植物体内活性氧的产生及清除机制研究进展
2021-04-24郭明欣刘佳佳侯琳琳张笑天刘含笑
郭明欣 刘佳佳 侯琳琳 张笑天 刘含笑
(洛阳师范学院<生命科学学院>,河南 洛阳 471934)
0 引言
干旱、盐碱、低温、重金属等非生物胁迫会导致植物细胞内活性氧(Reactive oxygen species,ROS)增加。由于ROS 的过量积累会产生氧化胁迫,因此,植物为了应对复杂的外界环境,在进化过程中形成了复杂的ROS 平衡调控机制。现有研究表明,植物对非生物胁迫的耐性与自身的抗氧化胁迫能力密切相关。本文对ROS 的产生、毒害及ROS 的清除进行综述。
1 植物体内活性氧的产生及毒害
在正常外界环境条件下,氧分子处于不太活泼的氧化状态。但是,当植物遭受病虫、干旱、盐碱、重金属、紫外光(UV)等生物或非生物胁迫时,植物体内电子传递和氧化还原动态平衡被破坏,细胞内产生的电子很容易使氧化态的氧转变为还原态的氧,产生超氧阴离子、羟自由基(OH-)、单线态氧(1O2)和过氧化氢(H2O2)等物质,这些物质被统称为活性氧。
1.1 活性氧的产生
研究表明,植物细胞的细胞壁、质膜、叶绿体、线粒体、 过氧化物酶体和内质网等部位都可产生ROS,其中叶绿体、线粒体、过氧化物酶体是ROS 产生的主要细胞器。
细胞壁: 植物细胞壁上存在多种产生ROS 的氧化酶,如过氧化物酶(Peroxidases, POXs),外界环境的刺激诱导POXs 产生ROS。ROS 的爆发在植物防御病原菌中发挥着重要作用[1]。另外,细胞壁的二胺氧化酶在醌的还原过程中也可以形成ROS。
细胞质膜: 植物细胞质膜上的还原型辅酶II(NADPH) 氧化酶是H2O2产生的主要来源。 该酶以NADPH 为电子供体催化胞外O2产生。NADPH 氧化酶在植物的生长发育及生物和非生物胁迫反应中起重要作用。
叶绿体:植物的叶绿体是ROS 产生的重要部位。叶绿体光合电子传递系统是ROS 的一个重要来源。在光系统I(PS I)中,叶绿素分子吸收能量后由基态上升到一个不稳定的、高能的激发态,在从激发态向较低能量状态转变的过程中会发生电子的漏渗,电子转移到O2产生单线态氧1O2。在光系统II(PS II)中激发的三线态叶绿素分子与O2相互作用也产生。UV 辐射或强光胁迫引起的光抑制作用也会导致叶绿体内1O2和的产生。
线粒体:线粒体呼吸作用过程中的电子传递链是ROS 产生的主要部位。线粒体呼吸链末端氧化酶把底物的电子传递到氧分子形成水或H2O2。 在哺乳动物中,大约有1-5% 的氧被消耗于ROS 的产生。在植物中,由于交替氧化酶(Alternative oxidase, AOX)的存在,ROS 在线粒体中处于一个比较低的水平。 在线粒体中,ROS 作为信号分子具有重要的调节功能, 调节植物的胁迫适应性及细胞的程序性死亡。
过氧化物酶体: 过氧化物酶体也是H2O2产生的主要细胞器。 在过氧化物酶体中发生的β-氧化伴随有H2O2的产生。另外,过氧化物酶体中的乙醇酸氧化酶能够把叶绿体光呼吸中产生的乙醇酸氧化生成乙醛酸和H2O2。
1.2 活性氧的毒害
ROS 在植物的生命周期中发挥着重要作用。ROS作为信号分子调控着植物的发育、 细胞程序性死亡、激素信号以及植物对生物和非生物胁迫的响应等过程[3]。然而,当植物体内ROS 积累过量时,就会产生氧化胁迫。过量积累的ROS 会破坏细胞的结构,损伤DNA、脂类、蛋白等生物大分子,并最终导致细胞死亡[4]。
