利用叶绿素荧光技术分析2 个草莓品种的低温适应性*
2021-04-13杨馥霞王卫成程万莉
杨馥霞,汤 玲,贺 欢,孔 芬,王卫成,程万莉
(1 甘肃省农业科学院林果花卉研究所,兰州 730070)
(2 甘肃省农业科学院旱地农业研究所)
叶绿素荧光技术是一种研究和探测光合生理的植物活体诊断技术,是研究无损叶片光合机构逆境损伤的有效工具,它的变化能反映出几乎所有的光合作用的变化[1]。利用叶绿素荧光技术是深入了解逆境条件下植物光化学过程及叶片功能变化的理想手段,广泛用于逆境胁迫的研究中[2-3]。有关低温胁迫研究表明,温度降至3 ℃,温州蜜柑叶片叶绿素荧光参数Fo上升,Fv/Fm下降,说明此温度下光化学效率受影响[4]。李庆会等[5]通过比较低温胁迫下2 个茶树品种的叶绿素荧光参数,初步判定耐寒性强的品种是‘迎霜’。在对低温下3 种阔叶树的光抑制及恢复研究中发现,非光化学猝灭的光保护机制在越冬树种PSII 免受光抑制中发挥重要作用[6]。
设施栽培的草莓,由于冬季低温,设施保温能力及措施不强,往往受到多种低温伤害,有偶发、极端、短时性的,也有持续性伤害,因此研究低温对草莓的影响在生产中有重要意义。关于低温下草莓叶绿素荧光特性的研究少有报道。针对不同草莓品种间响应低温胁迫、低温稳定性差异等问题,本研究以甘肃省主栽的一季品种‘红颜’和四季品种‘圣安德瑞斯’为试材,利用叶绿素荧光参数的瞬变特征,结合光反应中发生的3 个主要功能步骤(能量吸收、捕获和转换)[7],从快速叶绿素荧光诱导动力学参数角度来揭示2 个草莓品种原初光化学反应对低温胁迫的响应,旨在探讨2 个草莓品种低温下原初光化学反应的差异,同时结合分析相对电导率与叶绿素含量的变化,为进一步揭示草莓应对低温的机制提供依据。
1 材料与方法
1.1 试验材料
供试草莓品种为‘红颜’(Benihoppe)和‘圣安德瑞斯’(San Andreas),其中,‘红颜’为一季品种,‘圣安德瑞斯’为四季品种。
1.2 试验设计
每个品种选择长势一致的匍匐茎苗12 株栽于草炭土为基质的黑色营养钵中,营养钵高12 cm、直径12 cm。培养60 d 后植株长有5~6 片叶进行试验处理。将供试材料置于RXZ 型智能人工气候箱中,在昼25 ℃/夜20 ℃、空气相对湿度75%、PFD为200 μmol·m-2·s-1、光周期12 h 条件下进行培养,定期浇水。7 d 后,测定快速叶绿素荧光动力学参数及叶绿素含量和相对电导率。测定后,其余条件不变,温度降至4 ℃,分别培养1、2、4、6 d 再测定各指标。试验共4 个处理,常温(25 ℃)培养7 d 为对照。
