Seasonal and spatial variations of macrobenthic community in intertidal Sargassum field off Naozhou Island, South China Sea*
2021-04-14CaixueZHANGWeinanZHOUXingliSUNShengKEYaoqianLIUZhiguangSONG
Caixue ZHANG, Weinan ZHOU, Xingli SUN, Sheng KE, Yaoqian LIU, Zhiguang SONG
Monitoring Center of Ocean Resource and Environment, Guangdong Ocean University, Zhanjiang 524088, China
Abstract Seasonal and spatial variations in the macrobenthic community in an intertidal gulfweed field off Naozhou Island were investigated from April 2011 to January 2012. We identified 407 macrobenthic organism species: 64 macroalgae (15.72% of total species); 123 Arthropoda (30.22%); 107 Mollusca(26.29%); 67 Annelida (16.46%); 15 Cnidaria (3.69%); 14 Echinodermata (3.44%); 8 Porifera (1.97%); 4 Chordata (0.98%); 2 Urochorda (0.49%); and 1 each of Echiura, Sipunculida and Angiospermae (0.25%).Arthropods, mollusks, annelids, and macroalgae were four main groups in the benthic organism community and accounted for 88.70% of the total species. In terms of abundance of biomass, there were 15 dominant species, including 8 macroalgal, 5 mollusk and 2 arthropod species. The species diversity indices ranged from 1.21 to 4.42 with an average of 2.80; the annual average of evenness was 0.47. Species richness indices varied from 2.97 to 7.24 (average 4.97); and Simpson dominance indices varied from 0.20 to 0.92 in annual average of 0.75. Macroalgal distribution showed clear vertical zonation and seasonal changes. The number of macroalgal species found in the high tide zone, upper middle tide zone, middle tide zone, lower middle tide zone, low tide zone, and below low tide zone were 1, 8, 10, 16, 22, and 32, respectively. Biomass among the benthic organisms, macroalgae and crustaceans had significant positive correlation ( P <0.05)with the coeffi cients of 0.802, 0.894, and 0.592, respectively. Macroalgal biomass showed a clearly negative correlation with the concentration of dissolved inorganic nitrogen (DIN) ( R=0.486, P <0.05). During the winter and spring, when macroalgae were growing vigorously, DIN reached the lowest level in seawater and displayed a clear weak correlation between macroalgal growth and the nutrient concentration.
Keyword: macrobenthic marine algae; species diversity; seasonal distribution; spatial distribution; Naozhou Island
1 INTRODUCTION
Macroalgal beds are widely distributed in nearshore regions of the world. They are unique marine ecosystems primarily composed of various communities of benthic organisms (Zhang and Sun,2007). These seafloor ecosystems play important ecological roles and represent the most important primary production along coastal areas. For example,macroalgae and epiphytes provide food and spawning grounds for a variety of marine life, including spawning fish (Japanese Society of Fisheries Science,2000). The photosynthetic carbon sequestration of seafloor algae plays a crucial role in the ocean carbon cycle, as the main carrier of blue carbon sinks.Macroalgae are also an important pool of nitrogen (N)and phosphorus (P) in marine ecosystems. As macroalgae are able to buff er nutrients, their largescale cultivation may be able to balance the additional nutrient load generated by commercial breeding of marine animals. Their ability to remove N and P from seawater can reduce the risk of coastal eutrophication(Yang et al., 2005). Previous studies have shown that the nitrogen uptake capacity of a 1-km2Sargassum field is almost comparable to a sewage plant treating the wastewater produced by 50 000 people(Friedlander and Dawes, 1985; Brinkhuis et al.,1989). Macroalgae are one the major marine algal groups recommended globally for marine ecological restoration. Macroalgal fields provide highly heterogeneous and diverse living spaces, including shade, shelter, and narrow crevices for many marine animals and plants. They reduce the strength of waves, change the hydrodynamics of seafloor currents, buff er changes in the pH value of seawater,and dissolved oxygen content and water temperature.Therefore, they create a static, stable, submarine environment where various marine animals can thrive, and are in themselves ecological systems rich in biological resources such as poriferans,coelenterates, crustaceans, molluscs, echinoderms,and fish (Zhang and Sun, 2007). Naozhou Island was formed by a submarine volcanic eruption 200 000-500 000 years ago and its bedrock is composed of biotite granites. The island is about 40 km southeast of Zhanjiang City, Guangdong, China, and has a total land area of 56 km2and about 43.98 km of coastline.The eastern coastline experiences erosion from pounding waves. Before the 1980s, a wide variety of macroalgae—especially Sargassum naozhouense,S. hemiphyllum, S. fusiforme, and Ulva—used to grow in the low tide zone of the seafloor off eastern Naozhou Island. During the lower spring tides it was possible to observe a spectacular Sargassum field and various species of marine life through the clear seawater as the Sargassum swayed in the waves (Luo,2017). It has been reported that the annual production of Sargassum was more than 2 200 t in the Naozhou Island intertidal zone (Chen et al., 1995). Industry,agriculture and aquaculture have developed quickly for almost three decades, leading to increasing pollution in coastal waters. As a result, the Sargassum growing area is shrinking rapidly, and its specific native species— S. naozhouense, S. integerrimum and S. hemiphyllum—and the famous Naozhou abalone have almost vanished from the natural marine environment. Even the survival rate of farmed Naozhou abalone has fallen substantially because of the poor water quality. The production of nekton has also decreased dramatically, especially that of Siganidae, which feeds on algae. An earlier survey indicated that the quality of fishery resources and structure has declined rapidly in Naozhou Island, and that the percentage of traditional high-value fish species is becoming exhausted: there has been a
