和游离脯氨酸含量降低,可溶性糖含量则继续升高,这与孔维鹏等[15]对无花果Ficus ca rica、赵银河等[22]对紫荆C.chinensis的研究结果一致。以上研究结果说明,随着胁迫程度的加剧,植物体内的蛋白质表达、糖和脯氨酸合成增加,以增强其渗透调节能力;而在胁迫处理后期,干旱胁迫超过了植物所能忍耐的阈值,植物体内的合成代谢不能正常进行,细胞质浓度也有所降低,细胞内有机渗透调节物质逐渐减少,可溶性蛋白和脯氨酸含量逐渐减少,植物受到伤害,这是植物体对极端干旱胁迫的响应机制。
植物处于逆境胁迫时,植物体会积累过量的活性氧自由基,打破了活性氧代谢的平衡,细胞膜磷脂结构遭到破坏[18]。此时,植物组织防御系统中的重要抗氧化酶如CAT、POD和SOD等的活性大幅升高,加强了对活性氧的清除,使活性氧的产生和清除达到动态平衡,减少膜脂的过氧化,维护细胞膜的稳定,减少细胞内有机渗透调节物质的渗出[23]。因此,膜脂的过氧化程度和抗氧化酶活性存在一定的相关性。逆境胁迫导致脂膜过氧化程度加剧、MDA积累,蛋白质和核酸等脂膜大分子结构发生变性,膜流动性降低,膜通透性增强。因而许多研究者将MDA含量和质膜相对透性来作为衡量细胞质过氧化程度和植物对逆境耐受能力的重要指标。通过研究发现,黄山紫荆的CAT、POD和SOD等活性随着干旱胁迫时间的增加呈先升后降的趋势,这与魏丽等[24]对紫荆、赖小莲等[25]对黄檀Dalbergia hupeana的研究结果相一致。在干旱胁迫0~15 d时,叶片中的MDA含量与CK之间差异不显著,15 d后,MDA含量才呈持续上升趋势。这可能是因为在干旱胁迫前期黄山紫荆的抗氧化酶防御系统先启动,而当干旱程度加剧,膜脂过氧化程度加深,防御系统逐渐失去作用,MDA含量开始上升,质膜相对透性逐渐增强,这与前人的抗旱性研究结果一致[23,26-27]。
3.2 结论
在持续干旱胁迫下,随着胁迫时间的增加黄山紫荆植株萎蔫、嫩枝和老叶枯落,是叶片渗透调节物质、抗氧化酶活性和MDA等物质综合响应干旱胁迫的结果。在干旱胁迫早期,黄山紫荆能够通过自身调节维护体内的生理平衡,通过增加游离脯氨酸含量和抗氧化酶活性来适应逆境环境;在干旱胁迫中期,则需通过维持较高的渗透调节物质、抗氧化酶活性和MDA的含量来抵御干旱胁迫,减轻干旱伤害;在干旱胁迫后期,黄山紫荆体内代谢紊乱,各项指标值降低,植株受到伤害。黄山紫荆具有较好应对干旱胁迫的调节能力,这也是黄山紫荆在石漠化山地广布的原因。
