Development of high-resolution chloroplast markers for intraspecific phylogeographic studies of Phaeocystis globosa*
2021-04-14QingchunZHANGZhuangNIUJinxiuWANGChaoLIUFanzhouKONGXiaokunHUJiayuZHAORenchengYU
Qingchun ZHANG , Zhuang NIU ,4, Jinxiu WANG ,4, Chao LIU ,4,Fanzhou KONG , Xiaokun HU,4, Jiayu ZHAO ,4, Rencheng YU ,4,
1 CAS Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences,Qingdao 266071, China
2 Laboratory for Marine Ecology and Environmental Sciences, Pilot National Laboratory for Marine Science and Technology(Qingdao), Qingdao 266237, China
3 Center of Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China
4 University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Phaeocystis globosa is an important harmful algal bloom causative species distributing widely in temperate and tropical coastal waters in the world. The morphological, physiological, and biochemical characteristics are diff erent among geographic strains, which can not be distinguished with nuclear ribosomal DNA markers at present. Therefore, the genetic distance and phylogeographic relationships of nuclear 28S rDNA D1-D2 and ITS regions, and three chloroplast intergenic spacers ( pet N-t rn S1, trn M1- psb A, and rbc S- rpl 27) were analyzed and compared among 13 strains of P. globosa isolated from the Pacific Ocean and Atlantic Ocean in this study. In addition, the nucleotide polymorphisms of 28S rDNA D1-D2, ITS, and rbc S- rpl 27 regions were evaluated in two P. globosa strains. The various levels of nucleotide polymorphism were in the nuclear 28S rDNA D1-D2 region and ITS region, but no polymorphism was in the chloroplast rbc S- rpl 27 intergenic spacer. A reasonable intraspecific phylogeographic relationship was presented by rbc S- rpl 27 intergenic spacer, which had the strongest distinction to geographic strains compared to those of 28S rDNA D1-D2 and ITS regions. In the phylogenetic tree of rbc S- rpl 27 intergenic spacer, the two strains from the North Sea of the Atlantic Ocean were divided firstly from the species of P. globosa, and then formed an independent clade, while the other Atlantic strains and all of Pacific strains joined up to build the other clade. It was implied that at least two genetically distant populations of P. globosa existed in the Atlantic coastal regions. This study provided a high-resolution chloroplast marker to analyze intraspecific phylogeographic populations of P. globosa, and preliminarily clarified the genetic relationships of the Pacific and Atlantic strains of P. globosa.
Keyword: Phaeocystis globosa; chloroplast; DNA marker; phylogeny
1 INTRODUCTION
Genus Phaeocystis, a major contributor of dimethylsulfide propionate (DMSP) enzymatically converted into dimethylsulfide (DMS) and acrylate(Stefels and Van Boekel, 1993; Stefels et al., 1995), is not only an important primary producer distributing from the Antarctic waters to the Arctic Ocean, but also an ecologically important member of phytoplankton with a significant role in biogeochemical cycles, including the global sulphur and carbon cycles (Charlson et al., 1987; Liss et al.,1994; Wassmann et al., 2005; Whipple et al., 2005a).Phaeocystis contains some causative species of harmful algal blooms (HABs), during which ichthyotoxic and haemolytic substances can be often released with detrimental impacts on growth and reproduction of shellfish and macrozooplankton(Davidson and Marchant, 1992; Shen et al., 2004; Liu et al., 2007). To date, there are 10 species of the genus Phaeocystis recorded in the AlgaeBase (http://algaebase.org/), and three of them, P. antarctica, P.globosa, and P. pouchetii, are reported as the bloomforming species. P. antarctica and P. pouchetii are cold-water species prevailing in the Antarctic and Arctic waters, respectively. While P. globosa is a warm-water species blooming in the nutrient-enrich(mostly nitrates) coastal areas of the north Atlantic,the North Sea, the Norwegian Sea, the Dutch coastal waters, the Arabian Sea, and the Southeast Asian including China, Vietnam, Thailand, etc. (Lancelot et al., 1987; Cadée, 1996; Qi et al., 2004; Schoemann et al., 2005; Hai et al., 2010; Rousseau et al., 2013;Smith et al., 2014b). Extensive research interests are focused on the species of P. globosa not only for its wide distribution, but also for its inclination to form gelatinous colonies encased by polysaccharide envelopes under suitable environmental conditions(Zingone et al., 1999; Jacobsen, 2002). Thousands of single cells are embedded in a colony in size from several millimeters (Rousseau et al., 1994; Peperzak et al., 2000), even to 3 centimeters in the blooms in China (Chen et al., 2002; Shen et al., 2004; Smith et al., 2014b). Therefore, their negative eff ects are indirectly resulted from the high abundance of colony cells and oxygen depletion upon its degradation in marine ecosystems (Smith et al., 2014b).
Phaeocystis globosa is supposed to be diverged from the cold-water species at approximately 30 million years ago (MYA) by molecular clock calculations (Medlin and Zingone, 2007), and now has spread in the temperate and tropical waters of the global ocean (Schoemann et al., 2005). Accordingly,the substantial diversities have been accumulated among the diff erent geographic populations of P. globosa during the long time of evolution, such as the morphological characteristics, cell and colony sizes, pigment features, and even toxic impacts on marine organisms (Vaulot et al., 1994; Qi et al., 2004;Smith et al., 2014b; Wang et al., 2019). It had once been hypothesized that P. globosa might have suffi cient time to develop detectable mutations at the nuclear ribosomal DNA (rDNA) locus to distinguish distinctly geographic populations and genetic divergence (Lange et al., 2002; Chen et al., 2003;Decelle et al., 2012). However, the fact is that although the phylogenetic relationships are close among P. globosa strains from same geographic region(Lange et al., 2002; Chen et al., 2003), there are no perspicuous phylogenetic relationships in diff erent geographic populations of P. globosa based on nuclear 18S rRNA gene and ITS region, for a part reason that a high level of rDNA genetic overlaps and polymorphisms are present in diff erent geographic strains of P. globosa (Lange et al., 2002; Chen et al.,2003; Qu et al., 2008; Decelle et al., 2012; Hu et al.,2019a). Moreover, the polymorphisms of rDNA sequences also exist in a colony, or in diff erent clones from a single cell, which is a kind of phenomenon rarely occurred in the cold-water species P. antarctica(Lange et al., 2002; Medlin and Zingone, 2007). As a result, the existing of genetic overlaps and polymorphisms in rDNA sequences have greatly hampered the understanding of the phylogenetic relationships of P. globosa populations.
