Effects of temperature on the respiratory metabolism, feeding and expression of three heat shock protein genes in Anadara broughtonii*
2021-04-14XuxuLIUXiumeiZHANGYihangWANG
Xuxu LIU, Xiumei ZHANG , Yihang WANG
1 Key Laboratory of Mariculture, Ministry of Education, Ocean University of China, Qingdao 266003, China
2 Fisheries College, Zhejiang Ocean University, Zhoushan 316022, China
3 Laboratory for Marine Fisheries Science and Food Production Processes, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266237, China
Abstract Anadara broughtonii is one of the main commercially important species of shellfish in northern China. A. broughtonii lives in relatively stable subtidal zone where the clam could respond to environmental changes with minimum energy. Therefore, slight fluctuations in water environment may have a great impact on physiological processes such as feeding and metabolism. Large-scale mortality often occurs during the intermediate rearing processes, and high temperatures in summer are considered the leading cause of mortality. To understand the physiological and molecular response patterns of A. broughtonii at high temperatures and to estimate the appropriate metabolism temperature for A. broughtonii, the eff ects of temperature on the respiratory metabolism and food intake at diff erent growth stages were studied. The response patterns of heat shock protein genes to heat stress and post-stress recovery were also explored.Results show that the temperature and growth stage of A. broughtonii were two important factors aff ecting metabolism and feeding. The optimum temperature for feeding and physiological activities in each shelllength group was 24 °C. The temperature at which the peak metabolic rate occurred in each shell-length group increased with the increase in shell length. With the increase in heat stress, the expression of three heat shock protein genes ( Abhsp60, Abhsp70, and Abhsp90) in the tissues of two size groups of A. broughtonii increased significantly when temperature was above 24 °C and reached a peak at 30 °C. After heat shock at 30 °C for 2 h, the expression of the three heat shock protein genes rapidly increased, peaked at 2 h after the heat shock, and then gradually decreased to the level of the control group at 48 h after the heat shock. The three heat shock protein genes respond rapidly to heat stress and can be used as indicators to the cellular stress response in A. broughtonii under an environmental stress.
Keyword: Anadara broughtonii; temperature; oxygen consumption rate; clearance rate; heat shock protein gene
1 INTRODUCTION
China has a long history of bivalve mariculture by line-hanging and seed releasing on the sea bottom.The ark shell Anadara broughtonii is one of the most commercially important shellfish in northern China.A. broughtonii lives in the subtidal zone and is a typical filter-feeding shellfish that directly obtains food and oxygen from seawater. A relatively stable environment in subtidal zone is favourable for the species to tolerate environmental changes (Somero,2010). Therefore, slight fluctuations in the water environment (temperature, salinity, food supply, etc.)may have a greater impact on physiological processes such as feeding and metabolism. Cultured ark shells have suff ered heavy mortality in recent years,particularly in summer. As shown in experiments in Japan, survival was monitored in cages suspended from buoys and cages fixed to the seabed. Experiments began in June and after two months survival was 80%-90%. However, in the August there was a sudden massive mortality and by September the survival dropped to 15%-33% (Broom, 1985). In our investigation, A. broughtonii suff ered from a large number of deaths in August (maximum temperature:28.9 °C). Obviously, temperature played an important role in environmental stress as reported in previous studies on the large-scale mortality due to a rise in water temperature above 25 °C (Broom, 1985; Tang et al., 1994).
Temperature is one of the most important environmental factors aff ecting bivalves and is closely related to their feeding, energy utilization, and metabolism (Navarro et al., 2000). With the increase of ambient temperature, the metabolism consumption of shellfish increases. To adapt to the enhancement of metabolism, the feeding level of shellfish increased accordingly. However, when temperature continues to rise beyond the shellfish tolerable range, their metabolism and feeding are inhibited, and the shellfish would close its gape and stop the contact with the outside world. Feeding and metabolism are the key indicators that reflect the physiological status of shellfish. The gills, as well as the mantle, play an important role in respiration and feeding (Tang et al.,1994). The clearance rate is the volume of water completely cleared of particles per unit of time, which is used to reflect the feeding ability (Coughlan, 1969;Famme et al., 1986). The physiological activities of shellfish require a supply of energy. Aerobic metabolism is the main source of energy for shellfish,and the level of metabolism can be expressed by the amount of oxygen consumed by shellfish per unit of time (oxygen consumption rate) (Peck and Conway,2000; Peck et al., 2002; Goulletquer et al., 2004;Jansen et al., 2009; Brown et al., 2010).
The aquatic environment is a very complex ecosystem, and the health of aquatic animals is aff ected by environmental or biological factors such as water temperature, salinity, oxygen content,pollutants and bacterial pathogens. Bivalves are exposed to the substrate, so they are extremely vulnerable to changes in the environment of water and substrate. Due to their lack of mobility, it is diffi cult to avoid actively uncomfortable environments,and they can only adapt to the environment through their own homeostasis. As a type of stress protein,heat shock proteins (HSPs) can be triggered under many environmental stress conditions, such as high temperature, bacterial infection, hypoxia, and starvation. The function of heat shock proteins is to prevent proteins from forming wrong conformation or aggregation and to transport proteins across membranes in cells (Hartl and Hayer-Hartl, 2002;Kregel, 2002). By restoring the denatured proteins and degrading irreversibly damaged proteins, HSPs can help organisms to regulate stress responses and protect organisms from environmentally induced cellular damage (Feder and Hofmann, 1999). The expression of heat shock protein genes is closely related to the tolerance of organisms and the intensity of the environmental stress. The correlation between biological stress response and environmental stress intensity is one of the important ways of studying biological adaptability. The induction of gene expression of heat shock protein by heat and other forms of stress has been studied for decades (Sarge and Murphy, 1993). At present, the research focuses on the following aspects: the temperature threshold and limit temperature of hsp gene expression, the correlation between induced hsp gene expression concentration and environmental stress intensity, and the diversity of hsp gene expression under environmental stress. Relevant studies have been carried out in the shellfish of Crassostrea gigas,Crassostrea virginica, Mytilus edulis, etc. The results show that the expression of hsp genes changes significantly under the conditions of pathogen infection and environmental stress, and plays a protective role in cells and organism (Palmisano et al., 1999; Wang et al., 2007; Connoll et al., 2008). The heat shock protein family in molluscs not only plays a protective role on cells under heat stress, but also can synthesize and accumulate under other exogenous stimuli (such as heavy metal ions, DNA damage, free radicals, etc.). Therefore, the expression of the heat shock protein gene has been widely recognized and applied as an indicator of the biological response to adverse factors in the environment.
