临近根系间的亲缘识别、行为应对及其功能性状可塑性进化
2021-04-24肖万欣王延波张书萍
肖万欣王延波张书萍
(辽宁省农业科学院,辽宁 沈阳 110161)
植株受相邻物种根系竞争的影响,如何调节生长和生物量是一个具有重要意义的课题,其中重要的一点是要知道植物根系是如何察觉到邻居的存在并做出反应的[1]。有些植物根系能够通过“自我—非自我”识别模式感触并通过空间错位回避邻近根系;有些植物能够试图通过根系增殖在非同缘竞争对手存在时抢占资源和空间,避免与有亲缘关系的根系竞争[8],表现出合作行为,从亲缘识别中获益[3],另一些物种似乎缺乏这种能力,这种能力差异的影响因素尚未查明[3]。根系分泌物通过介导植物相互作用、促进有益微生物相互作用和影响养分循环,潜在影响着植物群落的生产力和组成。然而,对于根系分泌物在植物群落动态中的作用,特别是在田间条件下,还缺乏充分证据[11]。同时,不同形态性状对生态位分化的贡献以及抑制和容忍竞争对手能力的证据还非常有限[10]。在形态可塑性进化方面,也只从植物空间聚集和环境变化等变量开展一些研究[20]。当前的植物亲缘识别和功能性状可塑性进化等研究还缺乏深入和系统性,急需加强相关方面的研究工作。掌握植物根系对邻居识别和应对机制对生态系统功能和植物进化研究以及对自然保护和减少农作物间竞争均具有重要意义[4,5]。
1 影响临近根系间亲缘识别的因素
1.1 内在因素
1.1.1 根系性状 植株有能力限制个体行为所产生的能量损耗,通过改变形态而不是改变生物量的分配来增加根系密度[8]。在地下物种互作竞争中,成功的物种可能较少地依赖于竞争个体间的形态大小,而更多地依赖于竞争忍耐度的相关特征[9]。虽然,更大的根系和更广泛的水平扩展以及更低的根系组织密度能够有效地抑制邻近植物,然而,在竞争变异的最佳多性状模型中,不包括根系大小,而只包括根系形态、分支和垂直分布等与根系竞争耐受性有关的指标,而与抑制邻居的能力无关[10]。
1.1.2 根系分泌物 邻居的根系分泌物可能会创造一个引起局部和系统协同反应的土壤异质环境,这就会导致根系生长和形态的同时变化。无毒的根系分泌物在协同进化的生态系统中扮演着重要的角色,可以调节植物间的交流[11]。在识别邻居物种功能方面,已有研究表明,无毒的根系分泌物需要像暴露在入侵物种分泌的有毒分泌物中的大多数植物一样,也需要本地适应。植物能够探测到周围个体的邻近性,并对具有不同遗传特性的植物的临近做出不同的反应[18]。已有研究证实,在遗传距离较远的个体间,邻居识别是由根系分泌物介导的,并以牺牲繁殖为代价,促进根系增殖,从而获得较高的竞争能力,这种现象被描述为“公地悲剧”[12]。相关研究表明,根系分泌物可介导亲缘识别,并触发根的“行为”变化[2,8]。例如,根系分泌物可以携带邻居基因血缘、种群起源和物种身份特定信息,并在整个根系水平引发不同的反应,并在个体根系水平与本地分泌物完成直接对话。当目标植物暴露在与其基因无关的植物根分泌物中时,它们的根长和密度会增加。