1.3 测定项目与方法
1.3.1 叶绿素含量及相对电导率的测定
取新鲜叶片打孔器打孔后混匀,准确称取0.1 g,置于10 mL 乙醇∶丙酮(1∶1)提取液中浸提,采用UV-2550 紫外可见分光光度计,分别于665、649 nm 波长下测定吸光度值。相对电导率测定参考李俊才等[8]的方法。以上指标测定重复3 次。
1.3.2 快速叶绿素荧光诱导动力学
常温培养7 d 后,于9:00—11:00 测定其中心展开叶往外数第2 片复叶中心小叶的快速叶绿素荧光参数。标记该叶片,分别在4 ℃培养1、2、4、6 d 的9:00—11:00 测定其快速叶绿素荧光参数,重复测定6 次,取平均值。叶片经暗适应30 min 后,暴露于饱和脉冲光(3 000 μmol·m-2·s-1,PFD)下2 s,用连续激发式荧光仪Handy-PEA(Hansatech,UK)绘制快速荧光动力学曲线。
1.4 JIP-test 分析与数据分析
获得的OJIP 荧光诱导曲线进行JIP-test 分析,可得到以下荧光参数:最大光化学效率Fv/Fm=φPo=(Fm-Fo)/Fm;以吸收光能为基础的性能指数PIabs=(RC/ABC)×[φPo/(1-φPo)]×[ψo/(1-ψo)];可变荧光Fv占振幅FJ-Fo的比例Wk=(Fk-Fo)/(FJ-Fo);捕获的激子将电子传递到电子传递链中QA-下游的其他电子受体的概率ψo=1-VJ;反应中心吸收的光能用于电子传递的量子产额φEo=[1-(Fo/Fm)]×ψo;单位面积有活性的反应中心的密度RC/CSo=φPo×(VJ/Mo)×(ABS/CSo);单位反应中心吸收(ABS/RC)、捕获(TRo/RC)、用于电子传递(ETo/RC)及热耗散的能量(DIo/RC)的计算为ABS/RC=Mo×(1/VJ)×(1/φPo),TRo/RC=Mo×(1/VJ),ETo/RC=Mo×(1/VJ)×ψo,DIo/RC=(ABS/RC)-(TRo/RC)。
采用WPS Office 软件整理、统计数据以及作图,采用DPS 数据处理系统进行LSD 多重比较。
2 结果与分析
2.1 低温胁迫对2 个草莓品种快速叶绿素荧光动力学曲线及特殊位点荧光的影响
低温胁迫导致‘红颜’和‘圣安德瑞斯’OJIP曲线发生明显变化,在O 点已略微升高,在J 点处(2 ms)均高于各自的对照,低温胁迫1、2 d I 点(30 ms)的值明显高于对照,在胁迫4、6 d 有所下降,但仍高于对照,且低温胁迫延长了到达最大荧光所需要的时间(图1)。