To understand the causes oflarge-scale deaths of A. broughtonii in summer and enhance the survival rate of A. broughtonii, researchers have done a lot of works, such as optimizing the intermediate culture and seeding technology, studying the reproductive development process, and comparing the growth state and habitat environment of the clam in diff erent sea areas (Min et al., 2004; Kim et al., 2007, 2008; Sedova et al., 2012). In addition, a number of factors that maylead to the death of the clam were also studied,including the eff ects of heavy metals and organic pollutants on the physiology and reproduction of the clam, the eff ects of temperature, salinity and hypoxia on the physiological state of the clam and its physiological response under stress (Watanabe et al.,2005; Kim et al., 2006; Shin et al., 2006; Shin et al.,2008; An et al., 2009; Lee et al., 2009; An and Choi,2010). To better understand the eff ects of temperature on the feeding and metabolism of A. broughtonii, as well as the eff ects of heat stress on the expression of Abhsp60, Abhsp70, and Abhsp90, we measured the clearance rate and oxygen consumption rate of A. broughtonii under various water temperatures and examined the expression patterns of the three heat shock protein genes under heat stress conditions.
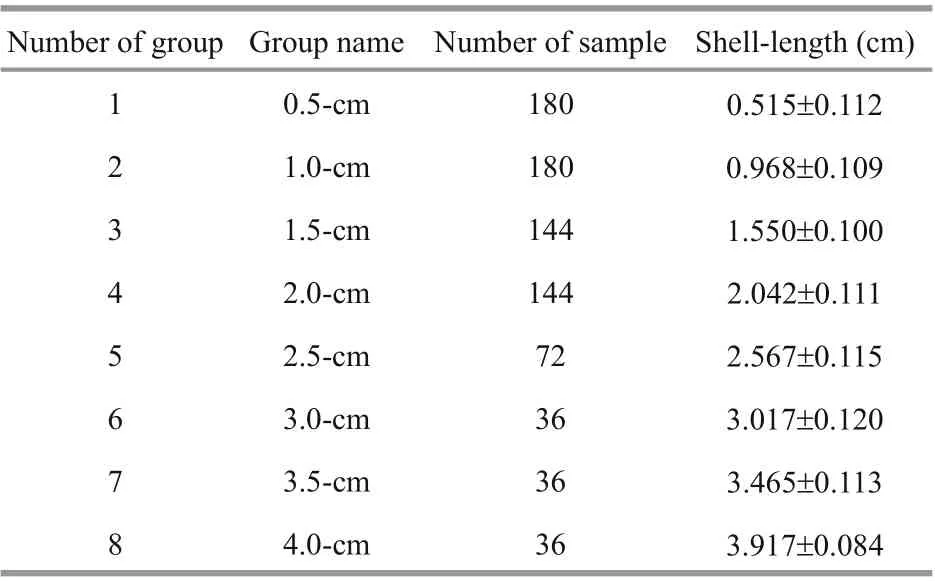
Table 1 Shell-length grouping of A. broughtonii used in this experiment
2 MATERIAL AND METHOD
2.1 Animal
Anadara broughtonii were collected from culture areas in Lingshan Bay, Qingdao, China. The individuals were placed in a damp plastic box with a layer of seawater ice and transported to laboratory.Upon arrival at the laboratory, they were acclimated for one week at the original ambient temperature to reduce the possible stress from transportation. The clams were maintained in chambers (40 cm×40 cm×50 cm) with aerated sand filtrated seawater at a salinity of 30. The clams were fed twice a day with algal slurry under a natural photoperiod.
2.2 Respiration and feeding experiment
2.2.1 Temperature acclimation
After the acclimatization, healthy individuals were selected for the experiment. The lengths of the clams should be measured first, to examine if there are variations on physiological parameters to be measured among clams with diff erent sizes, we defined 8 groups based on their shell-length ranges, named as 0.5, 1.0,1.5, 2.0, 2.5, 3.0, 3.5, and 4.0 cm. For example, the clam with the shell-length falling into 0.3 cm ≤ and≤ 0.7 cm range would belong to the 0.5-cm group. The grouping of the samples used in this experiment is shown in Table 1.We used six temperature levels (15, 18, 21, 24, 27,and 30 °C) to investigate the physiological variations under diff erent temperature. Therefore, each group would be further divided into six sets evenly and each set was cultured in seawater with pre-set temperature.The six temperature sets corresponded to the seasonal temperature range of the seawater in Lingshan Bay from May to November, during which 30 °C was the highest temperature in summer, while 15 °C is the suitable temperature at which A. broughtonii could have no stress (Cho et al., 2009). Before the start of experiments at each temperature, the clams were starved for 24 h (Widdows and Staff , 2006). The clearance rate and oxygen consumption rate were measured according to procedures reported by former researchers (Hawkins et al., 1996; Widdows and Staff ,2006).
2.2.2 Oxygen consumption rate
To measure the physiological responses of A. broughtonii, the clams were maintained in leakproof chambers. The experimental period was 4 h.Water samples were collected at the initiation and termination of the experimental period. Three replicates were used for each treatment in the experiments. The leak-proof chambers filled with filtered seawater but without organisms were used as blank control. The oxygen consumption rate (OR)was calculated using the following equation:
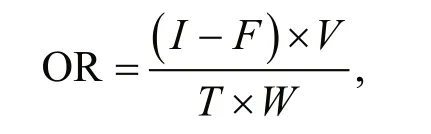
where I is the initial DO; fis the final DO; V is the volume of chamber; T is the time interval, and W is the wet weight of the shell.