dramatic drop in the number of bluish fish species that feed on algae, as well as in the number oflobsters,crabs and shrimp (Zhang et al., 2008). A systematic investigation and assessment of the ecological status of algae, the marine environment and the causes of algal degeneration are urgently needed to allow restoration and reconstruction. It has been demonstrated in Japan and the United States that once-degraded kelp beds can be restored and regain their luxuriance after a great variety of restoration measures have been implemented, and, most importantly, that biodiversity has been restored and fishery resources dramatically increased (Toshinobu,2003). We sought to learn from successful ecological restoration examples in foreign countries in our investigation of the macrobenthic community structure and environmental condition of the sea beds surrounding Naozhou Island. Our study aimed to analyze the causes of algal degradation and to explore possible approaches for the restoration of seaweed beds, and therefore to provide a scientific understanding of coastal ecological remediation and protection, as well as of the conservation and sustainable use of marine biological resources in this region.
2 MATERIAL AND METHOD
2.1 Sample collection and analysis
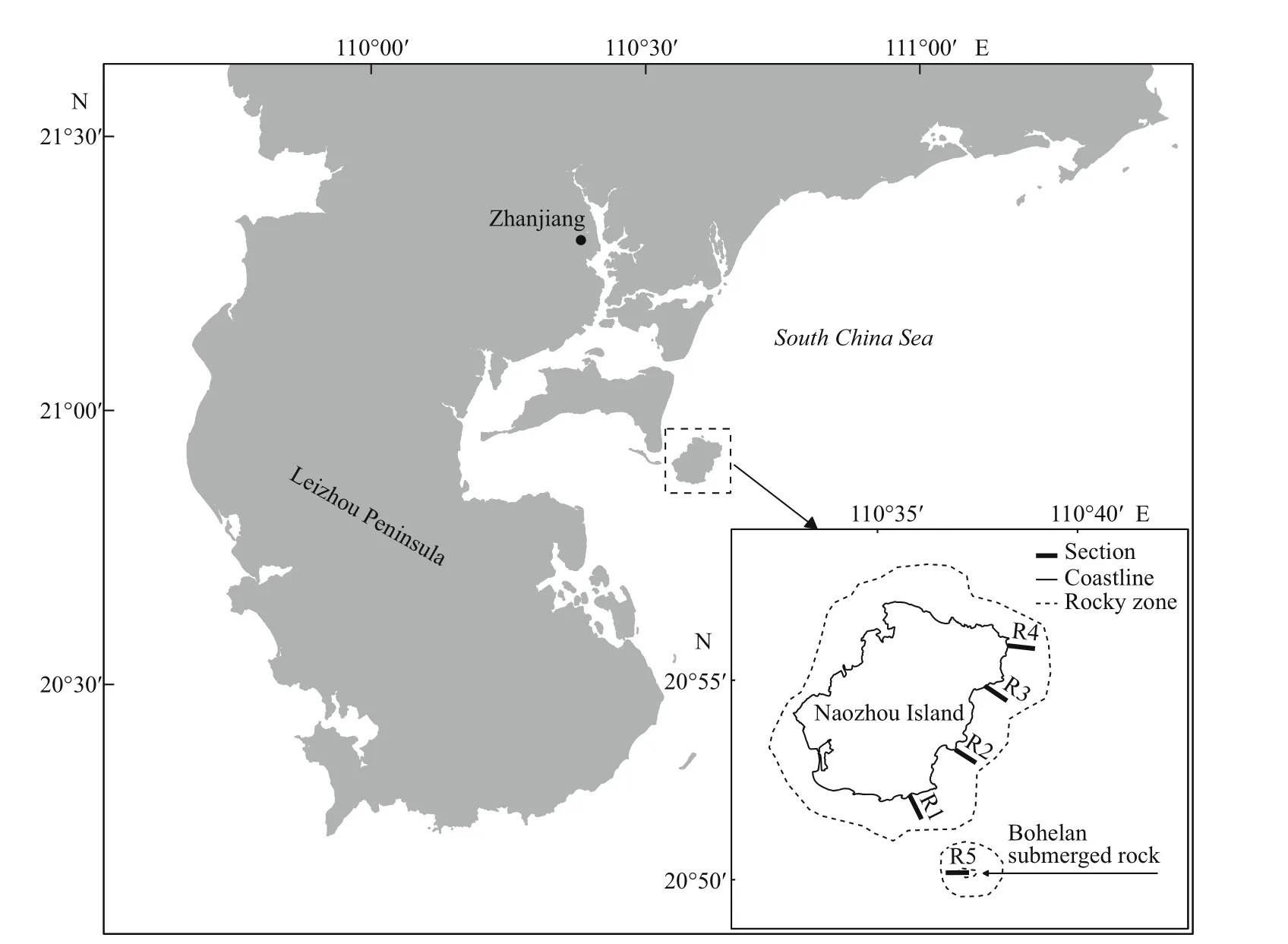
Fig.1 Sampling sections in the reef area off Naozhou Island, China
Naozhou Island is to the southeast of Zhanjiang City, Guangdong, China. It is about 40 km from the coast of the mainland and has a total land area of 56 km2and about 43.98 km of coastline. Five sampling sections (Fig.1) were assigned over the reef areas offNaozhou Island and the Bohelan reefs. Each section included six sampling stations: one in the high tidal zone, three in the mid-tidal zone (upper, middle and lower), and two in the low tidal zone (upper and lower). Sampling was carried out during high tides.The sampling sections and stations were chosen to represent the characteristics of native biological distribution. The longitude and latitude of the sampling station of each section (named R1-R5) were as follows: R1 was 20°52′14.59″N, 110°36′25.86″E;R2 was 20°53′26.5″N, 110°36′37.66″E; R3 was 20°55′17.1″N, 110°38′03.9″E; R4 was 20°56′0.04″N,110°38′16.36″E; R5 was 20°50′10.51″N,110°37′08.71″E (Fig.1). The rest of sampling stations in each section was arranged in a straight line in the off shore direction. Benthos sampling was carried out consecutively in four seasons: spring (April), summer(July), autumn (October), and winter (January) from April 2011 to January 2012. For quantitative analysis and to reduce sampling errors, two parallel 25 cm×25 cm quadrate seafloor areas were sampled at each sampling site, and all organisms within the quadrate areas were carefully counted and collected. When sampling, the fragile organisms were picked up first,then all aufwuchs, periphyton and troglobites were collected using scrapers and chisels. Qualitative sampling was performed at diff erent tidal ranges in each sampling section, and it was important to collect all living species within the sampling sections as far as possible. Zoobenthos was put into 500-mL widemouth plastic bottles, while macroalgae were packed into ziplock bags. All organism samples were soaked in a 5% formalin solution. Species classification and identification were carried out by first determining the order and then the species. The principle of number counting was that the brittle nemertean and polychaete worms were counted by head, while the empty mollusk shells were excluded. Group specimens were not counted by number. After classification and counting, wet samples were dried on filter paper and then weighed for biomass determination.
Seawater sampling was carried out when the high tide was falling. Seawater samples were taken in a 5-L plastic tank at a depth of 1 m below the sea surface in the low tidal area, and were then stored for later laboratory analysis. We collected 1-L surface seawater and added about 5 mL of 10 g/L MgCO3suspension solution to allow chlorophyll a (chl a) analysis. Water temperature, salinity and pH were measured at the sampling sites with a thermometer, Orion 130A salimeter and pH meter, respectively. The nitrite content was determined by using the N-(1-naphthyl)-ethylenediamine dihydrochloride spectrophotometric method. The concentration of PO43ˉ was determined by using P-Mo-V xanthic acid spectrophotometry.NH4+, NO3ˉ, SiO32ˉ and chl a contents were determined by the hypobromite oxidation method, Zn-Cd reduction method, Si-Mo spectra photometry and acetone extraction spectrophotometric method,respectively.
2.2 Species statistics and measuring methods for biological diversity

Species evenness was calculated according to the formula J= H′/(log2S), where H′ is the value of the former diversity index and S is the total number of species in the samples.
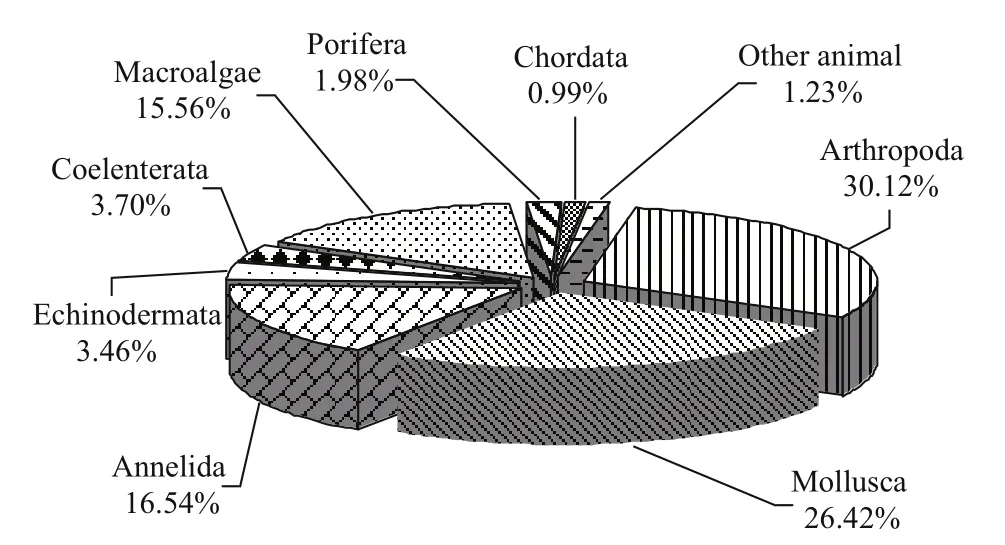
Fig.2 Species composition of macrobenthos in the intertidal rocky zone of Naozhou Island, China
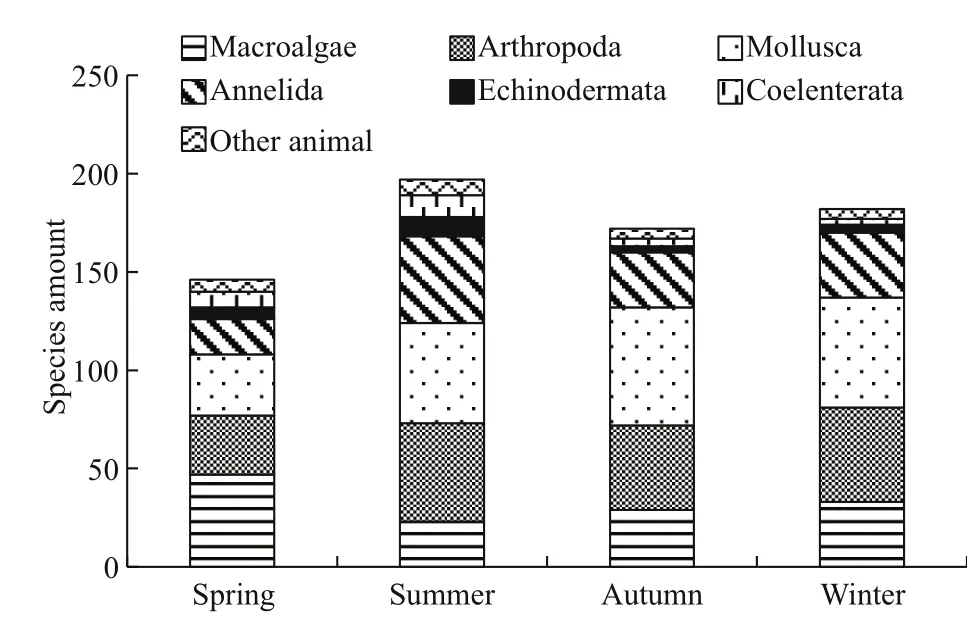
Fig.3 Seasonal changes in species number of macrobenthos in the intertidal rocky zone
The dominance index was determined using the formula Y=( ni/ N)× fi, where niis the total biomass of the i-th species, N is the total biomass of all species, fiis the i-th frequency at each station and the dominant species are selected by Y >0.02.
The species richness index was calculated using the formula d=( s-1)/log2N, where N is the total biomass of whole samples and s is the total number of species.

The Jaccard index is calculated using the formula P= c/( a+ b- c), where a and b are the number of benthic species in two diff erent seasons, and c is the number of benthic species in two diff erent seasons.The stability analysis of a species community is based on biomass abundance comparison; that is, by drawing the K-dominance curves of biomass and density into the same figure, and comparing the inhabitant density and biomass index to determine the disturbance of benthic communities. The abundance biomass comparison (ABC) curves of communities were analyzed using Primer 5.0 software.