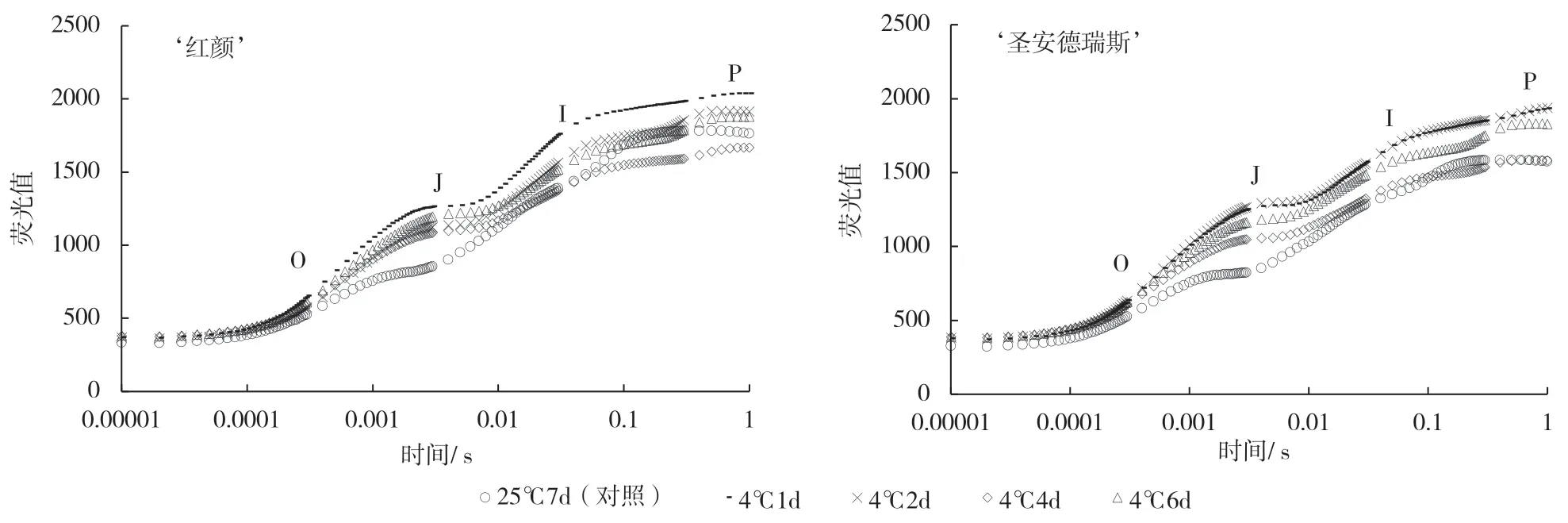
图1 2 个草莓品种快速叶绿素荧光动力学曲线
O 相荧光为初始荧光(Fo),反映反应中心受损伤的程度。2 个草莓品种Fo随着低温胁迫时间的延长而升高。从胁迫1 d 开始,Fo升高,‘红颜’在低温胁迫4 d 开始和对照差异显著;‘圣安德瑞斯’在胁迫1 d 时较对照升高了20.9%,且持续升高,与对照均呈显著差异。2 个品种间相比,‘红颜’在25 ℃处理下(对照),Fo值高于‘圣安德瑞斯’,但在低温胁迫下低于‘圣安德瑞斯’(图2-A)。低温处理后J 相(2 ms)的叶绿素荧光强度2 个品种显著高于各自对照,胁迫导致QB不能及时从QA-接受电子,造成QA-大量积累;低温胁迫下‘红颜’J 相的荧光强度随胁迫时间延长没有显著差异,而‘圣安德瑞斯’在胁迫4 d 时降低,又在胁迫6 d 时上升(图2-B)。I 相(30 ms)的荧光强度在低温胁迫2 d 略有升高,但随胁迫时间继续延长I 相的荧光强度逐渐降低;‘红颜’各处理间差异不显著,‘圣安德瑞斯’各处理间差异较大(图2-C)。P 相是最大荧光产量(Fm),是反应中心处于完全关闭状态时的叶绿素荧光发射强度,Fm值随着低温处理时间的延长呈先升高后降低的趋势,但‘红颜’差异不显著,‘圣安德瑞斯’在低温胁迫4 d 时显著低于胁迫1 d 和2 d(图2-D)。2 个品种的FJ、FI、Fm在初始0~2 d 接近,之后的4~6 d‘圣安德瑞斯’变化较大,暗示‘红颜’光系统II 较‘圣安德瑞斯’稳定。
2.2 低温胁迫对2 个草莓品种PSII 反应中心的影响
随着处理时间的延长,单位反应中心吸收的光能(ABS/RC)、单位反应中心捕获的用于还原QA的能量(TRo/RC)、用于电子传递的能量(ETo/RC)及耗散的热能(DIo/RC)均出现不同程度的变化,通过分析这4 个参数,可以了解当QA处在可还原态时单位反应中心活性的变化情况(表1)。常温下2个草莓品种反应中心各参数的差别不大。低温胁迫使2 个草莓品种单位反应中心吸收的光能(ABS/RC)减少,单位反应中心捕获的用于还原QA的能量(TRo/RC)有所降低,用于电子传递的能量(ETo/RC)也是显著低于各自对照,热耗散的能量(DIo/RC)随低温胁迫时间的延长在6 d 时均高于各自对照,且‘圣安德瑞斯’在低温胁迫4 d 开始高于‘红颜’。
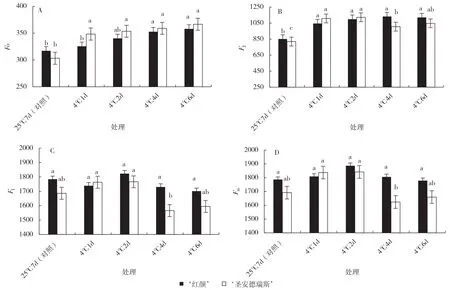