Compared to the nuclear ribosomal genes including 18S rDNA, 28S rDNA, and ITS region commonly used for phylogenetic analysis (Lange et al., 2002;Chen et al., 2003; Qu et al., 2008; Decelle et al., 2012;Hu et al., 2019a), haploidic organelle genes are often uni-parental inheritance, which exclude the impacts of genetic fusion during sexual reproduction on the geographic population analysis (Hu et al., 2019a).Previously, the mutation rates of P. globosa chloroplast genome had been believed to be lower than that in mitochondria and nucleus (Smith et al.,2014a), and the highly conservative chloroplast genes psa A, psb A, and RUBISCO intergenic spacer were often used as interspecific DNA markers of the genus Phaeocystis (Lange et al., 2002; Yang et al., 2004;Yang et al., 2015). Nowadays, more and more chloroplast genomes have been assembled and compared in fast-developing next-generation sequencing technology, and it is found that noncoding and coding regions of chloroplast genomes diff er greatly in the evolution rate of many species of plants and algae (Yan et al., 2015). More hypervariable and richer variations such as substitutions,translocations, inversions, insertions, and deletions are found in non-coding region (Katayama et al.,2012), and the chloroplast intergenic spacers, for example, represent ideal segments for interspecific phylogenetic studies at low taxonomic levels and reconstruction of genetic relationships (Bakker et al.,1999; Kimura et al., 2003; Hu et al., 2011; Carbonell-Caballero et al., 2015; Kholina et al., 2016; Raman et al., 2016; Zhang et al., 2016; Li et al., 2017; Vitales et al., 2019; Zhou et al., 2019). There is however no report on the evaluation and application of chloroplast intergenic markers for intraspecific phylogeographic studies on P. globosa at present.

Fig.1 Collection sites of thirteen strains of Phaeocystis globosa
In this study, the sequence polymorphisms, genetic distances and phylogeographic relationships of nuclear 28S rDNA D1-D2 and ITS regions, and three chloroplast intergenic spacers in diff erent geographic strains of P. globosa from the Pacific and Atlantic coastal waters were compared and evaluated. The results presented a high-resolution chloroplast DNA marker to diff erentiate the geographic strains of P. globosa; therefore, much useful information was provided about the phylogenetic relationships of the Pacific and Atlantic populations.
2 MATERIAL AND METHOD
2.1 Algal culture
Seventeen strains of Phaeocystis genus were used in this study, including 13 strains of P. globosa isolated from the Pacific and Atlantic coastal waters(Fig.1), one strain of P. antarctica, one strain of P. cordata, one strain of P. rex, and one strain of P. jahnii (Table 1). They were all cultured in the laboratory at 18± 1 °C with the light intensity of 100 μE/(m2·s) and a light cycle of 14 h꞉10 h (light꞉dark), with the except of P. antarctica at 5± 1 °C.Approximately 20 mL of algal cultures were filtered on a 0.4-μm HTTP membrane (Millipore, USA) under the vacuum level of 40 kPa, and the membranes were then kept at -80 °C until DNA extraction.
2.2 Design and selection of primers
The chloroplast genomes of P. antarctica(JN117275) and P. globosa Pg-G(A) (KC900889)were downloaded from the GenBank, and aligned by Clustal W. Three chloroplast intergenic spacers ( pet Ntrn S1, trn M1- psb A and rbc S- rpl 27) with high variability were selected as the target sequences, and three primer pairs were designed accordantly by Primer 5.05: pet N-t rn S1-F (5′-GTCCCCAAACTACTAATGCT-3′), pet N- trn S1-R (5′-GGTGGCTGAGTGGTTGAAAG-3′); trn M1- psb A-F (5′-AGCAGTTTGGTAGCTCGTCG-3′), trn M1- psb A-R (5′-TGTTGATTCTCAAGGTCGTGTA-3′); rbc Srpl 27-F (5′-GAAGCATACGCACCAAGC-3′), rbc Srpl 27-R (5′-CAGCCAGTAAAGGCAGGT-3′). The primer pair for 28S rDNA D1-D2 (forward primer LSU-F2: 5′-ASAGYCGCCTCCTGAATTGTAGTC-3′,reverse primer LSU-R3: 5′-TCGAGCTTGCCACTCTAGTACTC-3′) (Hu et al., 2019a), and the primer pair for ITS region (forward primer Euk P18S(1705-)-F: 5′-GCCGGACGCGACGCTCC-3′, and reverse primer ITS-2R: 5′-GCTTATTGATATGCTTAAGTTCAGCGGGT-3′) (Blomster et al., 1998; Lange et al., 2002) were applied as references.

Table 1 Information of microalgae in this study
2.3 Genomic DNA extraction, PCR amplification,and sequencing
Total genomic DNA of microalgal samples was extracted with the modified method described by Winnepenninckx et al. (1993), and stored at -80 °C until analysis. PCRs were executed with a final volume of 20 μL containing 8.2-μL ddH2O, 4 μL 5×rTaq Buff er (Mg2+), 4 μL 5×PCR solution buff er, 1.6-μL dNTP Mix (2.5 mmol/L), 1 μL of DNA template(10 ng DNA template in each reaction), 0.4 μL of each PCR primer (10 pmol/mL), and 0.4-μL super-fidelity Taq DNA polymerase (5 U/mL, TransGen Biotech Co., Ltd., China). The amplifications were performed with an initial denaturation temperature at 98 °C for 2 min, 32 cycles of 98 °C for 20 s, 58 °C for 20 s, and 72 °C for 30 s, and a final elongation at 72 °C for 5 min. The PCR products were confirmed by electrophoresis in 1% agar gel, and the targeted DNA bands were purified and then sequenced from both ends by Sangon Biotech (Shanghai). The obtained forward and reversed fragments were assembled by Invitrogen 11.0 ContigExpress.
