黄牛木种群空间分布格局研究
2021-04-24韦彩丽谢正生邓丽婷何晓慧孔令华
韦彩丽,谢正生,邓丽婷,何晓慧,孔令华
(华南农业大学 林学与风景园林学院,广东 广州 510642)
黄牛木Cratoxylum cochinchinense是亚热带地区的先锋树种[1],亦是我国南方地区的乡土树种[2],喜光,耐贫瘠,抗逆性好,常野生于向阳杂木林中,生长慢但萌芽力强,在石山的石崖灌丛中生长良好[3],其黄褐色树干形成的特色林内景观以及春夏形成的彩色繁花景观具有较高的观赏价值,是极具潜力的城市景观树种[4]。2018年10月,深圳市和中山市顺利获批为“国家森林城市”,至此,珠三角地区的9个城市全部成为“国家森林城市”,标志着我国首个森林城市群——珠三角森林城市群顺利创建。但是,该地区仍存在较多困难山林地亟需进行生态修复。黄牛木因其优良的抗逆特性和景观价值,可作为“新一轮广东绿化大行动”中景观优化、林相改造等生态修复工程的理想备选树种。
空间分布格局是种群生态学研究的热点内容之一。关于种群空间分布特征的研究,国内外学者已有大量的报道[5-7],研究方法也较为成熟[8-9]。但是,关于黄牛木选种、培育及种群生态学方面的研究及报道几乎为空白。为揭示黄牛木种群的生长及分布规律,本研究对黄牛木天然林集中分布区进行了实地调查,分析其种群空间分布格局和动态变化规律,以期促进黄牛木在珠三角地区森林生态修复中的推广和应用,为其天然林的抚育管理提供理论基础和科学参考。
1 材料与方法
1.1 研究地概况
分别在广东省的广州市和台山市布设样地。广州市是珠三角核心城市,地理位置为112°57′~114°3′E,22°26′~23°56′N,属海洋性亚热带季风气候,多台风,全年平均气温在20~22℃,平均相对湿度为77%,年降水量约为1 720 mm,雨量充沛,利于植物生长,为四季常绿、花团锦簇的“花城”。台山市位于珠江三角洲西南部,地理位置为112°81′~112°9′E,22°26′~22°81′N,属亚热带海洋性季风气候,全年平均气温为21.8℃,年平均日照为2 006 h,年均降水量为1 936 mm。
1.2 研究方法
1.2.1 样方设置与调查 通过查阅广东植物种质资源相关文献[10-11]以及珠三角地区各个城市的地方植物志,2016-2017年,陆续对珠三角地区进行林分踏查,实地考察各城市的植物组成及黄牛木种群的分布情况。研究采用典型样地法,在黄牛木集中分布区选择具有代表性的天然群落设置样地,分别在广州市茶山公园的鸡笼顶(东南坡)设置样地3 900 m2、白云山风景名胜区的林园景区(西北坡)设置样地3 900 m2以及台山市石花山森林公园的西严寺东侧设置样地3 200 m2(其中东南坡2 000 m2,西北坡1 200 m2),样地总面积为11 000 m2。在外业调查过程中,测定样地内树高>1.3 m的黄牛木个体的胸径、冠幅、树高、枝下高等生长因子,树高≤1.3 m的黄牛木个体测定地径、冠幅、树高等,并记录所有黄牛木个体的相对坐标。同时,将样地划分成10 m×10 m的栅格样方,在每一个10 m×10 m栅格的四个角以及对角线位置上布设2 m×2 m的小样方,记录小样方内所有的植被种类及群落生长状况。
1.2.2 群落结构划分 群落结构从树冠到地面进行划分,研究将黄牛木样地中小样方内的植被种类分为乔木层、灌木层、草本层以及层间植物。其中,乔木层可分为两个亚层,第一亚层高度在8 m以上,第二亚层高度在4 m以上,覆盖度在50%以上;灌木层高度一般在3 m以下,覆盖度明显受到上层林木的影响,一般为30%~40%,以常绿阔叶矮高位芽植物为主;草本层一般高度在l m以下,覆盖度在10%以下,局部分布或聚集生长;层间植物主要指藤本种类,包含寄生、附生植物。
1.2.2 种群发育阶段划分 采用“空间替代时间法”,即以林木的胸径大小作为指标来度量其年龄大小。参照林木发育阶段的划分方法[12],将不同胸径(DBH)大小的黄牛木个体划分为5个不同的龄组:幼苗(DBH<1 cm,树高≤1.3 m)、小树(1≤DBH<5 cm)、中树(5≤DBH<11 cm)、大树(11≤DBH<15 cm)、老树(DBH≥15 cm)。