3 RESULT
3.1 Benthic species composition in the reef zone
We identified 407 macrobenthic species; their species names and taxonomic categories are tabulated in Appendix I. The distribution of all species was: 64 macroalgae (accounting for 15.72% of total species);1 Angiospermae (0.25%); 123 Arthropoda (30.22%);107 Mollusca (26.29%), 67 Annelida (16.46%), 15 Coelenterata (3.69%), 14 Echinodermata (3.44%), 8 Porifera (1.97%), 4 Chordata (0.98%), 2 Urochordata(0.49%), 1 each of Echiura and Sipuncula (0.25%)(Fig.2). Arthropoda, Mollusca, Annelida, and macroalgae comprised 88.70% of the total species,and thus were the four primary benthic groups in the Sargassum field of the reef areas.
The macrobenthos species and their abundance in the Sargassum field of Naozhou Island varied as the seasons changed. The number of species was higher in summer, when 197 species were identified, whereas fewer species were found in other seasons (182 species in winter, 172 in autumn and 146 in spring).The number of species within the same category showed significant seasonal variation. For example,47 species of macroalgae were found in spring, but only 33 species in winter, 29 in autumn and 23 in summer. We found 50 Arthropoda species in summer,48 in winter, 43 in autumn and 30 in spring. The Mollusca had the largest number of species, as there were 60 species in autumn, 56 in winter, 51 in summer and 31 in spring. We found 44 species of Annelida in summer, 33 in winter, 28 in autumn and only 18 in spring (Fig.3).
A total of 44 species were detected in all four seasons: 9 macroalgae, 14 mollusks, 13 arthropods, 6 Annelida, 1 Coelentera and 1 Echinodermata. The total species found in two seasons were in the range of 66-105, associated with the Jaccard index ranging from 0.262 to 0.422 (Table 1). This shows that the highest index appeared in autumn/winter, and the
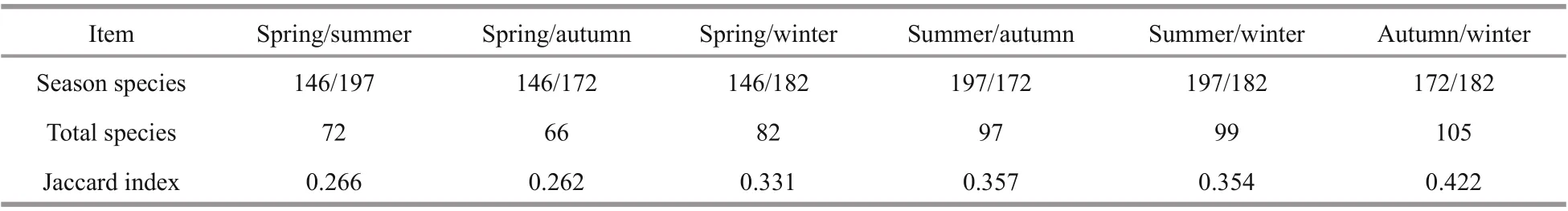
Table 1 Jaccard Index of macrobenthos species between two seasons in the rocky zone of Naozhou Island, China
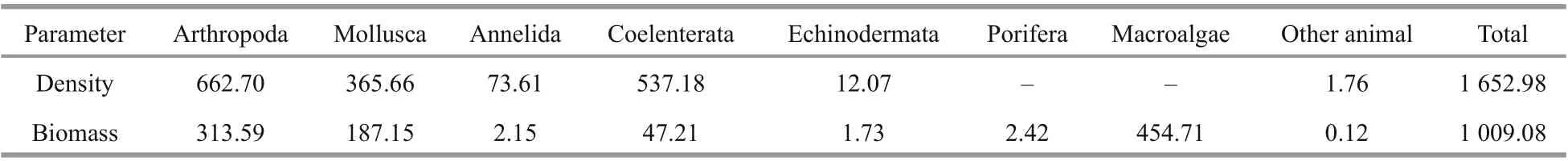
Table 2 Average inhabitant density (inds./m 2) and mean biomass (g/m 2) of various benthic groups in the investigation area
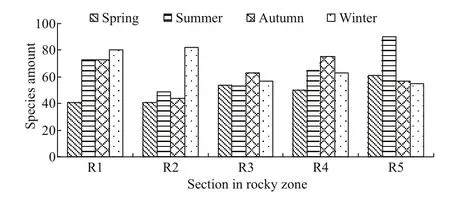
Fig.4 Distribution and seasonal variation of benthic species in each section in the rocky zone
3.2 Density and biomass distribution of benthos
3.2.1 Density and biomass of various benthic groups
The average density of species inhabiting the investigated area, by category, was in the order Arthropoda > Coelenterata > Mollusca > Annelida >Echinodermata > other animal. The mean biomass was in the order macroalgae > Arthropoda > Mollusca> Coelenterata > Porifera > Annelida > Echinodermata> other animal (Table 2).
3.2.2 Distribution of benthic community composition
There was spatial variation in the species distribution of macrobenthos in the Sargassum field off Naozhou Island. The average species number in each sampling section season was in the order R1(67 species) > R5 (66 species) > R4 (63 species) > R3(57 species) > R2 (54 species) (Fig.4). The total yearly species number in each sampling section was in the order R1 (183 species) > R3 (158 species) > R4(155 species) > R5 (154 species) > R2 (143 species).The R1 sampling section was the highest in both total species and average numbers, whereas the R2 sampling section was the lowest in both total and average species numbers.

Fig.5 Seasonal and spatial variation in benthic inhabitant density in the rocky zone
There was significant seasonal and spatial variation in the distribution of macrobenthos density in the Sargassum field off Naozhou Island between the sampling sections and seasons. Mean inhabitant density among the sampling sections was in the order R5 (3 242.0 inds./m2) > R3 (2 227.9 inds./m2) > R2(1 252.8 inds./m2) > R4 (882.7 inds./m2) > R1(809.4 inds./m2). The seasonal variations in inhabitant density was in the order summer (2 786.4 inds./m2) >autumn (1 986.8 inds./m2) > spring (1 033.7 inds./m2)> winter (924.9 inds./m2) (Fig.5).
The biomass of macrobenthos in the gulfweed field off Naozhou Island varied greatly among the seasons and sampling sections. The general order was R5(1 684.6 g/m2) > R3 (1 182.8 g/m2) > R2 (1 045.6 g/m2)> R1 (575.9 g/m2) > R4 (531.1 g/m2). There was also seasonal variation in each sampling section (Fig.6).
3.2.3 Spatial changes in the composition of benthic community
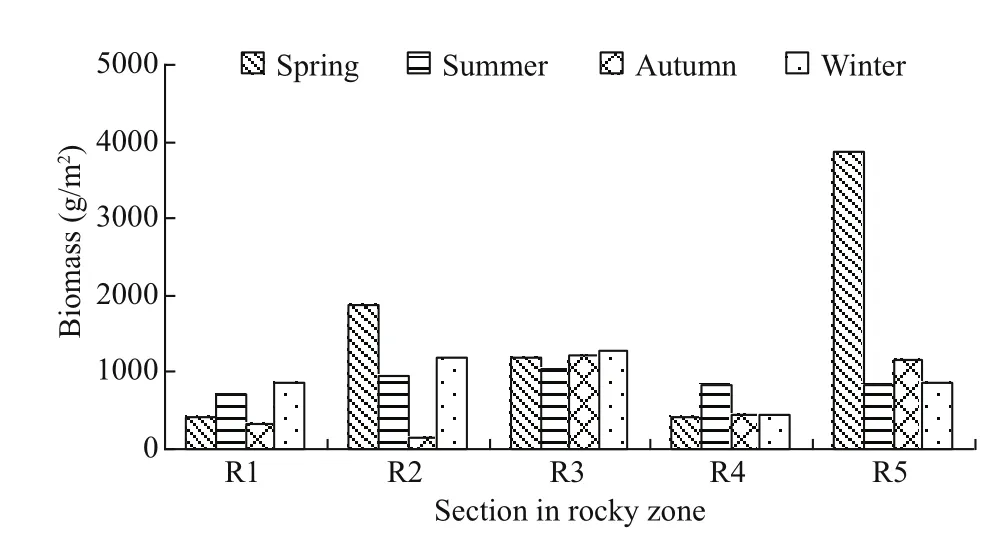
Fig.6 Seasonal variation in benthic biomass distribution in the gulfweed field
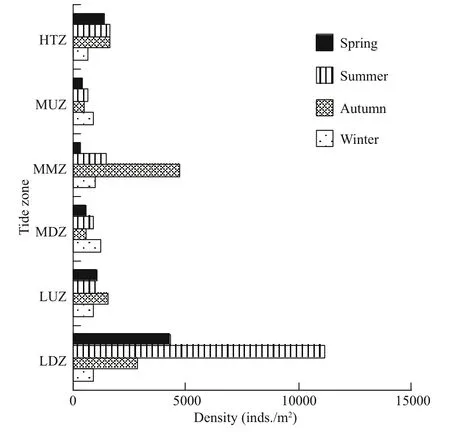
Fig.7 Spatial distribution and seasonal variation in benthic inhabitant density in the rocky zone
The composition and abundance of macrobenthos species displayed clear spatial variation in diff erent tidal zones in the gulfweed field off Naozhou Island.The species number was in the order high tidal zone(13 species) < upper middle tidal zone (99 species) <middle tidal zone (104 species) < lower middle tidal zone (126 species) < upper low tidal zone (131 species)< low tidal zone (169 species). This spatial variation in species distribution was largely due to the tidal influence and the wave breaking strength. The annual average inhabitant density of the benthos was also variable among tidal zones, and displayed the order lowtidal zone(4810.3inds./m2)>middletidal zone(1 863.5 inds./m2) > hightidal zone (1 334.8 inds./m2)> upper low tidal zone (1 129.5 inds./m2) >lower middle tidal zone (814.6 inds./m2) > uppermiddle tidal zone (615.4 inds./m2). The spatial change in the inhabitant density among the tidal zones was mainly due to changes in species and inhabitant conditions in diff erent tidal zones (Fig.7).