2.4 Clone library construction
Because the PCR productions ofiTS regions from 10 strains of P. globosa could not be assembled due to the nucleotide differences, the clone libraries were constructed respectively by using the purified PCR fragments to obtain their full sequences, and to show the polymorphisms in the target genes. The fragments were cloned into blunt simple cloning vectors(TransGen Biotech Co., Ltd., China), and the vectors were transformed into Trans 1-T1 competent Escherichia coli cells (TransGen Biotech Co., Ltd.,China), which were placed on LB ampicillin plates containing final concentration of 100 μg/mL ampicillin. Three colonies were random selected from each plate and sequenced from both ends.
In addition, the clone libraries of 28S rDNA D1-D2 region, ITS region and rbc S- rpl 27 intergenic spacer of P. globosa MEL43 and MEL47 were constructed as the above-mentioned, except that 30-50 colonies from each library were selected and sequenced from both ends.
2.5 Genetic distances and phylogenetic analysis
The sequences of 28S rDNA region and ITS region of P. globosa, P. antarctica, P. cordata were downloaded from the GenBank (Suppl. Table S1).Together with the sequences of 28S rDNA D1-D2 region, ITS region and pet N- trn S1, trn M1- psb A,and rbc S- rpl 27 intergenic spacers obtained in this study, the genetic distances of the diff erent target sequences were estimated by Mega 10 pairwise distance method with bootstrap replications of 1 000(Kumar et al., 2018). The maximum-likelihood trees of these five genes and intergenic spacers were built by Mega 10 variance estimation method with the bootstrap replications of 1 000 and the best-fitting nucleotide substitution models of T92, GTR+G,T92+G, T92 and K2+G were selected, respectively(Kumar et al., 2018). The Bayesian analysis of these five genes and intergenic spacers was constructed by MrBayes 3.2.6 (Ronquist et al., 2012) with generations of 5 000 000 and sampling frequencies of 100, and the best-fitting nucleotide substitution models of HKY+F+I, GTR+F+G4, HKY+F+I,HKY+F+G4, and K2P+G4, were selected using Modelfinder (Kalyaanamoorthy et al., 2017). The sequences of P. antarctica and P. cordata were as the outside group. Additionally, the sequences of 28S rDNA D1-D2 region, ITS region and rbc Srpl 27 intergenic spacer resulted from the clone libraries of P. globosa MEL43 and MEL47 were aligned by Clustal W. The multiple aligned sequences were used to analyze the relative frequency of the nucleic acid at each position by the online software WebLogo 3 (http://weblogo.berkeley.edu/logo.cgi)(Crooks et al., 2004).
3 RESULT
3.1 Primers design and their performances
The chloroplast genomes of P. globosa, and P. antarctica were downloaded and aligned, and three chloroplast intergenic spacers with a relatively high nucleotide variation level were identified and acted as the aimed regions, which were pe tN- trn S1,trn M1- psb A, and rbc S- rpl 27 intergenic spacers.Three primer pairs were designed and used for the amplification of the targeted chloroplast intergenic spacers from all the 15 Phaeocystis strains in this study in a diff erent sequence length, and the amplified sequences were submitted to GenBank(ID MN807922-MN807935, MN928971-MN928984, MN935489-MN935501, and MN956829, the detailed information is shown in Suppl. Table S2). In detail, the pet N-t rn S1 spacers of P. antarctica CCMP1374 and P. cordata CCMP3104 were 368 bp and 341 bp, respectively,while for the strains of P. globosa, the pet N-t rn S1 spacer of RCC736 was 347 bp and the other 12 strains were 528 bp. The trn M1- psb A spacers of P. antarctica CCMP1374 and P. cordata CCMP3104 were 549 bp and 418 bp, respectively, and that of P. globosa were from 648 bp to 682 bp. The rbc Srpl 27 spacers of P. antarctica CCMP1374 and P. cordata CCMP3104 were 1 115 bp and 709 bp,respectively. Within the species of P. globosa, the rbc S- rpl 27 spacer of RCC736 was 599 bp and the other 12 strains were 609 bp.
3.2 Genetic distances and sequence polymorphism of nuclear 28S rDNA D1–D2 region of P. globosa
The sequences of 28S rDNA D1-D2 region with the length of 522 bp were amplified by the primer pair LSU-F2 and LSU-R3 from the 13 isolated strains of P. globosa and submitted to GenBank (ID MN602538-MN602550, the detailed information is shown in Suppl. Table S2). Combined with other 17 P. globosa 28S rDNA D1-D2 region sequences downloaded from the GenBank (Suppl. Table S1), the genetic distances of 28S rDNA D1-D2 region from diff erent strains of P. globosa were analyzed. As shown in Table 2, the obtained sequences could be categorized into nine groups with one to eleven same members in each group, and the values of genetic distances between these groups ranged from 0.000 0 to 0.013 7,while the grouping of these sequences failed to be directly related to the geographic regions.
To examine the potential polymorphisms that might be neglected by the directly assemble of the forward and reverse PCR fragments, the representative strains of MEL43 and MEL47 were chosen for the clone library construction, and 34 sequences(GenBank ID MN602900-MN602933, the detailed information is shown in Suppl. Table S3) and 40 sequences (GenBank ID MN610012-MN610051,the detailed information is shown in Suppl. Table S3)of 28S rDNA D1-D2 region were obtained. The alignment result showed that there were seven and four SNP sites existed in MEL43 and MEL47 clones,respectively (Suppl. Figs.S1 & S2), and 7 SNP sites between the two strains (Fig.2), indicating a relatively low level of the gene variations in this region.