1.2.3 种群空间分布格局分析 将实地调查所得数据进行数字化处理,导入ArcGIS信息平台,分别绘制各样地黄牛木种群的空间分布点位置图,结合方差均值比率法和聚集度指标探讨种群的分布格局。
(1)方差均值比率法 方差均值比率法建立在Poisson分布的预期假设上,一个Poisson分布的总体有方差(S2)和均值(X)相等的特性,即S2/X=1。如果S2/X>1,则种群趋于集群分布,如果S2/X<1,则种群趋于均匀分布。方差和均值的计算方法如下:

式中,N为基本样方的个数,Xi为第i个样方内的个体数。
实测与预测的偏离程度可用t检验验证,t值的计算公式如下:
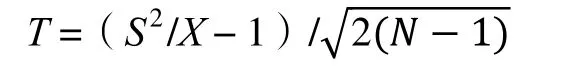
通过查自由度(N-1)、置信度为95%的t分布表,进行显著性检验。当|t|<tN-1,0.05双侧时,种群为随机分布,否则为聚集或均匀分布。
(2)聚集强度指标
①负二项参数(K)的计算:

当K< 0时,种群呈均匀分布;当K> 0时,种群呈聚集分布;当K值趋于无穷大时(一般为8以上),种群逼近Poisson分布,呈随机分布。K值愈小,种群聚集程度愈高。
②扩散指数(C)的计算:

当C=1时,种群呈随机分布;当C>1时,种群呈聚集分布;当C< 1时,种群呈均匀分布。
③Cassie指数(Ca)的计算:

当Ca=0时,种群呈随机分布;当Ca> 0时,种群呈聚集分布;当Ca< 0时,种群呈均匀分布。
④丛生指数(I)的计算:
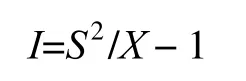
当I=0时,种群呈随机分布;当I> 0时,种群呈聚集分布;当I< 0时,种群呈均匀分布。
⑤平均拥挤度(m)与聚块性指标(m/X):当m/X=1时,种群呈随机分布;当m/X> 1时,种群呈聚集分布;当m/X< 1时,种群呈均匀分布。
2 结果与分析
2.1 黄牛木群落结构
外业调查发现,3个样地均为黄牛木天然群落,植被繁茂,多为华南地区常见种类。群落结构相似且成层现象明显,调查样地中植物高度多集中于2~10 m。对小样方内的群落组成和分层情况进行统计,结果见表1。
从表1可以看出,调查样地的植物种类共有122种,隶属49科96属。其中,茶山样地共有38科62属74种,不低于5种的科分别为樟科Lauraceae、大戟科Euphorbiaceae、含羞草科Rubiaceae、桑科Moraceae;白云山样地有31科52属59种,其中,茜草科Rubiaceae和禾本科Gramineae的属数和种数最多,均有6属6种,分别占白云山样地总属数和总种数的11.54%和10.17%;石花山样地有41科70属78种,其中,大戟科的属和种明显多于其他科植物,有7属9种,分别占石花山样地总属数和总种数的10%和11.54%;茜草科植物种类次之,有6属8种。从科、属、种的数量来看,均表现为石花山样地>茶山样地>白云山样地,表明黄牛木样地植被丰富度依次为石花山样地>茶山样地>白云山样地。
从表1还可以看出,调查样地各层级植物种类数量由多到少依次为灌木层(41种)>乔木层(40种)>草本层(29种)>层间植物(12种),分别占植物种类总数的33.61%、32.79%、23.77%、9.83%。其中,茶山样地的乔木层有28种,灌木层有23种,草本层有16种、层间植物有7种,分别占样地种类总数的36.84%、30.26%、22.37%、10.53%;白云山样地的乔木层有13种,灌木层有22种,草本层有16种、层间植物有8种,分别占样地种类总数的21.41%、37.93%、27.59%、12.07%;石花山样地的乔木层有18种,灌木层有30种,草本层有20种、层间植物有10种,分别占样地种类总数的23.08%、33.33%、23.58%、9.76%。
2.2 种群空间分布格局分析
通过对调查的数据进行处理和分析,借助GIS地理信息平台,将黄牛木个体的分布格局进行直观化呈现,得到黄牛木种群空间分布点位置图,见图1。