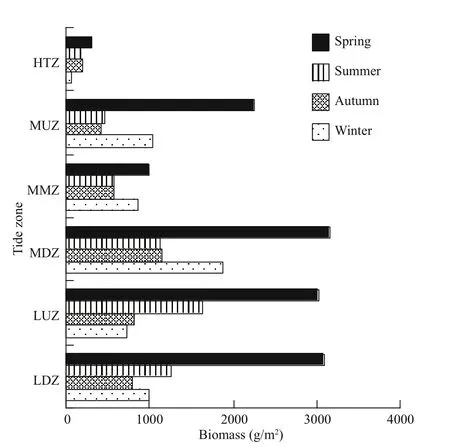
Fig.8 Spatial and seasonal variation of benthic average biomass in tidal zones
The biomass of macrobenthic organisms also displayed spatial change among the tidal zones, as the annual average biomass in diff erent tidal zones was in he order oflow tidal zone (1 539.4 g/m2) > middle idal zone (1 825.4 g/m2) > upper low tidal zone 1 555.9 g/m2) > upper middle tidal zone (1 043.9 g/m2)> middle tidal zone (754.0 g/m2) > high tidal zone 180.2 g/m2) (Fig.8). The spatial distribution in macrobenthic biomass also showed temporal variation as the seasons changed. This was mainly because of seasonal change in the dominant species of macroalgae, as well as the local harvest of economic species.
3.3 Dominant species and seasonal variation
Dominance indices of benthic organism species in tidal zones off Naozhou Island were determined statistically according to the biomass of each species.We identified 15 dominant species: 8 macroalgae, 5 mollusks and 2 arthropods. Examination of the seasonal distribution of dominant species found 5 in spring, 6 in summer, 7 in autumn and 5 in winter.There were no species found to be dominant through all four season, but only Sargassum hemiphyllum,Pterocladia capillacea, Tetradita squamosa squamosal, and Clypemorus bifasciatus are found to be the dominant species over three seasons (Table 3).
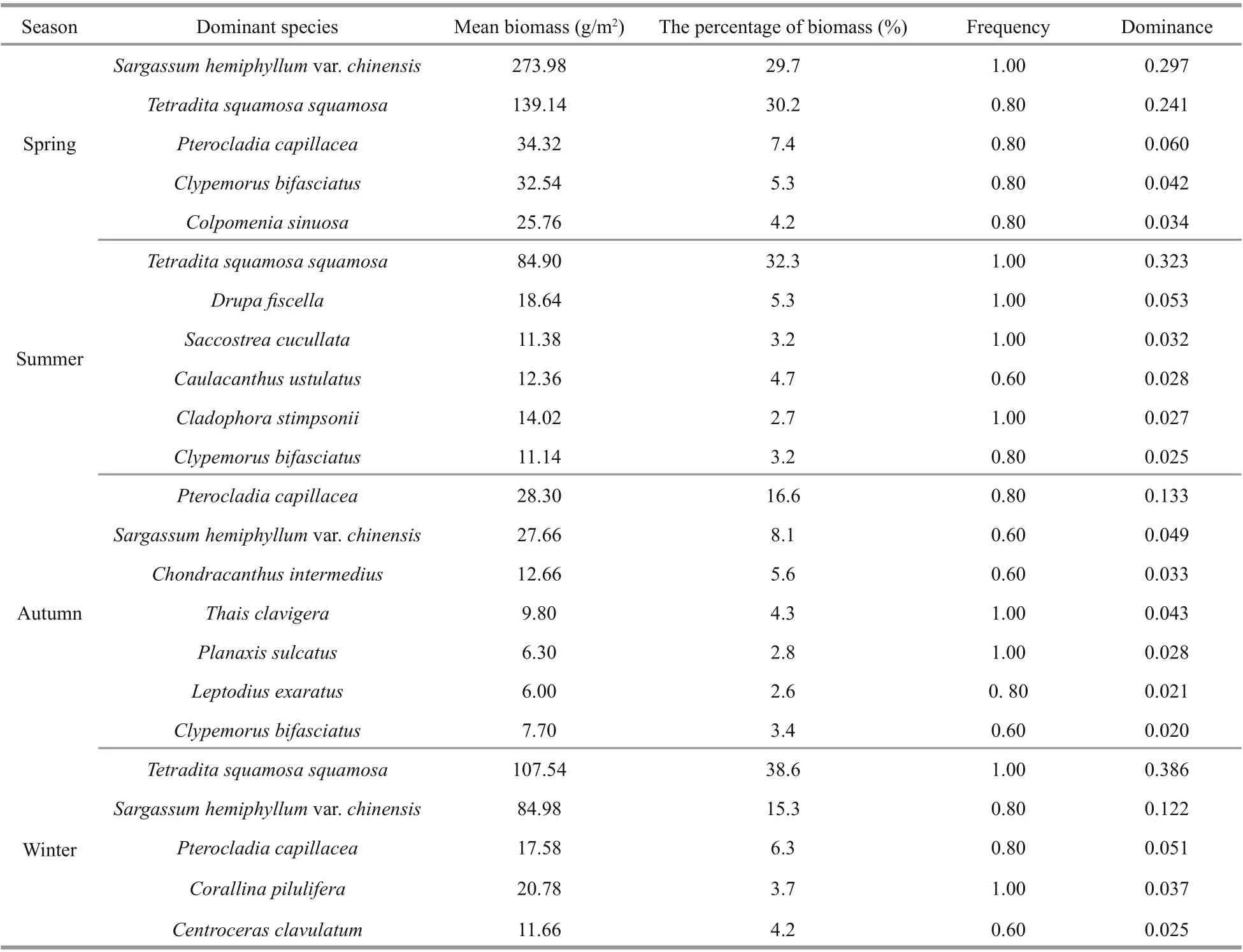
Table 3 Seasonally dominant species in the Sargassum field off Naozhou Island
3.4 Diversity of benthic species
Biodiversity parameters for the macrobenthic algae in the rocky zone off Naozhou Island (calculated according to the methods described in Section 2) are listed in Table 4. Analysis of the ecological parameters of macrobenthos in the tidal zones off Naozhou Island, based on the biomass of each species, showed that the diversity index was in the range of 1.21-4.42,with an average of 2.80. The highest diversity index occurred in autumn and the lowest in spring. The evenness index varied from 0.22 to 0.71 with the highest value in autumn and the lowest in spring. The species richness varied from 2.97 to 7.24 with the highest in autumn and the lowest in spring. The Simpson dominance index was in the range 0.20-0.92, with the highest in winter and the lowest in spring (Table 4).
3.5 Macrobenthic distribution and seasonal change
The distribution of macrobenthic species in the gulfweed field was heavily disturbed by humanactivities. The Bohelan reef area (R5 section) showed the highest annual average biomass as it was furthest from the land and less disturbed by human activities.R1 was heavily aff ected by human activities and displayed the lowest annual average biomass. The annual average biomass of macrobenthic organisms in the five sampling sections were in the order R5(776.01 g/m2) > R4 (506.53 g/m2) > R2 (438.90 g/m2)> R3 (422.92 g/m2) > R1 (129.20 g/m2).