1.1.3 化感物质与化学信号 由于土壤中缺乏光照,土壤生物的行为常常以感知化学信号为基础[13]。在无性系中,与不同类型的临近根系一起生长的植物,在不同物种之间表现出的根系行为模式是不同的,除获取资源外,其他因素可能参与了对它们的根系生长行为的调控,即邻近的根还能通过释放化感化学物质和无毒信号来广泛地影响植株的生长。根系分泌物含有多种植物的初级和次级代谢产物,其中一些已明确显示具有化感作用[14]。化感作用释放的有毒的生物化学物质(化感化学物质)抑制邻近植物生长,通过改变土壤微生物群落,直接或间接地影响其他物种的营养动态和资源竞争[4]。它们对邻近植物生长的影响取决于根系分泌物中的化感物质浓度和释放的量、临近植物生育时期和生物与非生物环境条件。如果它们被释放量较大且在环境中维持活性时间较长,那结果肯定是负面的。在受控条件下,目标植物的生长期、生物量和密度是改变向根际释放的化感物质有效性的重要因素,在进行化感物质研究时应加以考虑。例如,与密度较小的群体相比,密度较大的群体就需要提供浓度更高的化感物质,才能产生相似的结果。可见,在一个地点得出的结论,在另一个地点不一定能被重复,因为,它们所处的种群密度有可能不同。
1.2 外在因素
1.2.1 基质体积 植物所占的基质体积对其产量影响较大。例如,同样营养量供给的植株在较大基质体积下营养生长量更大。植物根竞争反应的实验,通常是在恒定基质体积和每株营养等量下进行的。不同物种在生长敏感性和向生殖器官生长量分配上,对基质体积的反应不同。“公地悲剧”和“自我—非自我”识别的实验中得到的某些结果确实归因于基质体积的不同。两株根系存在竞争共存时,其每株根系体积往往大于它们单独生长时的根系体积。研究表明,分隔盆栽中,空间越有限,根系的化学自抑制能力越强;在未被分隔、存在两株根系竞争的盆栽中,基质体积越大,无效根分布量也越大。当根系生长空间缩小时,根系对土壤资源获取量减少,导致根系结构、形态和激素的变化。
1.2.2 个体间形态和生物量的均匀度 单个个体间生物量差异小、整体均匀度高是产生“公地悲剧”的原因之一。如果群体内个体间形态和生物量不均匀,那么,观察到的“公地悲剧”将是一种由不对称竞争引起的数学必然性结果,而不是生物过程的结果。当基质体积和植物生物量之间存在负向关系时,即与竞争者一起生长的植物生物量与植物单独生长的生物量表现较大的不一致(由大小不对称引起的竞争),那么,与竞争者一起生长的植株(劣势株),其单株平均生物量将低于每株单独种植时的单株生物量。同时,由于根冠异速生长,平均植株生物量的减少也会引起较低的根冠比。因此,临近根系间亲缘识别相关研究,应尽量保证每个个体生物量与群体内总生物量平均值一致。
1.2.3 土壤异质性 即使每个植株的资源量和基质体积保持不变,竞争对手的根系为获取资源也会产生资源耗竭区,从而形成一种比单独种植植株时更不均匀的土壤资源分配模式,即土壤异质性。如果竞争者的根系重叠,单位根长的养分吸收量可能会更低,因为,仅存的较少的资源有可能已被竞争对手获取。尽管土壤异质性会显著影响植物生长,但在以往关于亲缘识别和公地悲剧的研究中,临近根系间的养分吸收与分配相关研究较少。
1.2.4 相邻物种密度 资源竞争的强度受植株密度的影响,密度可能会强烈影响植物对邻居的遗传身份的反应。当面对非亲缘邻居时,植物会随着邻居密度的增加而增加自身竞争特性的投入(如根系增殖);而当被亲缘邻居包围时,这种反应被抑制。在邻居密度较低条件下,亲缘识别也可能触发不同的表型,这意味着只有在不同邻域密度下观察植物,亲缘识别才可能真正被检测到,并完全理解它。研究表明,遗传亲缘关系对植物性状的影响一直依赖于相邻密度。随着邻近植物密度的增加,有亲缘关系的植株用于种子繁殖的投入显著提高,在竞争器官上的生长分配减少,而在没有亲缘关系的个体之间没有表现出这样的反应[3]。
2 亲缘识别中的根系行为应对策略
2.1 根系行为模式