2.2.3 Clearance rate
The methodologies for the feeding experiments were similar to those used in the respiration experiment. However, the clams used were diff erent from those used for the respiration experiment measurements to meet the assumption ofindependence(Underwood, 1997). The initial concentration of algal cells ( Cylindrotheca fusiformis) was 25 000 cells/mL.The water was sampled from every chamber at 1 h intervals over a period of 4 h. At the same time, the faeces were collected from the chambers, and the suspensions in the chambers were replaced by the initial concentration of algal suspensions. The control chambers, without A. broughtonii, showed no significant decline in cell concentration over the entire experimental period. Clearance rate (CR) was calculated using the following equation:
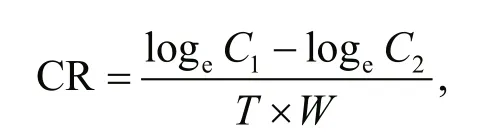
where C1and C2are the cell concentrations at the beginning and end of the experimental period; T is the time interval; and W is the wet weight.
2.2.4 Absorption effi ciency
Food absorption effi ciency (AE) was measured according to Conover (1966) by comparing the proportion of organic matter in the algal cells and faeces. Algal food and faecal pellets were collected on GF/C Whatman filters. The filters were ashed and pre-weighed with a balance before collecting the samples. The filters with the samples were dried at 90 °C, weighed, and ashed in a furnace at 450 °C for 5 h, and at last weighed again. The samples of algal food and faeces were collected from the chambers at hourly intervals over a 4-h feeding experiment period.Food absorption effi ciency (AE) was calculated using the following equation:
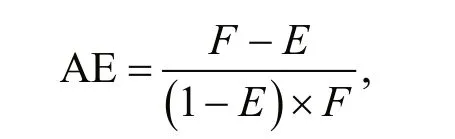
where fis the ash-free dry weight꞉dry weight ratio of the food, and E is the ash-free dry weight꞉dry weight ratio of the faeces.
2.3 Thermal stress experiment
2.3.1 Thermal stress
After 10 days of acclimation at 15 °C, clams of two sizes in length of 2.0 and 3.5 cm were chosen for experiments. To compare the expression diff erentiation under diff erent temperatures, clams were exposed to 18, 21, 24, 27, and 30 °C for 2 h. The control group was maintained at an ambient seawater temperature of 15 °C. The clams were sampled rapidly after thermal stress, and the gills and mantles of the clams were frozen in liquid nitrogen and stored at -80 °Cuntil use (5 biological replicates).To investigate the expression changes of these genes in diff erent treatment times under a fixed temperature, another group of the clams was exposed to a temperature of 30 °C for 2 h and then returned to the acclimation temperature (15 °C) for recovery. The clams were sampled at 0, 2, 4, 6, 8, 12, 24, and 48 h post treatment. The gills and mantles of the clams were frozen in liquid nitrogen and stored at -80 °C until use (5 biological replicates). The control group was maintained at an ambient seawater temperature of 15 °C during the whole experimental period.
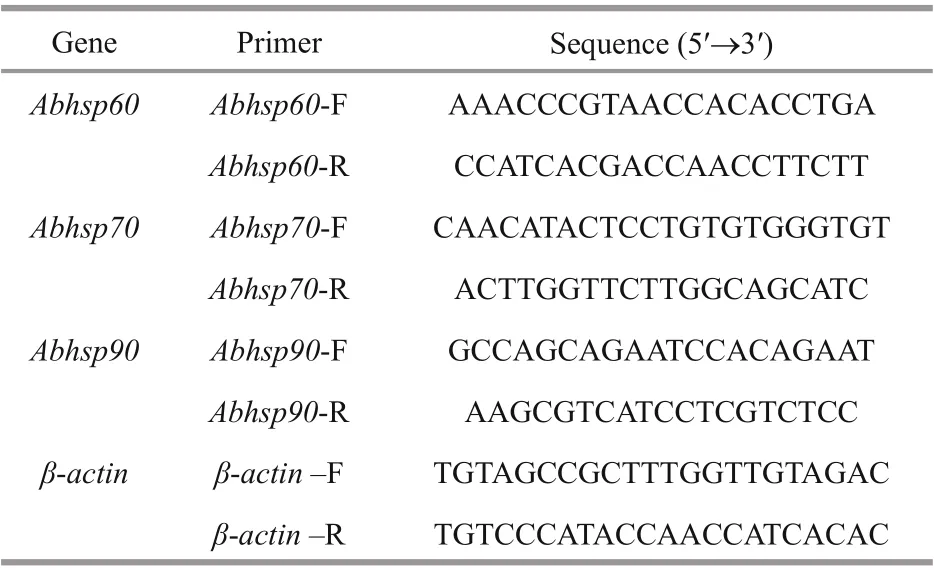
Table 2 Primer information
2.3.2 mRNA expression analysis by real-time quantitative PCR
Total RNA was extracted from the gills and mantles of shell with Trizol (TransGen) following the manufacturer’s instructions. The expression levels of diff erent transcripts of Abhsp60, Abhsp70, and Abhsp90 in tissues were detected by qRT-PCR to compare the control and the treatments (5 biological replicates). The RNA concentration and purity were determined by a Nanodrop 2000 spectrophotometer(Nanodrop Technologies) at the ratio of absorbance at 260/280 nm as an indicator. RNA quality was confirmed by gel electrophoresis. The first-strand cDNA was synthesized using the TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix(TransGen) as a qRT-PCR template. The specific primers for Abhsp60, Abhsp70, and Abhsp90 were designed based on the obtained sequences of the open reading frames (ORF) using Primer 5.0 (Table 2). The β-actin gene (ACTB) was used as the housekeeping gene (Huang et al., 2015). The relative transcript levels of the three heat shock responsive genes were determined using TransStart®Green qPCR SuperMix(TransGen) with a StepOnePlus (ABI Inc.). Each sample was run in three technical repeats.