从图1中可以看出,不同研究样地种群个体的聚集程度存在一定的差异,单个样地的黄牛木种群呈现从某个中心位置逐渐向外扩散的趋势。运用方差均值比率法和聚集度指标测定3个样地的黄牛木种群分布格局,结果见表2。从表2可知,不同样地下,黄牛木种群的t检验结果表现为聚集或均匀分布,负二项式参数K值表现为聚集分布。研究表明,当样方内个体聚集程度极低或极高时,t检验与实际有所偏离,而K值检验几乎不受影响,故主要以K值来判定种群的空间分布格局,K值越大表示种群的聚集程度越低,反之则高。因此认为,研究样地的黄牛木种群均为聚集分布,聚集程度为白云山样地>茶山样地>石花山样地。同时,对种群的聚集度指标Cassie指数、丛生指数以及聚块性指标进行分析,所得结果也呈现与之相一致的结论,这与图1呈现的种群空间分布情况相吻合。
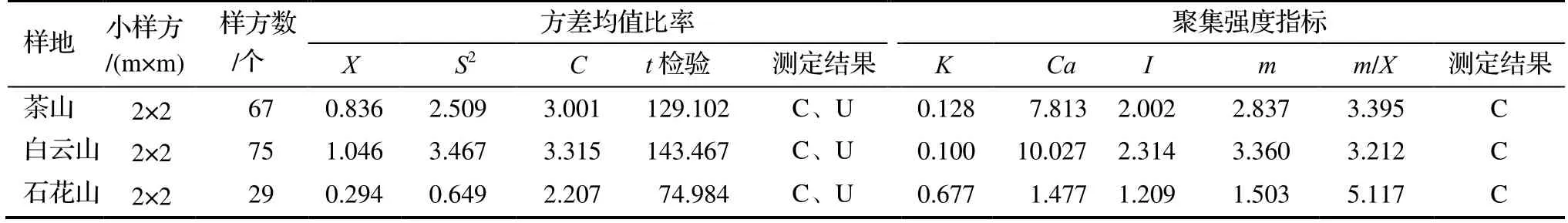
表2 不同样地的种群空间分布格局测定结果Table 2 Spatial distribution pattern of C. cochinchinense in different sample plots
2.3 坡向对种群空间分布格局的影响
种群的海拔、坡向等生境条件不同,其分布格局可能存在一定的差异。由于各样地之间的海拔差异较小(茶山样地海拔85~93 m,白云山样地海拔130~139 m,石花山样地海拔90~96 m),故研究主要探讨坡向对种群空间分布的影响。广州和与台山两地相距约130 km,相比之下,广州茶山和白云山相距约6 km,可近似认为位于同一地理位置。采取胸径代替年龄时间进行分析,运用方差均值比率法和聚集度指标测定相同样地不同坡向的种群空间分布格局,结果见表3。

表3 不同坡向的种群空间分布格局测定结果Table 3 Effect of aspect on spatial distribution pattern of C. cochinchinense in different sample plots
从表3中可以看出,茶山-白云山样地东南坡的K值为0.128,西北坡的K值为0.100,表明其聚集程度为西北坡>东南坡,即阴坡>阳坡;石花山东南坡的K值为-0.516<0,为均匀分布,西北坡的K值为0.222>0,为聚集分布,表明其种群聚集程度亦为西北坡>东南坡,即阴坡>阳坡。综上,黄牛木种群的聚集程度表现为阴坡>阳坡,说明种群对阴坡具有一定的选择性。
2.4 不同发育阶段种群空间分布格局分析
黄牛木种群特征始终处于动态变化中,不同生长时期的黄牛木种群斑块大小不同,对环境空间大小和生存条件的要求也有不同。同一个体的密度、不同物种间的竞争、动物的影响以及阳光、水分等环境因子的不均匀分布,都有可能改变种群的聚集分布和空间格局。运用2.3中的方法测定不同年龄阶段的种群分布格局动态,结果见表4。
从表4中可以看出,在不同的发育阶段,各个样地的黄牛木种群空间分布格局存在差异,但总体分布趋势一致。茶山样地和白云山样地中的黄牛木在幼树和小树阶段表现为聚集分布,在中树和大树阶段为均匀分布,老树则为随机分布;石花山样地的黄牛木仅在幼树阶段表现为聚集分布,小树到大树阶段为均匀分布,老树为随机分布。综上,随着年龄的增大,黄牛木种群呈现出扩散趋势,种群从幼树至老树的时间序列上,空间分布格局表现为从聚集分布至均匀分布至随机分布。

表4 不同发育阶段的种群空间分布格局测定结果Table 4 Spatial distribution pattern of different aged C. cochinchinense in different sample plots
3 结论与讨论
3.1 结论
茶山、白云山、茶山3个样地均为黄牛木天然群落,植被繁茂且多为华南地区常见种类,各样地植被丰富度表现为石花山样地>茶山样地>白云山样地。黄牛木群落结构相似且成层现象明显,各层级植物种类数量由多到少依次为灌木层(41种)>乔木层(40种)>草本层(29种)>层间植物(12种)。各样地黄牛木种群的空间分布格局均表现为聚集分布,在母树周围的聚集分布现象最为明显,并呈现逐渐向外扩散的趋势。不同样地的种群聚集程度存在差异,表现为白云山样地>茶山样地>石花山样地。不同坡向黄牛木种群的聚集程度存在差异,表现为西北坡>东南坡,即阴坡>阳坡,表明黄牛木种群对阴坡具有一定的选择性。不同发育阶段黄牛木聚集程度呈现从幼树、小树、中树至老树逐步降低的现象,空间分布格局表现为从聚集分布至均匀分布至随机分布;随着发育程度的增加,种群具有扩散趋势。