第一种是回避模式:两个相互竞争的植株根的分离导致根在基质中分布相对均匀(图1)。因此,植物间基质中一定体积区域内的根质量应远远小于距离相邻方向相同大小的两体积基质中根质量的总和(图1中F3区域的根量 < F2+F4区域的根量。这一预测假定两株植物之间的距离足够远,以至于每一株植物的根都不会延伸到F3的远端边界之外)。第二种是侵入模式:如果根系向邻近植物的根系生长,就会出现高度混杂的根系。两株植物间同一基质体积的根量应大于相邻两体积基质中根量之和,且两体积基质的根量大小相等(图1中F3区域的根量>F2+F4区域的根量)。第三种是无响应模式:如果根对邻居的根的存在没有任何反应(这包括对其他根系创建的资源耗竭区没有反应),那么,这样的情况,根系有可能是混合的,但混合量小于侵入模式。两株植物之间一定体积基质中的根质量应等于两体积基质中根质量的总和,两体积基质的大小相等,且朝向相邻的方向(图1中F3区域的根量=F2+F4区域的根量)。
如果相互连接的根系之间存在整合,所观察到的根系生长模式将是根对其当地环境的直接反应结果,并且受到同一植株上不同条件的其他根系影响。根据竞争对手的根的特性,植物也可能表现出不同的根系布局策略(表1)。此外,从根的完全分离到根的极端聚集,可能会观察到根系行为是一个连续的过程,而不是上述3种离散模式。
2.2 根系应对策略
根系间亲缘识别常常发生在地下资源竞争最为激烈的根系重叠区,即资源耗竭区。不同物种的根系在所形成的耗竭区内广泛的重叠,导致了根系对土壤资源的竞争,降低了资源获取的效率,因为相同的土壤体积内的资源将由多个根同时进行“觅食”,可见,如果同一植物的根具有与基因型相同的个体间根系分离的能力(避免耗竭区重叠),那将对它平衡资源分配特别有利。当有基因型不相关的竞争者存在时,有些植株也有能力将根系增殖至竞争者根系占据的空间里来潜在地提高相对适合度,减少根系间重叠,吸收新的资源,同时阻止竞争对手获得这些资源。因此,植物可能受益于具有区分“自我”和“非自我”根的能力,当遇到自己的根或与自己的根密切相关的个体,通过重新定向或抑制根系生长,并通过占用与自己相关性很小或不相干的竞争者的空间来增加根系生长。或者,即使没有一个特定的机制来识别竞争对手,植物也可以依靠对相邻根系属于同一基因型或不同基因型的长期概率采取不同的策略(例如,相邻根的隔离和混种),这种策略在不同物种间变化较大,例如,在排列密集的无性系(密集方阵式)与分布广泛的无性系(游击式)物种中和在物种多样性的不同群落中,植物采取的策略均不相同。
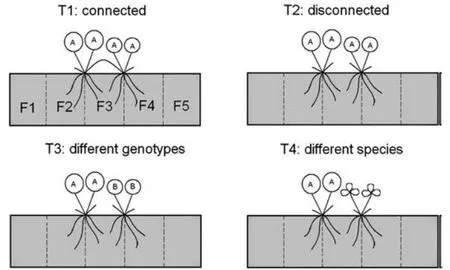
图 1 根系行为模式

表 1 与不同类型的邻居物种共生时,可能的根系生长策略和预测的根系布局模式
2.2.1 分配量 与非亲缘物种共生相比,与亲缘物种共生时,根系或地上部的竞争均有所减少[2,16]。共享一个花盆的美洲海滩芥(CakileedentulaL.)幼苗表现出明显的亲缘识别反应,同亲缘组的根分配量比非亲缘组的少。在凤仙花(ImpatienspallidaL.)与藜(ChenopodiumalbumL.)的共享盆栽实验中也得到了印证。可见,同缘和非亲缘物种之间存在根系分配量的差异最可能的原因就是根系具有亲缘识别的能力[16]。另外,拟南芥(Arabidopsis thalianaL.)的一项研究表明,即使在幼苗期,群体内的竞争互作也会改变群体表型[2],因此,对于同缘和非亲缘在根分配量上的差异,另一种可能的解释是同亲缘和非亲缘群体之间表型变异差异的竞争互作结果。