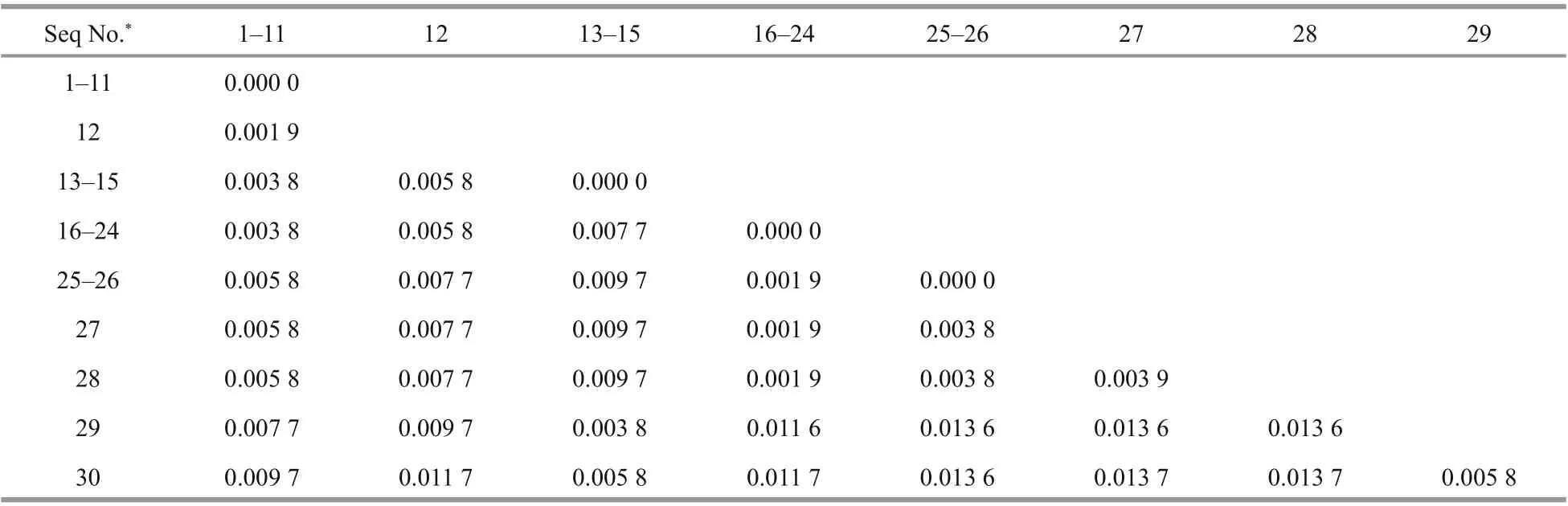
Table 2 Genetic distances between 28S rDNA D1–D2 region sequences of Phaeocystis globosa

Table 3 Genetic distances ofiTS region sequences from the clones of ten strains of P. globosa
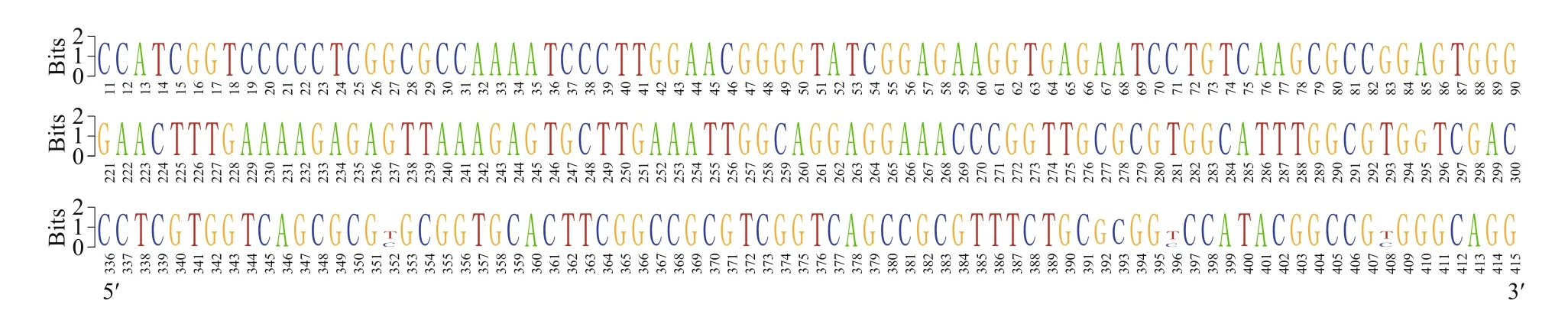
Fig.2 WebLogo of 28S rDNA D1–D2 region between two strains of Phaeocystis globosa MEL43 and MEL47
3.3 Genetic distances and sequence polymorphism of nuclear ribosomal ITS region of P. globosa
The sequences of nuclear ribosomal ITS region were successfully amplified by the primer pair Euk P18S and ITS-2R from all thirteen strains of P. globosa, but for only three strains of MEL71,PGDH2019C and RCC736, the forward and reverse PCR fragments could be directly assembled with the length of 871-872 bp. For the other ten strains, ITS clone libraries were constructed, and then three clones from each library were randomly sequenced from both ends to get the full gene length of 895-925 bp.Accordantly with the direct assemble result of PCR productions, ITS sequence variations from the clone libraries were observed in every strain of P. globosa with the divergent rate of 0.001 4-0.040 7 (Table 3),indicating a high polymorphism level in this region.The genetic distances were analyzed for the obtained ITS sequences (GenBank ID MN603007-MN603009,MN603057-MN603086, the detailed information in Suppl. Tables S2 & S4) and the other nineteen sequences of P. globosa downloaded from GenBank(Suppl. Table S1). Their values of genetic distances ranging 0.000 0-0.067 4 (Suppl. Table S5), were much higher than that of 28S rDNA D1-D2 region.
To further evaluate ITS polymorphism level in diff erent strains of P. globosa, the clone libraries sampled from strain MEL43 and MEL47 were constructed, and 28 sequences (ID MN603081-MN603083, MN603166-MN603190, the detailed information is shown in Suppl. Table S4) and 35 sequences (ID MN603084-MN603086, MN603191-MN603222, the detailed information in Suppl.Table S5) were sequenced and assembled respectively.The webLogo analysis revealed 160 and 119 sites nucleotide variation, insertion and deletion in the MEL43 and MEL47 clone libraries (Suppl. Figs.S3 &S4). In addition, there were 177 nucleotide sites varying between the two strains, which accounted for 19.13% sites of the full sequences (Fig.3). The results displayed a rather high level of mutation rate in the ITS region of P. globosa.