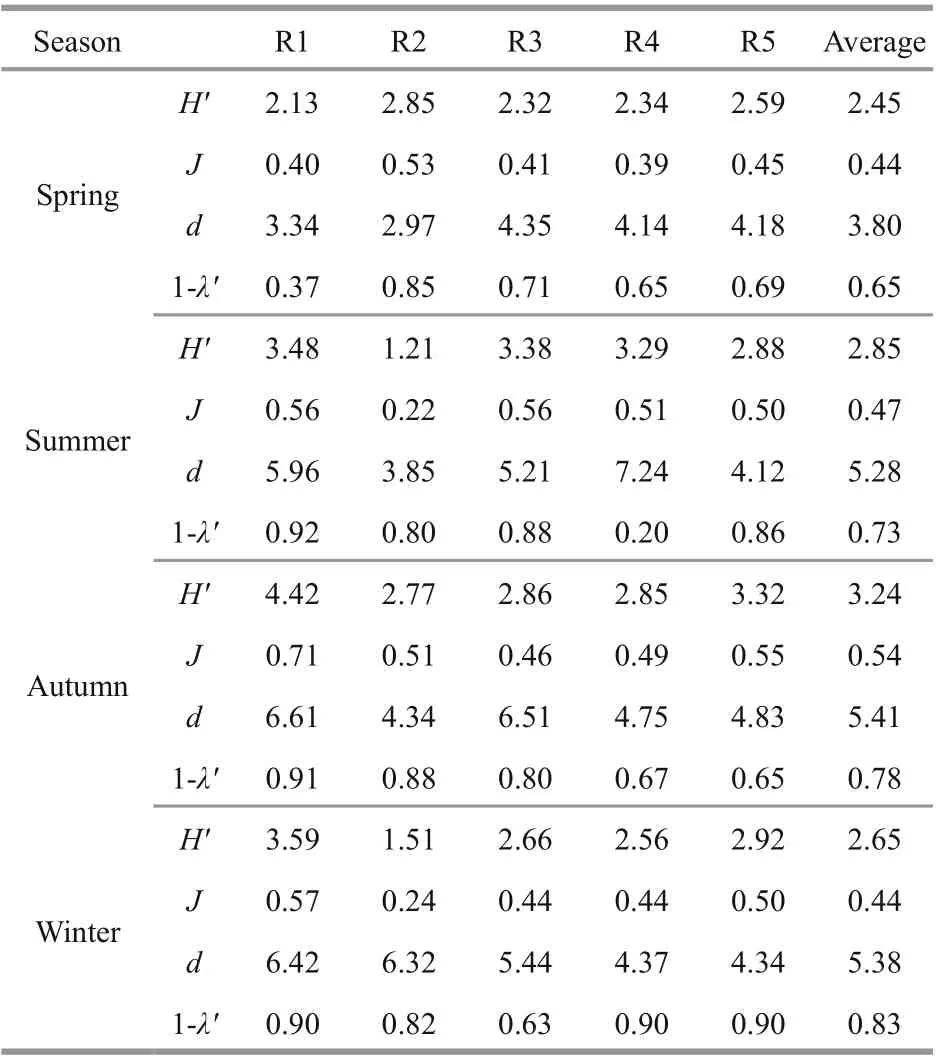
Table 4 Biodiversity parameters for the macrobenthic algae in the rocky zone off Naozhou Island, China
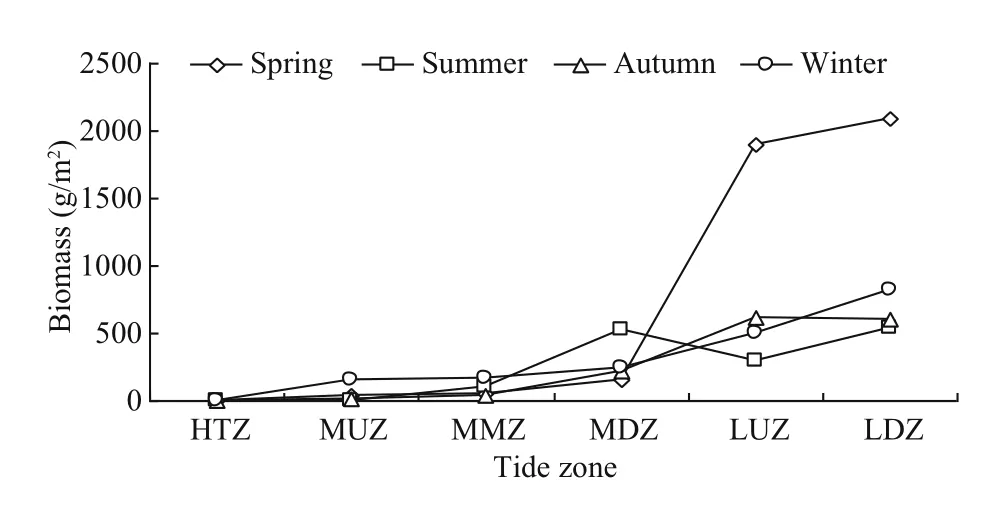
Fig.9 Vertical distribution of macroalgal biomass in each season
There was clearly spatial variation in the distribution of macroalgae species, as 1 macroalgae species was found in the high tidal zone, 8 in the upper middle tidal zone, 10 in the middle tidal zone,16 in the lower middle tidal zone, 22 in the upper low tidal zone and 32 in the low tidal zone. Annual average biomass (inds./m2) in each tidal zone was: 1 009.82 in the low tidal zone, 827.45 in the upper low tidal zone,284.84 in the lower middle tidal zone, 93.41 in the middle tidal zone, 51.15 in the upper middle tidal zone and 0 in the high tidal zone (Fig.9).
The macroalgae displayed obvious seasonal preferences, as lower temperature species such as S. hemiphyllum, S. naozhouense, S. integerrimum, S.henslowianum, Hizikia fusiforme and Asparagopsis taxiformis grew vigorously in winter. These dominant species grew faster during the cooler temperature in winter and reached their maximum biomass in spring,then gradually died out in late spring. Hightemperature species, including Rhizoclonium implexum, Cladophora stimpsonii, Galaxaura oblongata, Neogoniolithon sp., Padina australis, and Bryopsis harveyana, appeared in summer. Of these high-temperature species, the first four were found only in summer, P. australis occurred in spring, and disappeared in autumn, whereas B. harveyana occurred in summer and disappeared in winter.Eurythermal macroalgal species such as S. cinereum,Pterocladia capillacea, Valonia utricularis,V. aegagropila, Caulacanthus ustulatus, Corallina sp., C. pilulifera, Chondracanthus intermedius, and Centroceras clavulatum were found in all seasons.The numbers of macroalgal species found in each of the four seasons were: 47 in spring, 33 in winter, 29 in autumn and 23 in summer. The average biomass was 848.14 g/m2in spring, 378.57 g/m2in winter, 297.99 g/m2in autumn and 294.15 g/m2in summer (Fig.10).
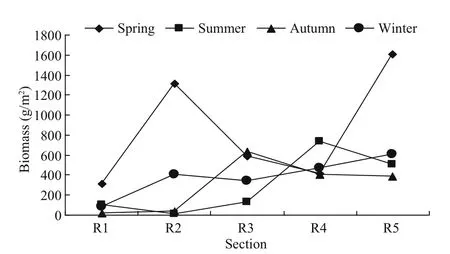
Fig.10 Seasonal change in macroalgal biomass in each section investigated
4 DISCUSSION
4.1 Benthic community characteristics
The features of the sea bed play an important role in the environmental selection of benthic organisms.The gulfweed field off Naozhou Island has a rocky bed, and its benthic communities are characterized by large numbers of sessile, attached, crawling, adhesive and cave-living organisms, as well as various creatures living or hiding in the gaps of the reef rocks. In the field we investigated, sessile organisms included 8 species of Porifera, 15 species of Coelentera, 7 species of Mollusca, 8 species of Arthropoda and 2 species of Ascidiacea. The attached organisms included 64 macroalgae species and 10 species of Mollusca, whereas crawling and adhesive organisms included 19 species of Mollusca, as well as four species of boring benthos and cave species. The benthic species living on rock beds accounted for 33.50% of total benthic species. The number of species living in rock gaps, coral reefs and seaweed and under oyster shells comprised 38 polychaetes,34 mollusks, 75 crustaceans, 9 echinoderms, and 2 chordates; these species accounted for 38.92% of the total benthic species. The 15 dominant species with the specific ecological habits of adapting to the rocky bottom consisted of 2 sessile species, 8 attaching species and 5 species living on the rocks and coral reefs. A detailed observation has shown that speciesliving in loose sediments, such as members of the genus Nereis, fail to adapt to hard rocky beds, as this habitat cannot provide suitable living conditions(Zhang and Sun, 2007). Polychaete species accounted for only 16.50% of the total species, and mean biomass accounted for only 0.19% of the total biomass. Clearly, a rock basement is suitable for a large number of sessile, attached, adhesive, crawling and cave-living organisms, as well as for various species living or hiding in coral reefs. Their total species numbers, inhabitant density and biomass are greatly exceeded the mollusks and large marine macroalgae living on the other types of basements (Li et al., 1993; Lai et al., 2019). The rocky basement also provides a point of fixation for porous animals,cnidarians and sea squirts, and off ers hiding and living space to crabs; while the thick seaweed near the basement in the lower middle tidal zone provides an excellent habitat for numerous small crustaceans. The number and density of crustaceans, therefore, make up the largest proportion of the ecosystem, while their biomass is second to that of macroalgae.

Table 5 Comparison of macrobenthic diversity off Naozhou Island and other ocean areas
With the high primary productivity and unique food web feeding on macroalgae, nutrients can be used by macroalgae for manufacturing organic matter by photosynthesizing. The mean biomass of macroalgae accounts for 40.53% of the average macrobenthic biomass throughout the world’s seas.The dominant benthic species comprised eight species of macroalgae (Table 1) and accounted for half of the dominant species in this oceanic region. Their biomass and dominance indices show that they comprised the majority of all organisms in spring, autumn and winter, but not in summer. In spring, the biomass of macroalgae accounted for 45.81% of macrobenthic biomass, and S. hemiphyllum and Brassica chinensis were the most important structural organisms. Species that depend directly on seaweed include Anthocidaris crassispina, Temnopleurus toreumaticus, Haliotis diversicolor, Monodonta labio, Chlorostoma rustica,Trochus chloromphalus, T. maculatus, Turbo brunneum, Nerita yoldii, Nerita albicilla, Batillaria zonalis, B. cumingi, Mauritia arabica, Amphitoe valida, Sunamphitoe plumosa, Cymodoce japonica,Charybdis japonica, Sphaeroxius nitidus, Pisidia serratifrans, Hyastenus pleione , and Ligia exotica. It is estimated that about 10% of primary production passes directly into the food web by feeding, and the remaining 90% enters the food chain as debris or dissolved organic matter (Komatsu and Kawai, 1986;Komatsu et al., 2003).