2.2.2 代谢 植物具有通过根系分泌物和挥发物识别亲缘关系的能力[2,8,17],并可根据邻居的遗传关系调整它们的生长、生物量分配和形态[3],这种可塑性代表了一种间接的遗传效应特例,它被定义为基因型对同种邻居表型的影响[18]。在许多植物中,根对同一生理个体的根的反应不同于对其他植物的根,例如,灌木蒿(ArtemisiatridentateL.)表现的自我—非自我的识别并不是与根系相关,而是在同亲缘植物受伤时,伤口分泌的挥发物要比面对非亲缘植物受伤时分泌的挥发物毒性低。
2.2.3 竞争回避 物种的竞争能力可以被看作是一种行为特征,它依据他们相邻物种的身份对自身个体进行能量分配管理,平衡资源竞争投入和繁殖投入之间的关系[19]。当个体频繁地遇到有亲缘关系和无亲缘关系的竞争对手时,该个体将得益于对亲缘的识别能力和对有亲缘关系个体间竞争的选择性回避能力,但这种能力在进化中有可能被限制[3]。例如,一些通过无性繁殖后代的多年生植物,将后代繁殖在附近,形成个体间基因型完全相同的群体,该物种具有非识别回避特征,这种特征使它在与所有邻居竞争时均处于优势地位,因为,它们与无亲缘关系的个体几乎不发生相互作用,亲缘选择特别强烈。选择性竞争回避排除了激烈的同胞竞争,提高了亲缘选择效率和直接适应性收益率,即投入到竞争器官用于增殖的资源小于用于繁殖的资源,因此,具有竞争回避能力的物种,产量较高。
3 邻居身份与环境互作影响的功能性状可塑性进化
不同的邻近物种可以被看作是一种可选的生物环境,与邻居的相互作用,取决于相互作用植物的身份对植物功能性状施加的选择压力。当邻居的身份在空间和时间上稳定时,由不同物种组成的植物邻居将选择特定的表型进行遗传分化和本地适应。如果邻居构成的空间和时间变异需要改变形态才能成功完成生存和繁殖时,那么植物对邻居身份的表型可塑性将经历选择压力。
在植物中,邻居身份的变异性很大程度上取决于物种的生活史特征和群落特征。由于有限的种子传播或繁殖的后代的空间聚集,邻近个体间可能主要是同基因型个体。随着群落物种丰富度的降低,增加了与任何特殊相邻物种相遇的可能性,而群落均匀度的降低,则使优势物种成为最可能的邻居。直接相邻物种组成的变化,导致了植物依据相邻物种身份改变形态的能力增强。专家预测,如果替代环境(如,与不同的邻近物种共生)很常见,并且以相似的频率出现,可塑性最有可能进化。植物对邻居身份的反应取决于其所在群落中测验的邻居被该目标物种遇到的频率,在与相邻种(同种和异种)同时发生且频率高而均匀的物种中,功能性状的可塑性最强。与邻近物种的互作可以促进功能性状可塑性选择,而功能性状的可塑性也可以通过性状分化和生态位分化来影响物种共存和群落结构[20]。
4 问题与展望
4.1 根系分泌物
研究已表明,根系分泌物可以传递关于邻居遗传身份的高度特异性线索,并在整个根系系统和与分泌物直接接触的个体根系水平上触发不同的反应。同物种根系分泌物引起的根系增生主要是通过形态调整实现的,而不是通过增加生物量投入实现的[8]。在博弈论模型中,将根系形态可塑性作为一种在竞争对手存在时调节根系生产成本和收益。现在已经明确根系分泌物作为邻居识别的一般机制,但还需要进一步的研究来确定根系分泌物中哪些化合物或化合物组合触发了观察到的这些反应。
4.2 化感物质与化学信号
与不同类型的临近根系一起生长的植物在不同物种之间表现出的根系行为模式是不同的,除获取资源外,邻近的根还能通过释放化感化学物质和无毒信号来广泛地影响植株的生长行为。它们对邻近植物生长的影响取决于根系分泌物中的化感物质浓度和释放的量、临近植物生育时期、生物和非生物环境条件等,在受控条件下,目标植物的生长期、生物量和密度是改变向根际释放的化感物质有效性的重要因素。在未来,应该对资源高效利用的群落物种进行更多的研究,估计这些群落中根系相互作用的重要性和后果。还需要进行更多的研究来确定与上述研究中观察到的根系行为模式相关的生活史和形态性状以及群落特征。