Table 4 Genetic distances between sequences of pet N- trn S1 intergenic spacer of fourteen strains of Phaeocystis globosa
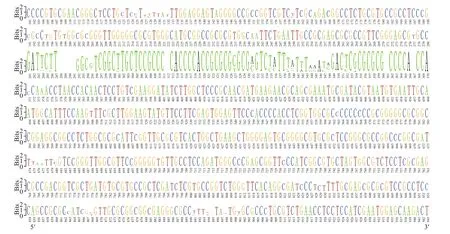
Fig.3 WebLogo ofiTS region between two strains of P. globosa MEL43 and MEL47
3.4 Genetic distances and sequence polymorphism of the chloroplast intergenic spacers of P. globosa
The genetic distances of three chloroplast intergenic spacers of P. globosa were analyzed and compared.As shown in Table 4, the pet N- trn S1 spacer sequences had no difference in the nine strains from SCS, ECS and Gulf of Thailand Pacific, and Caribbean Sea, Gulf of Mexico and English Channel Atlantic, while theother two strains from ECS Pacific (MEL71 and PGDHC) were diff erent and showed a moderate variation compared to other ECS Pacific strains, but same with the one from the North America Atlantic(CCMP2754). The highest divergence was observed in two strains from the North Sea Atlantic (RCC736 and Pg-G(A)), indicating their early separation from other strains.

Table 6 Genetic distances between sequences of rbc S- rpl 27 intergenic spacer of 14 strains of Phaeocystis globosa
As shown in Table 5, there was no nucleotide difference in the trn M1- psb A spacer sequences of six strains from SCS, ECS, and Gulf of Thailand in the Pacific Ocean, and these strains had genetic distance of 0.011 6 with three strains from Caribbean Sea, Gulf of Mexico, and English Channel Atlantic (CCMP628,CCMP629 and RCC2055), respectively. The CCMP2754 strain and the other two strains from ECS Pacific (MEL71 and PGDHC) had further genetic distance with the value of 0.011 6-0.021 5. And the highest divergence (0.315 2-0.320 1) was observed in two strains from the North Sea Atlantic (RCC736 and Pg-G(A)).
As shown in Table 6, the relationships of genetic distances for rbc S- rpl 27 intergenic spacer sequences were similar with that for trn M1- psb A. But it was worth noting that the four SCS strains generated genetic divergence in rbc S- rpl 27 when compared to other strains isolated ECS and Gulf of Thailand,which exemplified that rbc S- rpl 27 provided better performance to distinguish the diff erent geographic strains of P. globosa than the other two markers of pet N- trn S1 and trn M1- psb A. The polymorphisms of this region were then further testified by clone sequencing sampled from MEL43 and MEL47,respectively. The results showed that there was no nucleotide variation of rbc S- rpl 27 in 40 sequences from MEL43 (GenBank ID MN990301-MN990340,the detailed information in Suppl. Table S6) and 32 sequences from MEL47 (GenBank ID MN990269-MN990300, the detailed information in Suppl. Table S6).
3.5 Phylogenetic relationship of diff erent geographic strains of P. globosa
Thirty sequences of 28S rDNA D1-D2 region of P. globosa strains from the Pacific Ocean and Atlantic Ocean, were used to build the phylogenetic tree, with the sequences from P. antarctica and P. cordata as the outgroup branch. These strains of P. globosa were mainly divided into two clades (Fig.4): the sequences of 11 strains of P. globosa isolated from the Pacific Ocean and the Atlantic Ocean formed one clade with the other six strains sampled from China Sea, the Pacific Ocean, and the North Atlantic Ocean. In the other clade, 9 strains ( Phaeocystis sp. Ros1,Phaeocystis sp. JD-2012 Ros2, Phaeocystis sp.RCC679, Phaeocystis sp. AC618, P. globosa CCMP2754, P. globosa RCC64, P. globosa RCC678,P. globosa K1321, and P. globosa NIES388) had no nucleotide difference, and then gathered with PGDHC, MEL71, RCC736, AC618 and three endosymbiotic strains ( Phaeocystis sp. Ind12,Phaeocystis sp. Ros1, and Phaeocystis sp. JD-2012 Ros2). It was obvious that the phylogenetic tree of 28S rDNA failed to diff erentiate the P. globosa strains from distinct regions.
Thirty-three ITS region sequences of P. globosa obtained in this study and the other nineteen P. globosa ITS region sequences downloaded from the GenBank were used to construct the phylogenetic tree. The phylogenetic relationship of diff erent geographic P. globosa strains was more complex than that in the 28S rDNA tree. The MEL71 and PGDHC isolated from the Sansha Bay of ECS stayed together,and then gathered with two sequences isolated from a P. globosa bloom in the Bohai Sea in the year of 2005(Qu et al., 2008), staying at the base position of P. globosa clade. They were close to the three sequences of CCMP2754 clones, and were followed by RCC736 and seven sequences of P. globosa isolated from the English Channel. The clone sequences of nine strains of P. globosa (MEL43,MEL44, MEL69, MEL45, MEL47, CCMP628,CCMP629, CCMP1524, and RCC2055) stayed together, and formed a big branch with the sequences of the diff erent clones of CCMP627 isolated from the Gulf of Mexico in the Atlantic Ocean, as well as Santou 97 and JD-2012 collected from SCS (Fig.5).As a result, the phylogenetic tree ofiTS region interlaced, and the phylogenetic relationships of P. globosa were inconsistent with their geographic distribution (Fig.5).