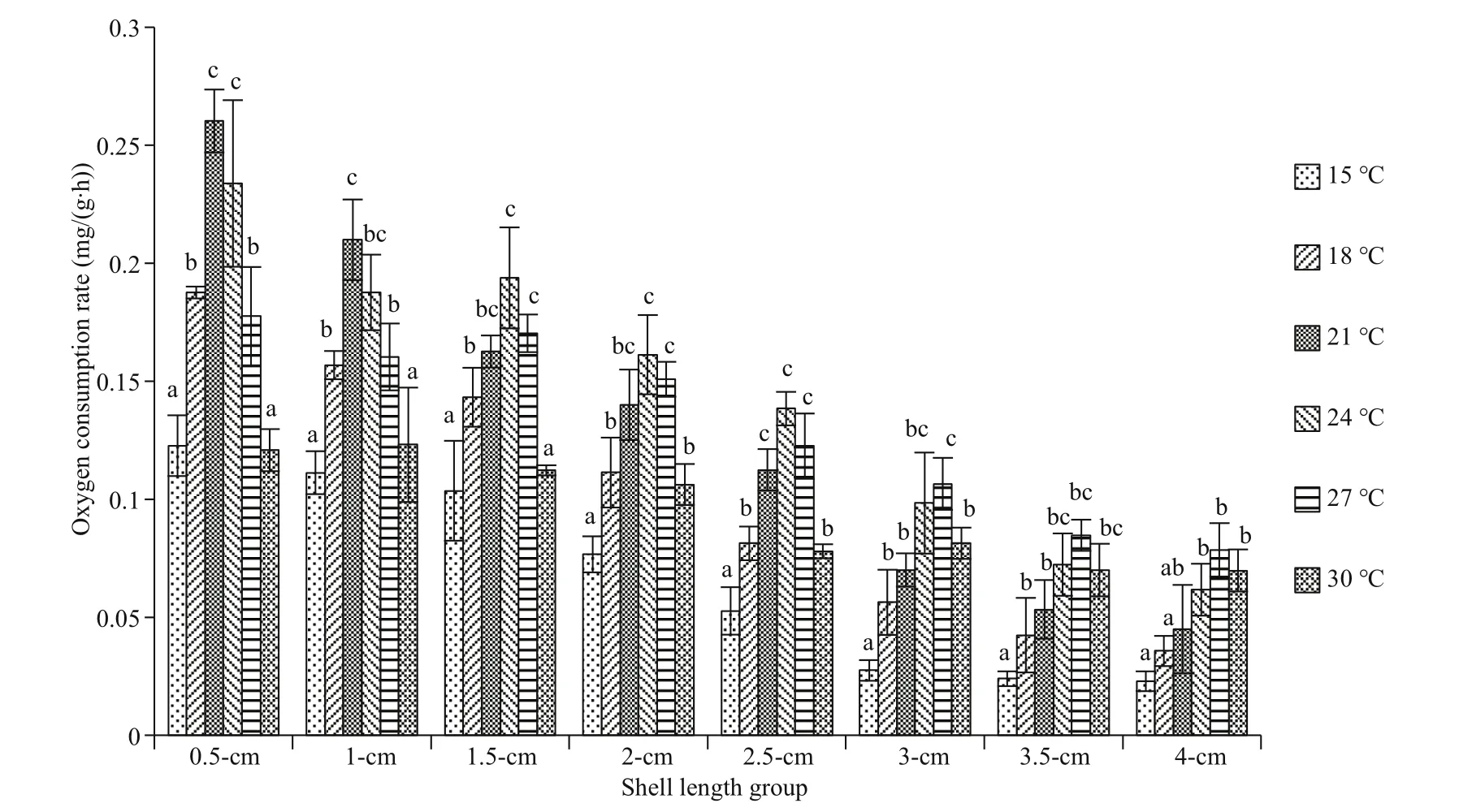
Fig.1 The oxygen consumption rate of diff erent arkshell shell-length groups under diff erent temperatures
2.4 Statistical analysis
The 2-ΔΔCtmethod was used to analyze the relative transcript expression levels of three heat shock response genes in the gills and mantles of the clams.The expressions of the 15 °C-set were used as controls. Values were normalized against the β-Actin transcripts. The results of the respiration, feeding and thermal stress experiments were statistically analyzed using one-way analysis of variance (ANOVA), and the relationships among physiological parameters,temperature, and shell length were evaluated with the Pearson correlation. The level of statistical significance was set at P <0.05 unless otherwise stated.The 3D surface plots and the corresponding two-order polynomial equations were obtained through the simulation and regression analysis of numerical results by Matlab and SPSS.
3 RESULT
3.1 The oxygen consumption rate of the clams at diff erent temperatures
In this study, the maximum oxygen consumption rate was 0.26 mg/(g∙h), which appeared in the treatment of the 0.5-cm group at 21 °C, and the minimum was 0.023 mg/(g∙h), which appeared in the 4.0-cm group at 15 °C (Fig.1). At a certain temperature,the oxygen consumption rate of A. broughtonii gradually decreased as the shell length increased.However, in the same shell-length group, with the gradual increase in temperature, the oxygen consumption rate showed a trend ofincreasing first and then decreasing. For diff erent shell-length groups,the temperature at which the peak oxygen consumption rate occurred was diff erent. The peak oxygen consumption rates of the 0.5-cm (0.26 mg/(g∙h)) and 1.0-cm (0.21 mg/(g∙h)) groups occurred at 21 °C. For the 1.5-cm (0.194 mg/(g∙h)), 2.0-cm (0.161 mg/(g∙h)),and 2.5-cm (0.138 mg/(g∙h)) groups, the peak values appeared at 24 °C. For the 3.0-cm (0.107 mg/(g∙h)),3.5-cm (0.085 mg/(g∙h)), and 4.0-cm (0.079 mg/(g∙h))groups, the peak values appeared at 27 °C.
3.2 The clearance rate of the clams at diff erent temperatures
The maximum clearance rate of this study was 3.47 L/(g∙h) in the 0.5-cm group at 21 °C, and the minimum clearance rate was 0.25 L/(g∙h) in the 4.0-cm group at 15 °C (Fig.2). At a given temperature, the clearance rate showed a gradual decreasing trend with the increase in shell length. At a constant shell length,with a gradual increase in temperature, the filtration rate first increased and then decreased, and the peak clearance rate of each shell-length group appeared at 24 °C.