图2 2 个草莓品种特殊位点荧光对低温胁迫的响应
RC/CSo反应的是单位面积内有活性反应中心的数量。结果显示,低温胁迫使2 个草莓品种的RC/CSo与对照相比数量增多。‘红颜’的RC/CSo在低温胁迫下差异不显著,胁迫6 d 时‘红颜’和‘圣安德瑞斯’分别较对照升高了26.2%和24.8%。2 个品种间相比,‘红颜’的RC/CSo增加幅度在低温胁迫1 d 和2 d 时少于‘圣安德瑞斯’,但随着胁迫时间延长变化稳定,‘圣安德瑞斯’则出现较大变动,且‘红颜’反应中心数量增加后吸收的能量多用于推动电子传递(ETo/RC),而‘圣安德瑞斯’热耗散能量(DIo/RC)增加的较多。可见,‘圣安德瑞斯’在低温胁迫后期(6 d),虽然单位反应中心吸收的光能较‘红颜’多,但是没有将吸收的能量更多地用于推动电子传递,而是通过热耗散减轻对植株的伤害。

表1 低温胁迫对2 个草莓品种叶片单位PSII 反应中心活性的影响
2.3 低温胁迫对2 个草莓品种PSII 供体侧的影响
Wk是K 相可变荧光Fk占振幅FJ-FO的比例,通过比较Wk的升高程度可看出放氧复合体被破坏的程度[9]。从图3 可以看出,随着低温胁迫时间的延长,2 个草莓品种的Wk均低于各自对照,结合OJIP曲线中K 相未出现,进一步表明2 个草莓品种叶片PSII 供体侧在4 ℃低温下没有受到伤害。
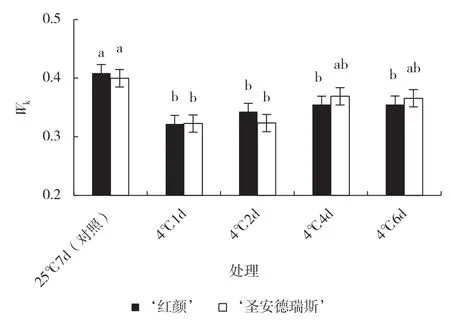
图3 2 个草莓品种叶片K 相可变荧光Fk占振幅FJ-FO的比例
2.4 低温胁迫对2 个草莓品种ψo和φEo的影响
叶片反应中心捕获的激子中,电子传递到QA以下的效率(ψo)随低温胁迫时间的延长而降低,‘红颜’ψo在低温胁迫1 d 时下降了18.6%,胁迫2、4、6 d 时分别下降了20.0%、27.1%、27.4%;而‘圣安德瑞斯’在低温胁迫1 d 时下降了22.0%,胁迫2、4、6 d 时分别下降了23.1%、23.2%、26.1%;与各自对照相比,2 个草莓品种差异均显著(图4)。φEo反映了反应中心吸收的光能用于电子传递的量子产额,值越高表明植物受低温胁迫的影响越小。2 个草莓品种的φEo均降低,低温胁迫1 d 时‘红颜’下降了18.8%,胁迫2、4、6 d 时分别下降了20.3%、28.9%、29.5%;‘圣安德瑞斯’的φEo在低温胁迫1 d 时下降了23.0%,下降幅度较大,胁迫2、4、6 d时分别下降了24.2%、27.5%、29.8%(图4)。从图4 还看出,2 个草莓品种的ψo、φEo在低温胁迫1~2 d 有差异,而之后的6 d 出现趋同。