The phylogenetic trees of pet N- trn S1, trn M1-psb A, and rbc S- rpl 27 intergenic spacers were similar (Figs.6-8), all of P. globosa strains stayed together, closing with P. antarctica, and far away from P. cordata. However, the branch of P. rex and P. jahnii had greater genetic distances with P. globosa than P. antarctica and P. cordata in the tree of rbc S- rpl 27. In the three trees, all of P. globosa were divided into two independent clades,respectively. Collected from the Netherlands coastal waters of the Atlantic Ocean (Smith et al., 2014a),the two strains of P. globosa (RCC736 and Pg-G(A))early separated from other strains and formed a clade staying at the base position. The rbc S- rpl 27 tree had the highest resolution in the other clade, for that it could distinguish the four SCS strains (MEL43,MEL44, MEL69 and MEL45) from the two strains collected from the adjacent coastal waters of ECS and the Gulf of Thailand (MEL47 and CCMP1524),however all of them still stayed in a same branch in both pet N- trn S1 and trn M1- psb A trees. Moreover, a common feature of this clade was that the Pacific and Atlantic strains were generally clustered separately, except that MEL71 and PGDHC collected from ECS gathered with the Atlantic strain CCMP2754 with a bit difference.
4 DISCUSSION
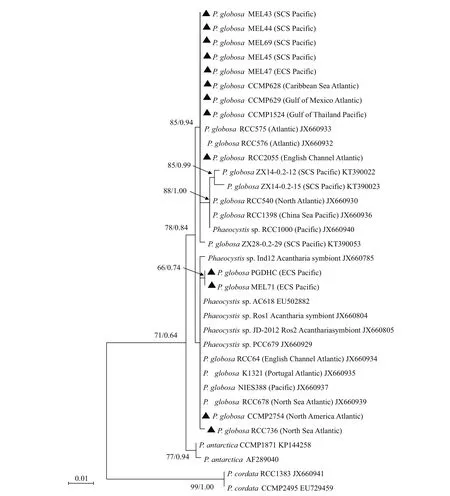
Fig.4 Phylogenetic tree of 28S rDNA D1–D2 region of Phaeocystis globosa
Diverging from cold-water species at 30 MYA,P. globosa distributes in both temperate and tropical oceans worldwide (Schoemann et al., 2005; Medlin and Zingone, 2007). Diverse geographic distribution patterns had urged the existence of evolutionary divergence, and genetic diff erentiation had taken place within the P. globosa taxon (Lange et al., 2002; Chen et al., 2003; Hu et al., 2019a). The convincing genetic divergence among diff erent geographic strains is confirmed by a lot of studies on the basis of sequences of the nuclear 18S rDNA (Lange et al., 2002; Qu et al.,2008), 28S rDNA (Hu et al., 2019a), ITS (Lange et al.,2002; Chen et al., 2003; Qu et al., 2008; Liu et al.,2010; Hu et al., 2019a), and the concatenation of 18S rDNA, 28S rDNA, psb A and rbc L genes (Decelle et al., 2012). Morphological characteristics and pigment features also provide reliable evidences (Vaulot et al.,1994; Qi et al., 2004; Hu et al., 2019b; Wang et al.,2019), such as the giant colonies of Southeast Asian strains (Qi et al., 2004; Smith et al., 2014b), and the change of concentration of characteristic pigments 19’-hexanoyloxyfucoxanthin (Hex-fuco) and 19’-butanoyloxyfucoxanthin (But-fuco) of diff erent geographic strains (Vaulot et al., 1994; Antajan et al.,2004; Zapata et al., 2004; Seoane et al., 2009; Wang et al., 2019). In brief, the diff erentiation of morphological features, biochemical characteristics, and molecular information indeed reflect the existence of the multiple populations within the P. globosa taxon, and their diversities among the diff erent geographic populations.
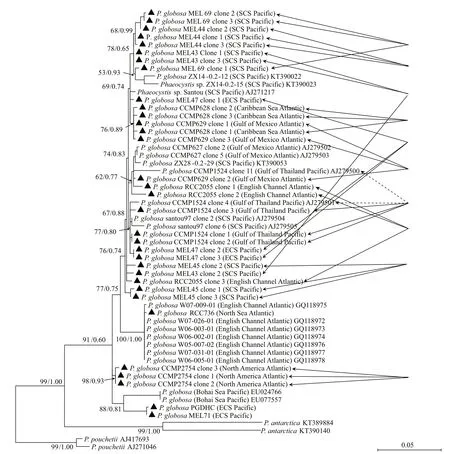
Fig.5 Phylogenetic tree ofiTS region of Phaeocystis globosa
4.1 Evaluation of the nuclear ribosomal and chloroplast DNA markers in phylogenetic analysis of geographic populations of P. globosa
At present, the phylogenetic relationship of P. globosa populations distributing in these diverse habitats has not been well understood. In general, the strains within a geographic population are easy to acquire similar evolutionary rate, and have small genetic distances under the same environmental pressure. However, there is no obvious correlation between phylogenetic relationship and geographic distribution among P. globosa strains based on the molecular data of nuclear ribosomal genes and intergenic spacers (Lange et al., 2002; Qu et al., 2008;Liu et al., 2010; Hu et al., 2019a). For example, it was found that a strain of P. globosa from the Bohai Sea is closely related to the strain isolated from the Atlantic Ocean, but far away from the strains from SCS base on the 18S rDNA sequences (Qu et al., 2008). Furthermore,our study found that there are nucleotide variations in 28S rDNA D1-D2 region and ITS region within a single strain cultured from either a colony or a single cell (Suppl. Figs.S1-S4), and similar results are also supported by the previous studies (Lange et al., 2002;Chen et al., 2003; Hu et al., 2019a). The reasons for the non-homogeneous copies of the nuclear ribosomal genes in P. globosa are somehow enigmatic.