The rocky sea floor provides a high degree of spatial heterogeneity for various benthic organisms,so it is very rich in biological resources (Jiang, 2015),and its species diversity is very high (Zhang and Sun,2007; Liu, 2013). A comparison between a nature reserve on Nirayama Island and the intertidal zone in Nanji Archipelago revealed that the macrobenthos and macroalgae species in a seaweed farm offNaozhou Island were more diverse, as the diversity index of the macrobenthos and macroalgae was higher than off other islands (except for the Nanpeng Islands)(Table 5). It indicates that species diversity is related to latitude, and that the lower the latitude, the richer the species diversity.
The species living in the high tidal zone were highly adaptable organisms such as Littoraria articulata, Nodilittorina radiata, N. trochoides,Onchidium verruculatum, Uca arcuata, U. lactea,Sesarma plicata, and Ligia exotica, which can tolerate dramatic fluctuations in their environment. Some species living in the high tidal zone, such as Amphibalanus amphitrite, Chthamalus malayensis,and Capitulum mitella, are able to breathe air directly,tolerate drying and extremely high temperatures, and can withstand exposure to air for a long time. Although a large number of small, mobile crabs live in the upper middle tidal zone, their benthic inhabitant density there is very low as there are few sessile and attached species, and these small crabs migrate with the tide.However, because Saccostrea cucullata lives in this region, the biomass of the upper middle tidal zone is slightly higher than that of the middle tidal zone. The benthic habitat density is high, but the biomass in the middle tidal zone is lower, as large numbers of Tetradita squamosa squamosa, Clypemorus bifasciatus, Planaxis sulcatus, and Typosyllis variegata, which have a small biomass, live in the middle tidal zone. The lower middle tidal zone is dominated by a community comprising Tetradita squamosa squamosa, Clypemorus bifasciatus,Leptodius exaratus, Pterocladia capillacea, and Hyale schmidti. The vigorous growth of Tetradita squamosa squamosa and Pterocladia capillacea inhibits some sessile and attached species, whereas the changes in the amount of dominance of Tetradita squamosa squamosa and Leptodius exaratus play an important controlling role in the benthic biomass of the lower middle tidal zone.
The low tide zone is submerged beneath the water for most of the time. Benthic algae with weak antidehydration tolerance grow in clusters and form dense clumps of seaweed. Sargassum hemiphyllum,Corallina pilulifera, Colpomenia sinuosa, and Cladophora stimpsonii, for example, mainly grow in the low tidal zone, while Pterocladia capillacea,Centroceras clavulatum, Caulacanthus ustulatus, and Chondracanthus intermedius can grow in both the lower middle tidal zone and the low tidal zone. The few low-adaptability species, such as sponges,cnidarians and echinoderms, mainly inhabit the low tidal zone where the air exposure time is very short.Many mobile species retreat with the tide into the lower tidal zone; the dominant species Thais clavigera, Drupa fiscella, and Leptodius exaratus, for instance, are mainly distributed in the area between the lower middle tidal zone and the low tidal zone.Because of their strong space competitiveness,macroalgae form dense bands in low tidal areas and therefore inhibit benthic animal growth and distribution. The distribution of small bivalve and algae-feeding gastropods in the low tidal zone is clearly inhibited through predation by carnivorous gastropods, such as Thais clavigera (Zhang et al.,2000). Macroalgae cannot be counted individually,but their benthic biomass in the low tidal zone is high although the inhabitant density is not high. The vertical distribution of benthic organisms in diff erent sampling sections varies. The vertical zoning of benthic organisms in the intertidal zone of a rocky reef area broadens and becomes deeper as wave strength increases. Surf species of Sargassum in reefs,for example, grow vigorously where waves are strong,and this phenomenon is more apparent in off shore islands (Zheng et al., 2004; Peng et al., 2009). The vertical zoning of macroalgae is closely correlated with the intensity oflight and transparency of seawater. Macroalgae mainly absorb the complementary light of their own color. For example,the red and part of the purple light in the daylight spectrum is mostly necessary to Chlorophyta, the orange and part of the yellow light is mainly used for photosynthesis in brown algae, whereas red algae use mainly yellow and green light for photosynthesis.Therefore, the vertical zoning of algae varies greatly,depending on their usage oflight from the daylight spectrum.
The number of benthic species is very variable in diff erent seasons in the investigated region. The high temperatures of summer are suitable for various benthic animals, and this is in agreement with the finding that the lower latitudes (higher temperature)are typically associated with greater biodiversity.However, the number of macroalgal species is at its highest in spring and lowest in summer (Fig.3). One reason for this is the strong space competitiveness of macroalgae as they spread and occupy most of the bed of a rocky reefin spring. Reproduction of benthic flora and fauna also influences the variations in seasonal biomass. Benthic animals mostly breed in spring when the gonads are full, thus their biomass is highest in spring. Because the temperature range of macroalgal growth is narrow, the biomass of macroalgae shows significant seasonal variation.Most macroalgae flourish in winter and spring, and die out in summer, so their biomass reaches its maximum in spring and minimum in summer. For example, Sargasso germinates in late autumn; the growing season is 1-3 months, it flourishes in mid-April and dies and decomposes in late May. The high temperatures in summer and low temperatures in winter will force some active species to hide to avoid extremely hot or cool temperatures. For example,some benthic species, such as Thais clavigera,Chlorostoma rusticum, and Pachiyarapsus crassipes,often hide or gather in rock gaps, under rocks or in pools in hot summers, and migrate to the low tidal zone in cold winters. Seasonal changes in the species and biomass of benthic organisms reflect their adaptability to changes in water temperature, as well as to their growth and breeding features (Fig.11).
Among the sampling sections, the total number of all species in R1 was the largest because of the multiple nature ofits rocky, sandy and silty seafloor.The total number of all species in R2 was the lowest,as this section was close to a popular beach where fishing has a greater impact on the species. It has been illustrated that spatial heterogeneity and human activities may be significant factors aff ecting the distribution and diversity of species and communities.Sampling section R5 had the largest number of total species and biomass because it was far from the coastline and experienced limited disturbance from human activity, in particular during spring (Fig.6).This demonstrates the significance of environmental and human activity factors, and also indirectly explains the reason for seaweed field degradation.The regional distribution of benthic organisms is,therefore, obvious.
4.2 Structure of the macrobenthic organism community and aff ecting factors
Physical, chemical, environmental, human and many other factors aff ect the structure of the macrobenthic community in intertidal zones. As the transition unit of the land-sea interaction system, the intertidal zone is highly unstable (Shi, 2001).Following the increasing need for high quality nutritious and delicious seafood, the goal of fishery was far beyond the virtually self-suffi cient and it is more economic activity. Fishing increased at an unprecedented rate and scale, resulting in considerable damage to the marine organism communities and environments (Peng et al., 2009). Consequently, the numbers of not only economic species, but also noneconomic species, fell greatly or even disappeared.These species included Haliotis diversicolor,Barbatia virescens, Perna viridis, Saccostrea cucullata, Ruditapes variegata, R. philippinarum,Monodonta labio, Chlorostoma rustica, Lunella coronata granulata, Polinices sp., Rapana venosa,Murex trapa, Thais clavigera, T. luteostoma, Octopus ocellatus, Octopus sp., Capitulum mitella,Oratosquilla oratorta, O. inornata, Anthocidaris crassispina, Temnopleurus toreumaticus, Sargassum hemiphyllum, Hizikia fusiforme, S. naozhouense, S.henslowianum, S. maclurei, Charybdis cruciata,Portunus pelagicus, P. sanguinolentus, P.trituberculatus, and Scylla serrata. A previous study also showed that macrobenthos of high economic value, such as Charybdis cruciata, Portunus pelagicus, P. sanguinolentus, P. trituberculatus, and Scylla serrata, used to live in the intertidal zone but now can only be found below the tidal zone (Xiao and Li, 2003). One consequence is that non-economic species with a short life cycle, resilient reproductive capacity and wide distribution, such as Tetradita squamosa squamosa, Clypemorus bifasciatus,Septifer virgatus, Pterocladia capillacea, Colpomenia sinuosa, Caulacanthus ustulatus, Cladophora stimpsonii, Chondracanthus intermedius, Corallina pilulifera, and Centroceras clavulatum, have replaced economic species and become dominant, deepening the structure of the secondary community. Overfishing of fish and large crabs at the upper part of the food chain has destroyed the food chain, leading to the extensive reproduction of algae-eating animals including Ampithoe. Secondary species, such as the scale barnacle Tetraclita squamosa, have become dominant populations, occupying a large number of rock reef surfaces. This eventually resulted in large algae losing their attached bases and suff ering serious damage. In addition, shrimp farming and sewage discharge have polluted the reef area off Naozhou Island, especially in the late summer and during the late autumn to early winter. An increasing amount of effl uent carrying suspended solids and nutrients from shrimp farming has been discharged into the surrounding ocean, causing significant damage to the benthic macroalgae in the rocky reef area. During the germination season of macroalgae (especially gulfweed), in late autumn, the suspended solids and phytoplankton (especially benthic diatoms) from farming and aquacultural wastewater were deposited onto the surface of benthic macroalgae in the gulfweed area. This resulted in serious problems for the growth and reproduction of macroalgae, as the deposits blocked light from reaching the plants, aff ecting the adhesion of spores, and the germination and growth of buds. Furthermore, global warming has led to severe degradation of constructive species of Sargasso communities in the Sargasso field, simplifying the species comprising the benthic organism community in the intertidal zone. For example, the number of macroalgae species was 87 in 2006, but there are 64 species found at present, and the once-dominant species Gelidium pusillum, Valoniopsis pachynemam,and Chaetomorpha media and one ordinary species of Oscillatoria sp. have vanished. Other formerly dominant species, such as Asparagopsis taxiformis,Gigartina intermedia, Centroceras clavulatum, Ulva conglobate, and Enteromorpha intestinalis have now become rare (Hu, 1997; Zhao et al., 2006).