ROS 对DNA 的损伤,主要包括碱基的缺失、修改(烷化,氧化)、嘧啶二聚体化、双链的断裂等。DNA 受损将影响基因组的稳定、蛋白的翻译、信号的转导等多个方面,并使细胞膜受到破坏,光合功能减弱,最终阻碍植物的正常生长发育[5]。
ROS 主要通过共价的修饰使蛋白发生氧化,一般以蛋白的羰基化水平作为蛋白氧化程度的评价指标。不同的氨基酸对ROS 敏感程度不同, 其中精氨酸(Arg)、赖氨酸(Lys)、脯氨酸(Pro)、苏氨酸(Thr)、色氨酸(Trp)对ROS 比较敏感。ROS 倾向于氧化含硫和巯基的氨基酸,像甲硫氨酸(Met)、半胱氨酸(Cys)。ROS能够与甲硫氨酸发生反应,形成硫自由基,该自由基与另一个硫自由基发生反应形成二硫键。此外,活性氧能够使甲硫氨酸的残基发生反应形成硫的衍生物。大部分蛋白发生氧化后是不能复性的。Bartoli et al.(2004)的研究发现,线粒体中蛋白的羰基化程度要高于叶绿体和过氧化物酶体,这表明线粒体对氧化胁迫更加的敏感[6]。
ROS 对细胞质膜的损害可以作为对细胞内脂类损害的一个评价指标。ROS 对质膜的损害可以分为三个步骤,包括起始、发展和终止。起始是指OH-与不饱和脂肪酸的亚甲基发生反应形成碳自由基(R-),启动膜质过氧化。发展是指R-没有配对电子,能够与基态氧反应,形成过氧化自由基(ROO-);ROO-再与其他脂肪酸发生反应生成羟过氧化物(ROOH)和R-,该R-又参与下一轮的反应。终止则为R- 与自身交联形成脂肪酸二聚体,ROO-自身交联形成过氧桥联二聚体(ROOR)[4]。
2 活性氧的清除
由于体内过量ROS 的积累会阻碍植物的生长发育甚至导致植株死亡,为了应对不断变化的外界环境,植物进化出一套复杂的过量ROS 清除机制。植物体内ROS 的清除机制分为两种,一种是酶促清除机制,主要包括超氧化物歧化酶(Superoxide dismutase,SOD)、抗坏血酸过氧化物酶(Ascorbate peroxidase,APX)、谷胱甘肽过氧化物酶(Glutathione peroxidase,GPX)、过氧化氢酶(Catalase,CAT)、谷胱甘肽S -转移酶(Glutathione S -transferases,GST)。另一种是非酶促清除机制,主要是依赖细胞中抗坏血酸 (VC)、 还原型谷胱甘肽 (Lglutathione,GSH)、α-生育酚(VE)、脯氨酸(Pro)、生物碱(Carotenoids)、类黄酮(Flavonoids)等清除ROS(见图1)。
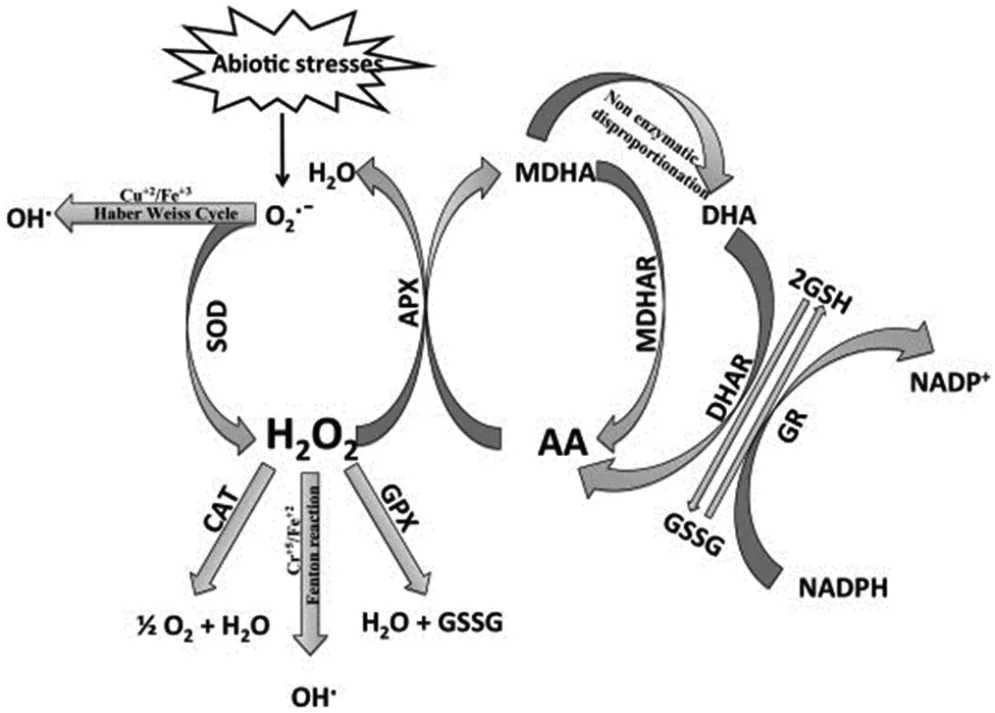
图1 植物体内活性氧清除机制[4]
2.1 酶促清除机制
SOD 是一种金属酶。 根据金属的差异,SOD 可分为Cu-Zn-SOD、Mn-SOD、Fe-SOD 三种类型。SOD 分布广泛,存在于线粒体、叶绿体、过氧化物酶体等细胞器以及细胞质基质中。 高等植物中SOD 以Cu-Zn-SOD 为主。SOD 能催化形成H2O2和O2, 降低ROS对植物的毒害。付畅等(2009)的研究发现将西伯利亚蓼的PsMnSOD 在酵母中表达提高了酵母在盐碱胁迫下的SOD 活性,并提高了酵母对盐碱胁迫的抗性[7]。