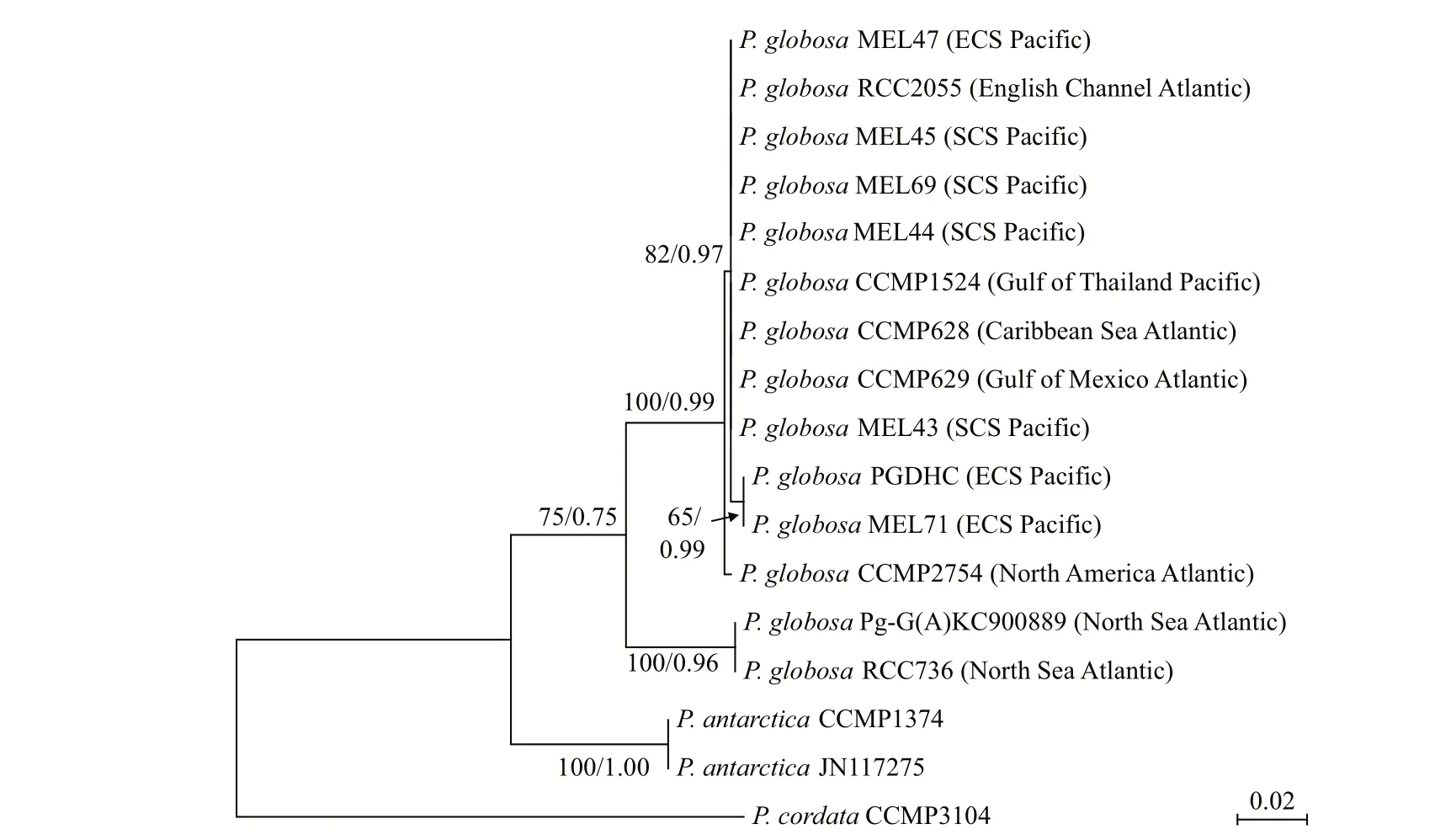
Fig.6 Phylogenetic tree of pet N- trn S1 intergenic spacer of P. globosa

Fig.7 Phylogenetic tree of trn M1- psb A intergenic spacer of P. globosa
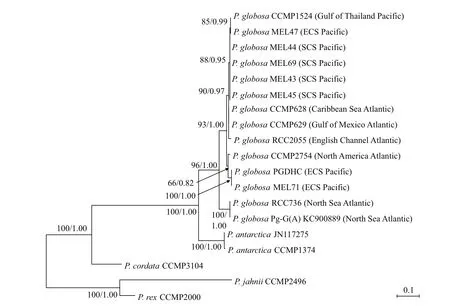
Fig.8 Phylogenetic tree of rbc S- rpl 27 intergenic spacer of P. globosa
A possible explanation is that P. globosa is indeed a multi-species complex as indicated by the variation in DNA content among several strains (Lange et al.,2002). The other explanation is that P. globosa has a complex polymorphic life cycle with both flagellated and colonial cells. A haploid-diploid life cycle with both ploidies alternately existing or/and co-existing has been evidenced for P. globosa (Whipple et al.,2005b; Rousseau et al., 2007; Peperzak and Gäbler-Schwarz, 2012). Two main prominent features of this cycle are that sexuality is prevalent in colony bloom formation and termination and two types of vegetative reproduction may exist at same time (Rousseau et al.,2007). Frequently sexual processes tend to cause genetic fusion between diff erent populations. For example, a pervious study found that four haplotypes were contained in the 28S rDNA genes of two P.globosa strains isolated from the Beibu Gulfin SCS in 2015 and 2017, that covered a unique haplotype of another P. globosa strain isolated from the same region in 2016 (Hu et al., 2019a). Additionally, the habitats in the temperate and tropical sea areas are widely aff ected by human shipping activities, which accelerate gene flow between diff erent populations contributing to form rDNA genetic polymorphism characteristics of P. globosa (Hu et al., 2019a). In any event, the non-homogeneous copies of the nuclear ribosomal genes are impossible to make interpretation of phylogenetic relationship within the P. globosa taxon.