3.3 The absorption effi ciency of the clams at diff erent temperatures
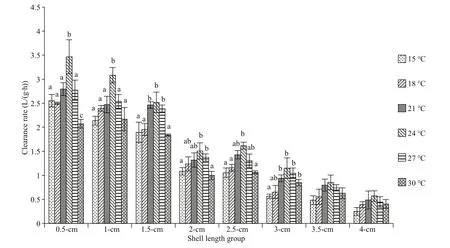
Fig.2 The clearance rate of diff erent arkshell shell-length groups under diff erent temperatures
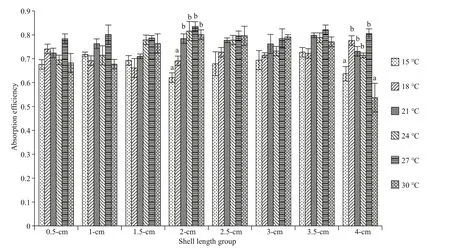
Fig.3 The absorption effi ciency of diff erent arkshell shell-length groups arkshell under diff erent temperatures
Under diff erent temperature conditions, the absorption effi ciency of each shell-length group fluctuated within the range of 53.7% to 83.5%, and there was no obvious change trend (Fig.3). One-way analysis of variance showed that the absorption effi ciencies of each shell-length group were not significantly diff erent at the same temperature( P >0.05). In the same shell-length group, as the temperature increased, the absorption effi ciency showed a certain degree ofincrease and then a decrease.
3.4 Relationship between oxygen consumption rate and clearance rate with temperature and shell length
According to the oxygen consumption rate and clearance rate of each shell-length group at diff erent temperatures, the relationship between the oxygen consumption rate (OR) or clearance rate (CR) with the temperature and shell length was obtained.
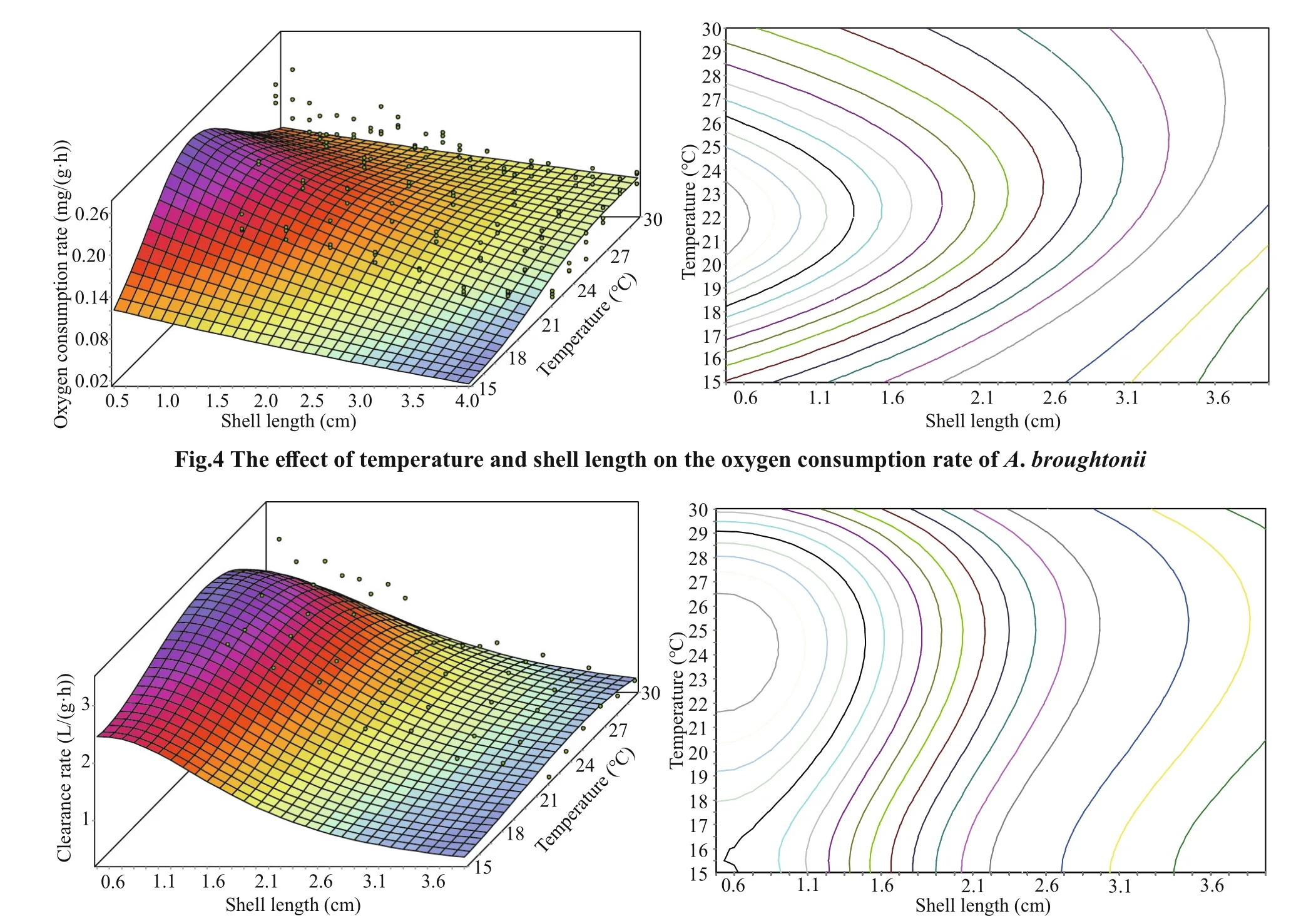
Fig.5 The eff ect of temperature and shell length on the clearance rate of A. broughtonii
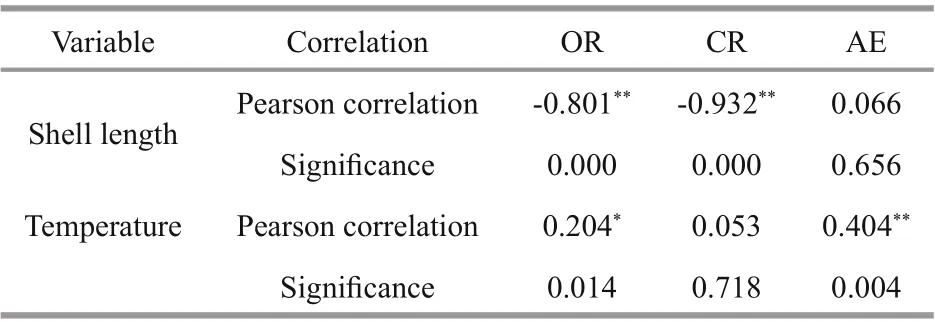
Table 3 Correlation between physiological parameters and temperature and shell length of A. broughtonii

where L is shell length, and T is the temperature. The root mean square error was 0.163 1. The R value was 0.981. The goodness of fit was 0.962 4 (Fig.5).