CAT 是植物细胞中清除ROS 的主要酶类之一,是唯一不需要能量的ROS 清除酶, 它催化H2O2形成H2O 和O2。CAT 主要存在于植物的过氧化物酶体和乙醛酸循环中。植物中的CAT 可以分为三类,第一类在光合作用组织中存在, 清除光合作用产生的H2O2;第二类清除乙醛酸循环中脂肪降解产生的H2O2;第三类存在于微管组织中,参与植物木质素的合成。在水稻基因组中已经发现有3 个编码CAT 基因, 分别是OsCATA、OsCATB、OsCATC(OsNOE1)。水分胁迫和外源ABA 能够诱导OsCATB 的表达,从而阻止H2O2的过量积累。 研究表明,OsCATC 功能丧失突变体noe1体内由于积累了过量的H2O2, 诱导了NO 的产生,而过量的NO 诱导了叶片的细胞死亡,使noe1 突变体表现为叶片早衰[8]。
POX 也能够将H2O2还原成H2O。POX 包括两类,即APX 和GPX。APX 通过抗坏血酸-谷胱甘肽循环(APX-GSH) 再生系统, 在单脱氢抗坏血酸还原酶(Mono-dehydroaseorbate reductase, MDAR)、脱氢抗坏血酸还原酶 (Dehydroaseothate reductase, DHAR)、谷胱甘肽还原酶 (Glutathione reductase, GR) 等的作用下,利用VC、NADPH、GSH 等抗氧化物,将H2O2还原成H2O。 水稻基因组中有8 个基因 (OsAPX1-OsAPX8)编码APXs。这8 个APXs 分布于细胞中的不同部位,包括叶绿体、线粒体、过氧化物酶体、细胞质。过量表达OsAPX1 增强水稻孕穗期的耐寒性。OsAPX8 的表达受到盐胁迫的诱导,而其他几个APXs的表达不受盐胁迫的影响。 作为POX 的另外一类,GPX 主要通过谷胱甘肽过氧化物酶循环体系,直接利用GSH 将H2O2还原为H2O。 生成的氧化型谷胱甘肽(GSSG) 在GR 和NADPH 的共同作用下重新转变为GSH。
谷胱甘肽S-转移酶 (Glutathione S -transferases,GST) 是一大类多功能的蛋白家族, 在植物中广泛存在。GST 催化具有亲电取代基的外源性化合物, 与内源GSH 发生反应。 许多外源化合物在生物转化第一相反应中极易形成某些生物活性中间产物,它们可与细胞生物大分子的重要成分发生共价结合,由此对机体造成损害。GSH 与其结合后可以防止这种共价结合。GST 具有清除ROS、 调节胁迫诱导的细胞程序性死亡等功能[4]。水稻基因组中有79 个GST 家族的成员, 分 为 以 下7 类:Tau、Phi、Zeta、DHAR、EF1G、Theta、TCHQD。Tau 家 族中OsGSTU3、OsGSTU4 的 表达受重金属和PEG 胁迫的诱导[9]。
2.2 非酶促清除机制
Vc 是一种在植物中大量存在的、 可溶性的抗氧化物质。所有的植物都能够合成Vc,其可以作为还原剂,直接与ROS 反应后清除ROS。另外,它也作为酶的底物在ROS 清除中扮演重要角色。VC 在叶绿体中含量丰富,是叶绿体中重要的抗氧化物质。外源的Vc能缓解盐胁迫产生的ROS 对小麦的损害。
GSH 是由Glu-Cys-Gyl 组成的三肽,是植物中重要的抗氧化剂,存在于植物的所有组织和细胞。在植物中,GSH 主要通过以下两种形式清除ROS。 第一,GSH 与1O2、HO-和O2-等直接发生化学反应, 清除ROS。 第二,GSH 作为供氢体, 通过与DHAR 反应使Vc 从氧化型转变为还原型,增加细胞中还原型Vc 的含量。
类黄酮(flavonoids)是植物中一类具有多种功能的次级代谢物质,主要包括黄酮醇、黄酮类、异黄铜、花色素等,在细胞内能够区隔化和清除ROS。在水稻中过量表达异黄酮还原酶基因OsIRL,增强了水稻的抗氧化胁迫能力[10]。