The seasonal variation in the physical and chemical conditions of the Sargasso field off Naozhou Island has significantly impacted macrobenthic growth,reproduction, spatial distribution, migration and seasonal succession. Diff erent organisms have their own life cycles and life histories, and their biomass and density are also influenced by their own habits,intraspecific and interspecific competition, food relationships, habitats, and seasonal succession.
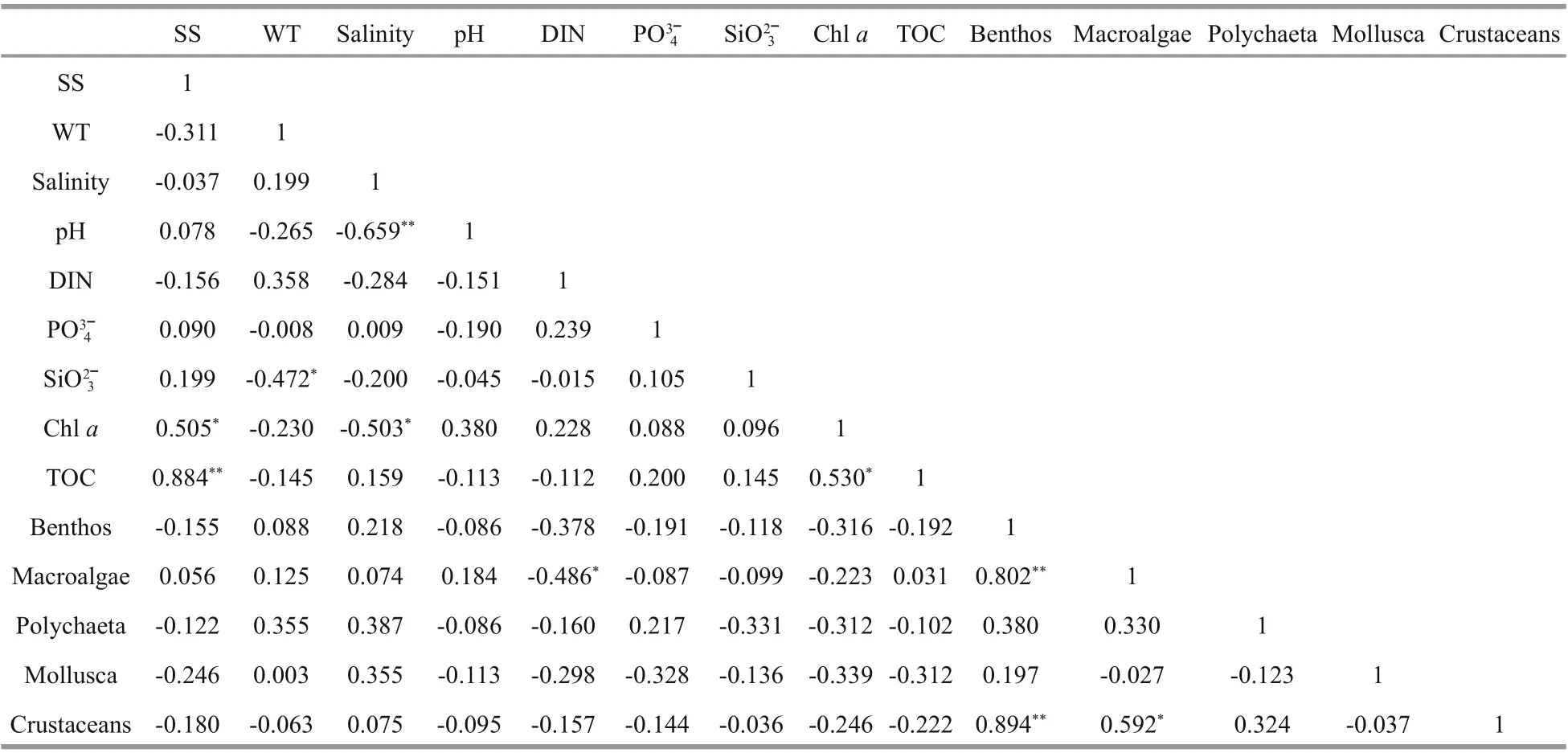
Table 6 Correlation between macrobenthic biomass and environmental factors
The horizontal distribution ofintertidal organisms is mainly aff ected by the impact of waves and currents(Cai et al., 1990; Yang and Chen, 1996). Species living in areas with waves, such as Tetradita japonica,T. squamosa squamosa, Littorina, and Sargassum,can grow and live very well in the surf zone at high inhabitant densities and biomass. Some species, such as Littorina, can even grow in the supralittoral zone about 2.5 m from the high tidal zone. The upper boundary of the vertical distribution ofintertidal creatures is related to zonation, whereas environmental factors (physical factors), such as dryness and temperature changes caused by waves and tides, also play a vital role. The lower boundary is controlled by biological features such as spatial competition between species and the ability to tolerate the invasion of harmful organisms (Zheng et al., 2004).
Analysis of the biomass and environmental factors of benthos, macroalgae, polychaetes, mollusks, and crustaceans in each sampling section and in each season revealed that the biomass of benthos,macroalgae and crustaceans were well correlated to each other, with correlation coeffi cients of 0.802,0.894 and 0.592 ( P <0.05, 2-tailed, n= 20, the same below) (Table 6), respectively. This indicated that crustaceans and macroalgae were the major contributors to benthic biomass. The variations between growth and decline of number and biomass of crustaceans were determined by the macroalgae biomass, suggesting the presence of an important food chain relation between crustaceans and macroalgae. Our analysis also found that there was no clear correlation between the biomass of benthos and the temperature and salinity of seawater; this may be related to the massive growth of macroalgae in winter and spring, which vanished in summer. The benthic biomass, pH, DIN (includes nitrate, nitrite and ammonia), phosphates, silicates, chl a, and TOC(total organic carbon) displayed weak negative correlations, indicating that the benthos directly or indirectly relied on the nutrients, chl a and dissolved organic carbon in the seawater. A negative correlation coeffi cient of 0.486 between macroalgae and DIN indicated that algae need to absorb DIN from water to enable growth, as DIN is in its lowest in spring and winter when macroalgae grow vigorously. This contrasts with other marine regions where nutrients accumulate over winter. Macroalgal biomass also showed a weak negative correlation with seawater phosphate and silicate content, indicating a competitive relationship between macroalgae and chlorophyll, and suggesting that these three factors—in addition to the main factor of DIN—also aff ected macroalgae growth. Polychaetes showed a weak positive correlation with temperature and salinity,indicating that their growth and reproduction are aff ected by these two factors. Although there was a close food chain relationship, there was a weak correlation between the biomass of polychaetes and that of algae and crustaceans. This was because most macroalgae live where there are waves, and the organic matter created as they rot washes away and cannot readily be used by polychaetes. Some polychaetes, such as species in the genus Nereis,even feed on macroalgal rhizoids (Sun et al., 2012).Mollusk biomass displayed a clear negative correlation with pH, DIN, phosphate, silicate, chl a,TOC, macroalgae, polychaetes, and crustaceans. It had a weak positive correlation with temperature and salinity, indicating that the growth of mollusks depends on nutrients, chl a and dissolved organic carbon from seawater. There are competitive relationships among mollusks, macroalgae,polychaetes, and crustaceans, while the growth and reproduction of mollusks is aff ected by temperature and salinity.