The Pearson correlation coeffi cients show that the oxygen consumption rate of A. broughtonii was negatively correlated with shell length significantly but positively with temperature significantly; that the clearance rate was negatively correlated with the shell length significantly; and that the absorption effi ciency was positively correlated with temperature significantly (Table 3).
3.5 The expression of three hsp genes of A. broughtonii under heat stress at diff erent temperatures
The expression of Abhsp60, Abhsp70, and Abhsp90 in tissues of A. broughtonii is shown in Fig.6. After 2 h of heat stress, the expression of the three heat shock genes increased with temperature increase.When the stimulation temperature was higher than 27 °C, the expression of the three heat shock genes in each tissue was significantly higher than that of the control group. After heat shock at 30 °C, the expression level of the three heat shock genes in each tissue was approximately 20-50 times that of the control group. There was a significant difference in Abhsp70 expression in the mantle and Abhsp90 expression in the gills between the two tested shelllength groups.
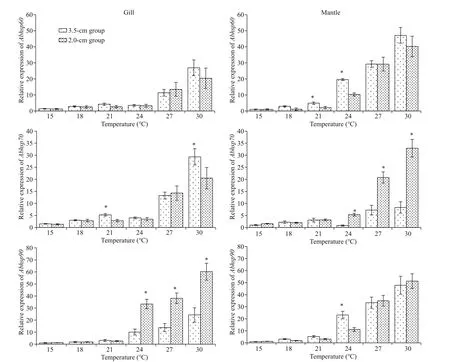
Fig.6 Transcript levels of three hsp genes of A. broughtonii under heat stress at diff erent temperatures
3.6 The expression of three hsp genes of A.broughtonii during recovery after exposure to 30 °C
The expression of the three heat shock genes in the tissues of A. broughtonii during the recovery process after heat shock at 30 °C for 2 h is shown in Fig.7. The expression of the three heat shock genes showed a similar trend. The expression rose rapidly after heat shock and reached its peak at 2 h after heat shock and then gradually decreased to the level of the control group at 24 h after heat shock. In the recovery process,there was a significant difference in Abhsp70 expression in the two tissues and in Abhsp90 expression in the gill between the two shell-length groups.
4 DISCUSSION
4.1 Eff ects of temperature on the respiratory metabolism of A. broughtonii
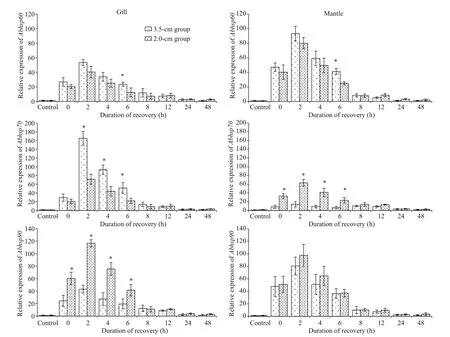
Fig.7 Transcript levels of three hsp genes in A. broughtonii after exposure to 30 °C for 2 h and recovery at 15 °C for diff erent durations
The oxygen consumption rate refers to the amount of oxygen consumed by shellfish per unit of time,reflecting the intensity of physiological metabolic activity in shellfish. The results indicate that, in this experiment, when the temperature was lower (15 °C)in the range of 15-30 °C, the oxygen consumption rate of A. broughtonii was lower too. The oxygen consumption rate gradually increased with the temperature. At the same time, when the temperature was higher than 24 °C or 27 °C, the oxygen consumption rate of each shell-length group decreased. This was due to the increase in the water temperature and the acceleration of biochemical reactions in organisms that resulted in a corresponding increase in the oxygen consumption rate, which is a common feature of poikilothermal animals (Aldridge et al., 1995; Bougrier et al., 1995; Gosling, 2015).However, as the temperature continued to rise, the homeostasis of shellfish was destroyed, causing harmful physiological reactions and inhibiting normal metabolic activities. When the temperature went beyond a certain limit, the oxygen consumption rate could decrease rapidly (Fan, 2010). However,this trend in oxygen consumption rate may vary due to unsuitable experimental conditions, the incomplete domestication of experimental animals, and the setting of the temperature range in experimental designs (Rodhouse, 1978; Widdows, 1978; Kim et al., 1980; Bougrier et al., 1995). A large number of studies have shown that the oxygen consumption rate of shellfish changed at a turning point when water temperature rose. Anestis et al. (2008) found that the metabolism and molecular response of Modiolus barbatus reached a maximum at 24 °C. Laing et al.(1987) found that the turning point of the change in the oxygen consumption rate of Mercenaria mercenaria was 25 °C. When seawater temperature for Musculista senhousia was higher than 30 °C its oxygen consumption rate began to decrease rapidly(Inoue and Yamamuro, 2000). In this study, we found that, as the temperature increased, the oxygen consumption rate of A. broughtonii in each shelllength group increased first and then decreased. The oxygen consumption rate in the 0.5-cm and 1.0-cm groups reached a peak value at 21 °C, so did in the 1.5-cm and 2.5-cm groups at 24 °C, and in the 3.0-cm and 4.0-cm groups at 27 °C. When the temperature rose to 30 °C, the oxygen consumption rates of the 0.5-cm to 3.0-cm groups were significantly lower than their maximum oxygen consumption rates. The respiratory metabolic activities of A. broughtonii were inhibited when water temperature was 30 °C,which was beyond the suitable range for its growth and survival.