3.2 讨论
种群是群落结构和功能的基本单位,不同种在同一空间的分布存在差异,同一个种在不同发育阶段、不同生境条件下也有明显差异[13-15]。种群空间分布格局是指个体在水平空间的配置和分布状况,是个体与环境长期适应和选择的结果[16],反映了环境变化、各种干扰、竞争作用、生活史策略以及森林更新等各种因子的综合作用[17]。种群在一定空间内都存在一个特有的分布格局,它可以反映物种之间的关系、环境适应机制和空间异质性等[18]。根据种群中个体的聚集程度,种群空间分布格局的类型一般可分为随机分布、均匀分布和聚集分布。本研究中的黄牛木种群呈现聚集分布,与其自身的生物学特性及其所处的生境特征密切相关,尽管黄牛木种子具翅,但其传播能力有限,使得幼苗多聚集在母树周围。同时,黄牛木种子数量大,自然脱落后易聚集萌发,因此,其种群多呈现显著的集群分布,这与前人的研究结果具有相似之处[19-20]。不同样地的种群聚集程度存在差异,可能是种子传播扩散条件、生境异质性、种内-种间竞争等因素相互作用的结果,也可能是取样尺度的影响。一般来说,种群在小尺度范围内表现为聚集分布,而在大尺度上则表现为随机分布,本研究仅对实际取样尺度进行分析,难免因种群对尺度的依赖性而导致分布格局的研究结果受到影响。
胥晓等[21]通过对不同坡向的四川冶勒红豆杉Taxus w allichianavar.chinensis种群空间分布格局的研究证实了种群分布对坡向具有选择性这一结论。黄牛木种群对阴坡具有一定的选择性,其原因可能是阴坡水分条件较好,有利于黄牛木种子的萌发;同时,位于同一海拔的光照强度阴坡较阳坡温和,更适合黄牛木幼苗的存活与生长。不同坡向影响着种群的分布格局,意味着在森林经营上可充分利用自然力,采取相应的抚育措施;不同坡向黄牛木种群的聚集程度大小,可作为造林密度对坡向的选择依据。
对种群不同发育阶段空间分布格局的研究报道较多[22-24],黄牛木种群不同发育阶段的研究结果支持前人的结论。这种从幼树、小树、中树至老树聚集程度逐步降低的现象,是由于随着幼树、小树的生长,植物对光照、水分以及营养条件的需求加大,为争夺足够的生存空间与资源,个体之间的竞争就会加剧,竞争力低的个体死亡,种群密度就可能下降,存活下来的个体呈扩散趋势,种群格局就可能发生变化;从小树发育成为中树的过程中,种群对空间、阳光、水分和其他资源的需求进一步加大,种内竞争更为激烈,从而造成自疏和它疏现象,使得种群的聚集强度进一步下降而呈现均匀分布;如果扩散慢,则种群仍保持为均匀分布,如果扩散快,种群分布类型就会发生变化,由均匀分布趋向随机分布。种群的这种分布格局变化模式反映了新生个体的产生、成年个体的死亡和人为干扰等[25]。梅世秀等[26]对不同发育阶段锐齿槲栎Quercus alienavar.acuteserrata种群的分布格局进行了研究,指出不同地区、不同年龄的锐齿槲栎林应采取不同的经营策略和管理措施,这对黄牛木种群的抚育管理具有借鉴意义。
在黄牛木天然林的保护管理和珠三角地区的绿化建设中,应采取以下措施:一是深入了掌握其分布及生长规律,了解周边环境对其生长演替的影响,适时进行人工疏伐,促进其天然群落的生长和更新;二是进一步研究黄牛木天然群落的生境条件,总结黄牛木天然林的植被组合特征,为林相改造工程的树种选择和配置提供参考;三是开展黄牛木种苗的繁殖培育工作,为造林工程提供充足的优质苗源。本研究对黄牛木种群的空间分布特征、生境选择以及各个发育阶段的生长分布规律进行了分析,为黄牛木种群的深入研究提供理论基础,下一步要扩大取样范围,并选择空间点格局分析等不受尺度影响的方法进行深入研究,提高研究结果的准确度和说服力,同时要对单株个体扩散速率进行研究,确定其单位面积的保存量和间伐量,促进其天然林的更新生长和保护管理,加快其在生态建设中的推广和运用。