4.3 基质体积
不同物种在生长敏感性和向生殖器官生长量分配上,对基质体积的反应不同。“公地悲剧”和“自我—非自我”识别的实验得出的某些结果与基质体积有关。在分隔盆栽中,空间越有限,根系的化学自抑制能力越强,对土壤资源获取量减少,导致根系结构、形态和激素的变化;在未被分隔、存在两株根系竞争的盆栽中,基质体积越大,无效根分布量也越大。上述研究并不表明根系竞争中的公地悲剧不存在,而是表明它不是对所观察到的现象的唯一可能的解释。然而,目前还没有系统的研究表明根系“自我—非自我”识别导致根生产过剩的类似机制,也没有证据表明选择性压力会导致根系竞争中的公地悲剧。
4.4 相邻物种密度
研究表明,亲缘关系对植物性状的影响一直依赖于相邻密度。随着邻近植物密度的增加,有亲缘关系的植株用于种子繁殖的投入显著提高,在竞争器官上的生长分配减少,而在没有亲缘关系的个体之间没有表现出类似的反应。当这一现象发生时,形态和生物量分配的变化与亲缘选择理论是一致的,避免了与同亲缘物种的竞争。然而,基因亲缘关系的影响随着邻居密度的变化而变化,这表明竞争的激烈程度也可以改变由亲缘识别引发的反应[3]。因此,在未来,需要在同亲缘物种竞争频率方面进行更多的研究工作,收集充足的田间数据,以提高我们对亲缘识别进化条件的理解。
4.5 竞争回避与竞争忍耐度
系统地考虑与竞争抑制和耐受性相关的功能特征,可能会更好地理解生态学中的关键问题,如,控制植物群落构成因素[21,22]、生物入侵和对全球变化的反应[23]。研究指出,物种的竞争能力可以被看作是一种行为特征,它依据他们相邻物种的身份对自身个体进行能量分配管理,平衡资源竞争投入和繁殖投入之间的关系[19]。选择性竞争回避排除了同亲缘物种竞争,提高了亲缘选择效率和直接适应性收益率。虽然前人研究结果与进化博弈论模型的预测一致,但还需要对不同竞争和扩散策略的成本和收益进行进一步的实证研究,以提高我们对促进物种共存机制的理解。
在地下物种互作竞争中,成功的物种较少地依赖于竞争个体间的形态大小,而更多地依赖于竞争忍耐度的相关特征[9],如根系形态、分支和垂直分布等,我们需要超越快速增长等同于竞争优势的原有思维框架,认识到竞争忍耐度相关特征的重要性[10]。此外,一些竞争忍耐度的相关性状可能反映了植物对微生物的依赖。在今后,应当做出更多的努力来揭示这些联系。
4.6 功能性状可塑性进化
相邻物种的种类和群体数量的变化,导致了植物改变形态的能力增强。邻域相互作用与可塑性之间的关系可以依据生态进化动力学的框架来看待[24,25],与邻居的生态相互作用推动了功能性状可塑性进化,反过来也可能对生态相互作用和空间格局产生影响。现有研究已证明,群落模式和可塑性之间的重要联系,但需要进一步的研究来证明这种关系背后的因果关系,并确定可塑性的差异如何反馈到生态相互作用和影响物种共存上的。今后还需要对同一物种的多个种群和不同年龄的群落进行研究,以阐明对邻居多样性的本地适应效率。一些植物物种表现出根据其邻近区域的遗传多样性,在与生长和寿命相关的功能性状之间管理其资源分配,引起凋落物分解、养分循环和植物—土壤反馈,这一发现强调了在将遗传多样性与生态系统过程联系起来时,需考虑植物基因型的功能性变异,并结合群落和生态学相关知识,针对表型可塑性开展整合研究。