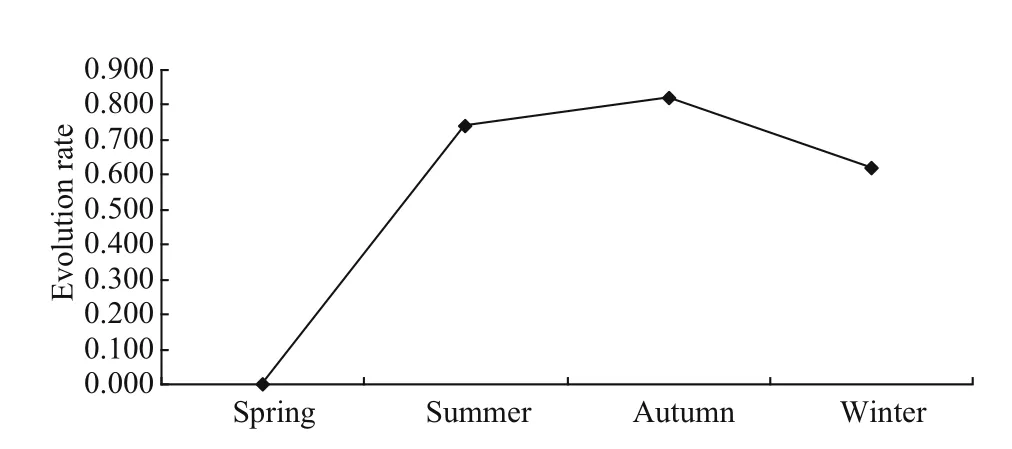
Fig.11 Seasonal evolution trends of macrobenthic communities in the investigated area
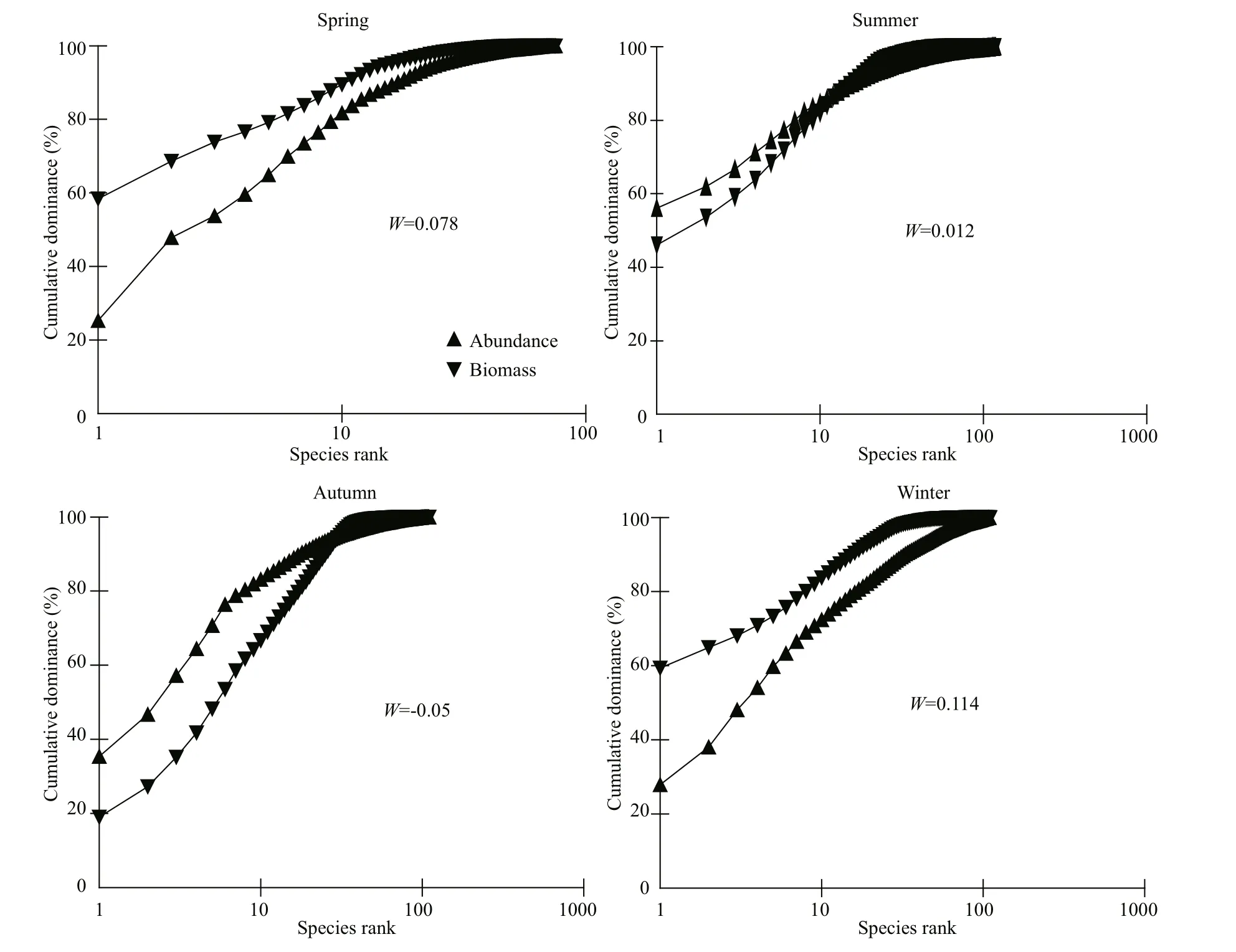
Fig.12 Macrobenthic species abundance and biomass dominance curve in each season
4.3 Stability of community structure
Using the ABC method, we analyzed macrobenthic species abundance (except for that of macroalgae and porous animals) and biomass (although there are no abundance data for groups, macroalgae and Spongia)(Warwick, 1986) (Fig.12). The results showed that the stability of the macrobenthos community structure varied in diff erent seasons. The abundant cumulative dominant curve mainly lay above the biomass accumulation curve, but intersections or overlapping also occurred in the summer and autumn. These results indicated that the species and number composition of small benthic organisms were those of dominant organisms that had been disturbed. The initial cumulative superiority of the abundance curve was lower and suggested that the species abundance superiority of benthic organisms was less obvious in summer and autumn, and that there was no flourishing of small individuals or group species. This also suggested that the impact of the disturbance was still in its early stages. However, the initial cumulative dominance curve in summer was higher than that in autumn; this may explain the fact that the size of an individual organism was smaller in summer but was associated with higher inhabitant density. The abundance cumulative dominance curve lay below the biomass accumulation curve in winter and spring,suggesting that the benthic community has remained stable during the inter winter and spring seasons.
Because Naozhou Island is far away from the coast of the Chinese mainland, disturbance to the gulfweed field off the island was mainly from effl uents discharged by shrimp aquaculture and increasing human activities. Late summer and late autumn to early winter are the harvest seasons for shrimp, and shrimp farming often discharges a huge amount of farming sewage to the coastal area in the late breeding season and at harvest. This results in an increase in suspended particulate matter in seawater and consequently a decline in seawater transparency,deterioration in water quality and disturbance to some sensitive species in the gulfweed farm. The activities of human harvesting have seasonal variations, and their frequency decreases during the colder seasons of winter and early spring, owing to the lower water temperatures. As a result, mollusks and crustaceans have more opportunity to grow, and their biomass increases. In summer and autumn, in contrast, frequent human harvesting poses a significant threat to the growth and reproduction of mollusks and crustaceans.
5 CONCLUSION
Our seasonal investigation and analysis of results enabled us to reach approximately four major points:
We identified 407 macrobenthic organism species in the gulfweed field off Naozhou Island. Arthropods,mollusks, annelids, and macroalgae were found to be the four dominant benthic macroalgae groups,accounting for 88.7% of total species.
There were significant seasonal variations in macrobenthos species: about 197 species were found in summer, 172 in autumn, 182 in winter, and 146 in spring. A total of 44 species were found in all seasons.The Jaccard similarity index was in the range 0.262-0.422, with the highest similarity index in autumn and spring, and the lowest in spring. Only four species( Sargassum hemiphyllum, Pterocladia capillacea,Tetradita squamosa squamosal, and Clypemorus bifasciatus) were found to be dominant over three seasons, although 15 species were found to be seasonally dominant.
Inhabitant density of the macrobenthos was in the range 809.4 inds./m2to 3 242.0 inds./m2with an average of 1 683.0 inds./m2; biomass was in the range 531.1 g/m2to 1 684.6 g/m2and averaged 1 004.0 g/m2.The species diversity indices ranged from 1.21 to 4.42 with an average of 2.80; the average annual evenness was 0.47; species richness indices varied from 2.97 to 7.24 (average 4.97); and the Simpson dominance index varied from 0.20 to 0.92 with an annual average of 0.75.
Seaweed fields have extremely high primary productivity and a unique food chain. The community structure in our study showed clear seasonal and vertical variation and distribution. The biomass of the benthic organisms, macroalgae and crustaceans displayed significant positive correlation ( P <0.05)with coeffi cients of 0.802, 0.894, and 0.592,respectively. The biomass of macroalgae showed a clear negative correlation with DIN, but the biomass of benthic organisms was somehow negatively correlated with the pH, DIN, phosphate, silicate,chl a, and TOC content of the seawater.
6 DATA AVAILABILITY STATEMENT
The authors declare that all data supporting the findings of this study are available within the article and its supplementary information files.
7 ACKNOWLEDGMENT
We acknowledge the assistance of the Third Institute of Oceanography, Ministry of Natural Resources, in identifying benthic species. We are grateful for sampling and embellishing assistance from our colleagues of Jibiao ZHANG and Guohuan YANG.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Predicting sediment flux from continental shelfislands,southeastern China*
- Laboratory simulation of dissolved oxygen reduction and ammonia nitrogen generation in the decay stage of harmful algae bloom*
- Development of high-resolution chloroplast markers for intraspecific phylogeographic studies of Phaeocystis globosa*
- Effects ofiron and humic acid on competition between Microcystis aeruginosa and Scenedesmus obliquus revealed by HPLC analysis of pigments*
- Effect of river plume on phytoplankton community structure in Zhujiang River estuary*
- Exploring the sublethal genotoxic effects of class II organophosphorus insecticide quinalphos on freshwater fish Cyprinus carpio