Chloroplast is an important organelle involved in photosynthesis, supplying the indispensable energy for photosynthetic organism growth and development.The haploid chloroplast genome shows maternal inheritance in the genus Phaeocystis, which has a red algae-derived secondary plastid (Smith et al., 2014a).The chloroplast DNA has less recombination, and is substantially diff erent from the nuclear genome(Wolfe et al., 1987). As displayed in this study, the phenomenon of non-homogeneous copies of genes is not present in the chloroplast intergenic spacers,which makes them a potential molecular marker for phylogenetic analysis among P. globosa geographic populations. Although it is confirmed that some conservative chloroplast genes and intergenic spacers are not suitable to act as intraspecific DNA markers of P. globosa (Lange et al., 2002; Yang et al., 2004;Yang et al., 2015), all of three chloroplast intergenic spacers selected in this study have high genetic resolution across diff erent geographic strains. In sum,the chloroplast intergenic spacers are better molecular markers for intraspecific phylogeographic relationships on P. globosa than the nuclear ribosomal genes and intergenic spacers.
Among the three chloroplastal intergenic spacers,the rbc S- rpl 27 intergenic spacer is a reasonable indicator of the genetic distance between Pacific and Atlantic strains, especially for the genetic difference between the Pacific strains, which can not be shown in the other two chloroplastal intergenic spacers.Moreover, the resolution ability of the rbc S- rpl 27 intergenic spacer to P. globosa strains is similar with that of the concatenation of three chloroplast genetic spacers (Suppl. Fig.S5). The results imply that the genetic variation of rbc S- rpl 27 intergenic spacer is the highest, and by far, it is most suitable to be served as the DNA marker for the intraspecific phylogeographic relationships of P. globosa.
4.2 Phylogeographic relationships of P. globosa in the Pacific and Atlantic coastal areas
In the genus Phaeocystis, the genetic relationships of three species P. antarctica, P. globosa and P. pouchetii are close, and they are speculated to be evolved from a common ancestor. When the Drake Passage opened and the Antarctic Circumpolar Current (ACC) formed, suffi ciently allowed ancestral populations in the Antarctic to separate from their warm-water ancestors (Lange et al., 2002; Medlin and Zingone, 2007). While a short but deep glacial period happened around 23 MYA (Paul et al., 2000),water temperature was cool enough to allow the ancestral Antarctic populations cross the equator from the south to the north (Darling et al., 2000, 2004;Lange et al., 2002; Medlin and Zingone, 2007). Then,a major warming event in the world’s oceans at approximately 15 MYA separated the two polar populations to allow them to diverge into two species( P. antarctica and P. pouchetii), and the relict populations in the temperate and tropical regions evolved into today's P. globosa (Lange et al., 2002;Medlin and Zingone, 2007).
In this study, the legible phylogeographic relationships of P. globosa from the Pacific and Atlantic coastal waters are shown in the tree of rbc Srpl 27 intergenic spacer. Two strains (RCC736 and Pg-G(A)) from the North Sea in the Atlantic Ocean form an independent clade, and detach from the P. globosa taxon firstly; however, other Atlantic strains stay with the Pacific strains. The population has unique pigment characteristics in the North Sea. Hex-fuco content is lower than But-fuco content in cell of RCC736 culture, which is diff erent from other 12 strains of P. globosa in this study (our unpublished data).Moreover, the pervious study reported that the pigment composition and DNA content of the North Sea strains were similar, but diff erent with the other geographic strains (Vaulot et al., 1994). These data support evidences for the uniqueness of the geographic population in the North Sea. As a result, it is deduced that there are at least two genetically distant populations of P. globosa in the Atlantic coastal regions, and these populations may have diff erent geographic ancestries.
In the other clade, there is no nucleotide difference in the rbc S- rpl 27 intergenic spacer among four SCS strains. Among them, MEL43, MEL44, and MEL69 were isolated from the Beibu Gulfin the western waters of SCS in the years of 2017 and 2018,respectively, and MEL45 was isolated from the Zhujiang (Pearl) River estuary in the eastern waters of SCS. The eastern and western waters of SCS are connected thought narrow Qiongzhou Strait. There is a strong year-round westward flow in the Qiongzhou Strait, which is a source of water transport to the Beibu Gulf existing cyclonic circulation (Shi et al.,2002). This kind of hydrological characteristics is well supported that four SCS strains are belong to a same geographic population. The other two Pacific strains (MEL47 and CCMP1524) have closely phylogenetic relationship with the SCS population.They were respectively isolated from the Quanzhou coastal area in ECS and the Gulf of Thailand, adjacent to SCS, which help to explain reasonably the close genetic relationship with the SCS population. It seems unreasonable that the two ECS strains (MEL71 and PGDHC) keep far away the other Pacific strains, and gather with the CCMP2754 strain from the North American coastal area. It is pluckily suspected these ECS strains may be invasive species from the Northeastern Atlantic region.
5 CONCLUSION
In this study, it was confirmed that the region of chloroplast intergenic spacer was a good target to be used for the intraspecific phylogeographic study of P. globosa, and a clear phylogenetic relationship of P. globosa strains was firstly revealed by the highresolution molecular marker, the chloroplast rbc Srpl 27 intergenic spacer. The results displayed that strains from the North Sea Atlantic separated from the species P. globosa firstly, and kept far away from the other Atlantic strains and all of Pacific strains, which implied that there were at least two genetically distant populations of P. globosa in the Atlantic coastal regions.
6 DATA AVAILABILITY STATEMENT
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
7 ACKNOWLEDGMENT
We deeply appreciate valuable comments of Dr.QIU Limei from the Institute of Oceanology, Chinese Academy of Sciences (IOCAS), and free provision of two strains of Phaeocystis globosa RCC736 and RCC2055 by Prof. Daniel Vaulot and Ian Probert from the Roscoff Biological Station.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Predicting sediment flux from continental shelfislands,southeastern China*
- Laboratory simulation of dissolved oxygen reduction and ammonia nitrogen generation in the decay stage of harmful algae bloom*
- Effects ofiron and humic acid on competition between Microcystis aeruginosa and Scenedesmus obliquus revealed by HPLC analysis of pigments*
- Effect of river plume on phytoplankton community structure in Zhujiang River estuary*
- Exploring the sublethal genotoxic effects of class II organophosphorus insecticide quinalphos on freshwater fish Cyprinus carpio
- An illustrated checklist of macrofaunal molluscs from softsediments of the Abrolhos Parcel (northeastern, Brazil) with 15 new records*