4.2 Eff ects of temperature on the feeding physiology of A. broughtonii
In the study of shellfish feeding physiology, the clearance rate can reflect the feeding level of shellfish.According to the Pearson correlation coeffi cient analysis (Table 3), temperature variation had a significant eff ect on the clearance rate of A. broughtonii( P <0.01) from 15 °C to 30 °C. In the experiment, as the water temperature increased, the clearance rate of A. broughtonii first increased and then began to decrease after reaching a peak at 24 °C (Fig.2). It was concluded that the physiological metabolic activities increased with the increase of the temperature, and all physiological activities, including feeding, were inhibited when the temperature exceeded its optimum range. Jørgensen et al. (1990) suggested that temperature might aff ect the swing of cilia on the upper gill of shellfish, and the swing frequency increased with increasing temperature. When temperature exceeded the suitable range, the metabolic state of the shellfish changed, and more energy was needed to adapt to the changes in the external environment, resulting in a decrease in the vitality of the filter feeding organs and a decrease in the clearance rate. This phenomenon has been found in the studies of many shellfish, such as Ruditapes philippinarum(Wu et al., 2002), Sinonovacula constricta (Pan et al.,2002), Solenaia oleivora (Xu et al., 2005), M. edulis(Schulte, 1975), Dreissena polymorpha (Aldridge et al., 1995), etc. As the Pearson correlation coeffi cient shown in Table 3, the correlation coeffi cient between clearance rate and temperature (0.053) was lower than that between the oxygen consumption rate and temperature (0.204), indicating that the influence of temperature on the clearance rate was weak. The results suggest that temperature had a significant eff ect on respiratory metabolism but had a little eff ect on feeding behavior. Navarro and Gonzalez (1998) found that there was no significant difference in clearance rate between 16 °C and 20 °C for Argopecten purpuratus. Hutchinson and Hawkins (1992) found that temperature had a little eff ect on the clearance rate of Ostrea edulis L. in the temperature range of 15 °C to 25 °C; only when the temperature was below 15 °C,the feeding rate was significantly reduced.
Absorption effi ciency tends to evaluate the ability of shellfish to convert acquired food into their own nutrients. The absorption effi ciency of A. broughtonii was significantly correlated with the temperature(Table 3), but the variation in the absorption effi ciency was not significant at each temperature set (Fig.3).Dong et al. (2000) suggested that in a suitable temperature range, the digestive enzyme activity of Ruditapes philippinarum increased with increasing temperature, the absorption intensity of the intake bait increased, and the absorption effi ciency increased.However, when temperature was extremely higher or lower, the activity of digestive enzymes in clams could be inhibited, causing the decline of absorption effi ciency. In a similar research, Winter (1969) found that the absorption effi ciencies of Arctica islandica and Modiolus modiolus were not aff ected under 4 °C to 12 °C, but increased at 20 °C by 16% and 15%,respectively. Elvin and Gonor (1979) found that the absorption effi ciency of California mussels at 15 °C was twice as that of 9 °C. Albentosa et al. (1994)found that the absorption effi ciency of Venerupis pullastra increased by 17% when temperature increased from 10 °C to 20 °C. Therefore, the absorption effi ciency of bivalves is susceptible to changes in water temperature, and the degree ofinfluence depends on the species.
4.3 Eff ects of temperature on the metabolism and feeding characteristics of diff erent shell-length groups of A. broughtonii
According to the Pearson correlation analysis, the size of an A. broughtonii individual has a significant correlation with the oxygen consumption rate and the clearance rate (Table 3). By comparing the clearance rate and oxygen consumption rate of diff erent shelllength groups, the oxygen consumption rate and filtration rate of the 0.5-cm group were significantly higher than those of the 4.0-cm group at all experimental temperatures, which had also been reported in other shellfish studies. In Trachycardium flavum, the small individuals (shell height 27.2 mm)have higher oxygen consumption per unit weight(Jiang, 2012). Three diff erent sizes of clams (13-23 mm, 28-35 mm, and 41-46 mm shell length) were studied under same environmental conditions, and the clams of 13-23 mm shell length had the highest oxygen consumption rates (Suja, 2007). During the growth of shellfish, the proportion of each organ would constantly change due to allometric growth. As the organism size gradual increased, the proportion of the gills decreased, resulting in the reduction of the metabolic rate oflarge individuals (Riisgård, 2001).With the growth of shellfish, their metabolic rate decreased, and the demand for energy intake decreased too. In addition, shellfish feeding was accomplished by cilia movement on the gill; the clearance rate is proportional to the number of cilia, and the number of cilia is proportional to the surface area of the gill filament. The relative surface area of gill filaments decreased with the growth of shellfish (Jones et al.,1992). The filtration rate declines more rapidly than metabolic rate, then the growth effi ciency decreases as individuals grow, which may ultimately limit the size of bivalves (Gosling, 2015). Sukhotin suggested that compared with small mussels, large ones were more sensitive to low salinity and high temperature,and thus had higher metabolic requirements, causing bigger mortality under stress (Sukhotin et al., 2003).
4.4 Eff ects of temperature stress on the expression of three hsp genes in A. broughtonii
In this study, the expression of three heat shock genes showed tissue-specific, the degree of expression change in tissue might be related to the specific function of this hsp gene (Xu et al., 2014). Despite the tissue-specific expression of Abhsp60, Abhsp70, and Abhsp90, the expression patterns of the three diff erent heat shock genes in diff erent tissues of A. broughtonii were the same under diff erent heat shock temperatures(18, 21, 24, 27, and 30 °C). The relative expression of Abhsp60, Abhsp70, and Abhsp90 increased with increasing heat shock temperature (Fig.6). When the heat shock temperature reached 24 °C, the heat shock gene in each tissue began to be highly expressed. The relative expressions of Abhsp60 and Abhsp90 were significantly higher than that of the control group( P <0.05) after 2 h of heat shock at 24, 27, and 30 °C,while the expression of Abhsp70 increased significantly at 27 °C and 30 °C only. The expression levels of the three heat shock genes reached a maximum after heat shock at 30 °C, which was 20-50 times of the control group ( P <0.05). Zhang et al.(2009) found that the expression of hsp70 gene in the gill, heart, brain, liver, and other tissues increased significantly after heat stimulation. Qiang et al. (2012)found that temperature stress had a significant eff ect on the expression of hsp70 gene in the liver of tilapia.Wu et al. (2014) found that hsp70 gene in the mussel tissue was significantly increased under temperature stress. These results suggest that hsp genes can be expressed rapidly and abundantly in response to extreme temperature stimulation. In this experiment,there were differences in the expression levels of the three heat shock genes due probably to the diff erent structures and functions of the diff erent heat shock genes, which have diff erent functions in the process of cell damage repair. Heat shock proteins such as HSP60, HSP70, and HSP90 function similarly in diff erent tissues; they function independently as molecular chaperones and also work together (Fink and Hedera, 1999; Fan et al., 2003). Among them,HSP70 is involved in protein folding and prevents the misfolding of proteins (Nollen and Morimoto, 2002);HSP90 is mainly involved in the conformational regulation of certain transcription factors, and protein kinases (Harris et al., 2004); and HSP60 can rapidly transfer from the cytoplasm to the mitochondrial matrix under the action of HSP70 to repair denatured proteins in the mitochondrial matrix (Itoh et al.,2002). The high expression of hsp genes in a short period can rapidly inhibit the aggregation of denatured proteins and promote the refolding or degradation of denatured proteins, thereby protecting the normal synthesis ofintracellular proteins and providing eff ective protection for cells. Each HSP fully utilizes its respective functions to maintain the homeostasis of the cellular protein environment (Georgopoulos and Welch, 1993; Kiang and Tsokos, 1998). Generally,the expression patterns of HSP60, HSP70, and HSP90 under heat shock were similar, which should be due to the combination of HSP60, HSP70, and HSP90 in assisting proteins folding. The HSP70 of Sarcophaga crassipalpis was responsive to hypoxia, but HSP90 showed a little response; in dehydrating, rehydrating,and heat-shocked larvae of African chironomid( Polypedilum vanderplanki), six heat shock proteins,including HSP60, HSP70, and HSP90, were all upregulated; during the progression of chronic obstructive pulmonary disease, HSP60 expression increased but not HSP70 and HSP90 (Xu et al., 2014).Brun et al. (2008) suggested that HSP responses of organisms to stresses may be related to their habitat in the natural environment and caused their diff erential capacities for adaptation to environmental change.Combined with above researches, we could find the complication of HSPs expression of diff erent organisms responding to various stresses (Xu et al., 2014).
In the recovery experiment after heat shock, the Abhsp60 gene in each tissue was rapidly expressed after heat shock, reached its expression peak at 2 h after heat shock ( P <0.05) and then decreased gradually to the level of the control group at 24 h( P >0.05). The expression of the Abhsp70 and Abhsp90 genes during heat shock recovery was similar to that of Abhsp60 (Fig.7). In the study of Hong Kong giant oysters, the transcription level of hsp70 gene in each tissue after heat shock was significantly higher than that of the control group 3-6 h after heat shock and returned to normal levels within 12-24 h. As molecular chaperones, HSP60, HSP70, and HSP90 are capable of transcriptionally translating and refolding denatured proteins and preventing their aggregation after heat stress (Morimoto and Santoro, 1998; Feder and Hofmann, 1999). The continuous synthesis of heat shock proteins requires an energy supply, and their overexpression will aff ect the synthesis of other proteins, even leading to the stagnation of biological growth (Krebs and Feder, 1997; Viant et al., 2003).The transcription levels of the three heat shock proteins increased continuously during the first 2 h of the heat shock recovery stage, decreased during the recovery from heat shock and finally reached normal levels. This trend may be one of the strategies by which the clams maintain a balance between cell repair and energy metabolism under heat stress.
In this study, the expressions of Abhsp70 in the mantle and Abhsp90 in the gills of small individuals were significantly higher than that oflarge individuals under heat stress. In the recovery process, the expressions of Abhsp70 in the mantle and Abhsp90 in the gills of small individuals were significantly higher than that oflarge individuals. However, the expression of Abhsp70 in the gill oflarge individuals was significantly higher than that of small individuals.Husmann et al. (2014) found that younger Antarctic bivalve ( Laternula elliptica) had the most significant difference in transcriptional expression under stress,indicating that age was an important factor in transcriptional defense response. Philipp et al. (2011)found that the older L. elliptica was more sensitive to environmental stress and water temperature fluctuation,and the ability of reburrow into the sediment was more limited (Peck et al., 2007). Older individuals also showed lower antioxidant defense and higher levels of oxidative damage (Philipp et al., 2005). The significant changes in gene expression in young individuals after stress are consistent with the physiological differences caused by age of marine bivalves (Philipp and Abele,2010). In summary, younger animals have better chances of survival and recovery than the older ones under an environmental stress.
5 CONCLUSION
The clearance rate and oxygen consumption rate of A. broughtonii under various water temperatures showed that an excessive temperature would decrease clearance rate and oxygen consumption rate; and the optimum temperature for the physiological activities of each body length group was approximately 24 °C.The excessive temperature could also lead to significant expression of hsp gene. When the heat shock temperature was higher than 24 °C, Abhsp60,Abhsp70, and Abhsp90 were expressed significantly.These three heat shock protein genes responded rapidly to heat stress and could be used as indicators of the cellular response in A. broughtonii to an environmental stress. The persistently high temperatures in summer could cause high mortality due probably to the high ambient temperature above 24 °C, and the ability of stress response exceeded the limit of normal metabolism. Over time, with the stress intensity gradually increases and the clams eventually enter the apoptotic process, leading to a high mortality.
6 DATA AVAILABILITY STATEMENT
All data included in this research are available from the corresponding author on reasonable request.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Predicting sediment flux from continental shelfislands,southeastern China*
- Laboratory simulation of dissolved oxygen reduction and ammonia nitrogen generation in the decay stage of harmful algae bloom*
- Development of high-resolution chloroplast markers for intraspecific phylogeographic studies of Phaeocystis globosa*
- Effects ofiron and humic acid on competition between Microcystis aeruginosa and Scenedesmus obliquus revealed by HPLC analysis of pigments*
- Effect of river plume on phytoplankton community structure in Zhujiang River estuary*
- Exploring the sublethal genotoxic effects of class II organophosphorus insecticide quinalphos on freshwater fish Cyprinus carpio