图4 低温胁迫对2 个草莓品种叶片ψo和φEo的影响
2.5 低温胁迫对2 个草莓品种Fv/Fm和PIabs的影响
Fv/Fm反映了暗反应后最大光化学效率,其值的下降是发生光抑制的重要指标。从图5 可以看出,2 个草莓品种的Fv/Fm随低温胁迫时间的延长整体呈现下降趋势,但在胁迫1 d 和2 d 时与各自对照相比变化不大,胁迫4 d 开始,Fv/Fm与对照差异显著;常温条件下2 个草莓品种叶片的Fv/Fm相近,但低温胁迫条件下‘红颜’叶片的Fv/Fm均高于‘圣安德瑞斯’。可知,低温胁迫开始时‘红颜’和‘圣安德瑞斯’叶片受到轻微光抑制,但在胁迫4 d 开始,2 个草莓品种光抑制均严重,且‘圣安德瑞斯’更严重。

图5 低温胁迫对2 个草莓品种叶片Fv/Fm和PIabs的影响
PIabs是以吸收光能为基础的性能参数,反映低温胁迫对光合机构的影响。2 个草莓品种的PIabs随低温胁迫时间延长而逐渐降低。与各自对照相比,‘红颜’在低温胁迫1、2、4、6 d 的PIabs分别下降了22.6%、31.1%、50.6%、52.7%,‘圣安德瑞斯’分别下降了35.0%、37.4%、51.3%、52.5%。常温条件下2 个草莓品种的PIabs几乎相等;4 ℃胁迫1 d和2 d 时,‘红颜’的PIabs高于‘圣安德瑞斯’,但胁迫4 d 后二者趋同。
2.6 低温胁迫对2 个草莓品种相对电导率和叶绿素含量的影响
低温伤害首先作用于细胞膜上,相对电导率可以反映植株受伤害程度。由图6 可知,常温下,2个草莓品种叶片相对电导率几乎相等,低温胁迫1 d‘圣安德瑞斯’叶片相对电导率高于‘红颜’,但在胁迫2、4、6 d 时略低于‘红颜’。这说明‘圣安德瑞斯’在低温下叶片细胞膜状态没有‘红颜’稳定。
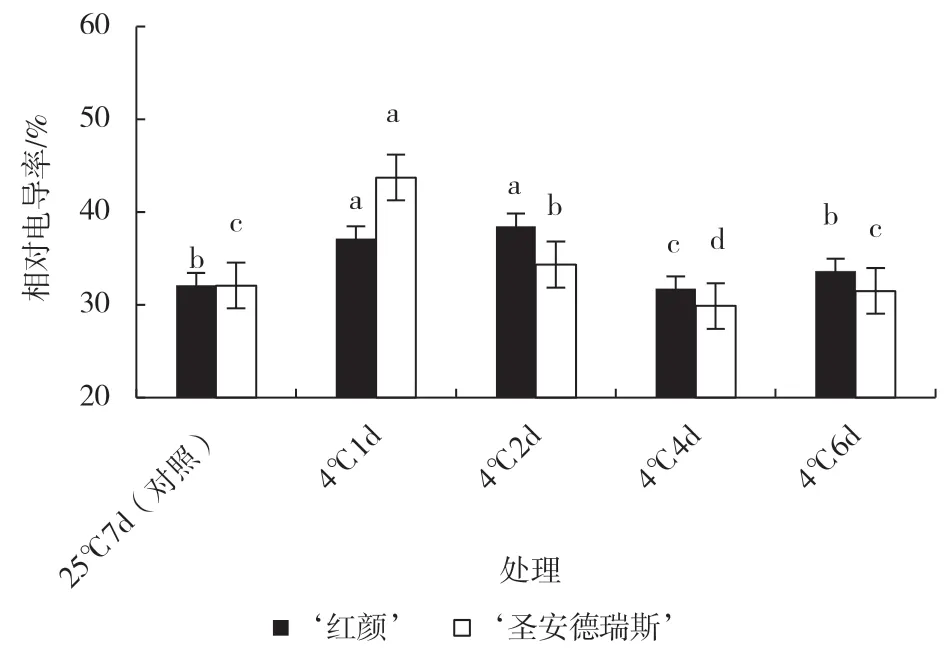
图6 低温胁迫对2 个草莓品种叶片相对电导率的影响
低温延长处理降低了‘红颜’叶片中叶绿素含量(表2),其中在胁迫6 d 时叶绿素a 含量降低了7.9%,叶绿素b 含量降低了26.6%,导致叶绿素a/b值高于对照。‘圣安德瑞斯’在低温胁迫6 d 时叶绿素a 含量降低了4.3%,叶绿素b 含量降低了16.7%,叶绿素含量尤其是叶绿素b 含量的降低,降低了草莓叶片对光的捕获能力。2 个品种相比,常温下叶绿素b 含量‘红颜’高于‘圣安德瑞斯’,在低温胁迫6 d 时二者叶绿素b 含量几乎相等。
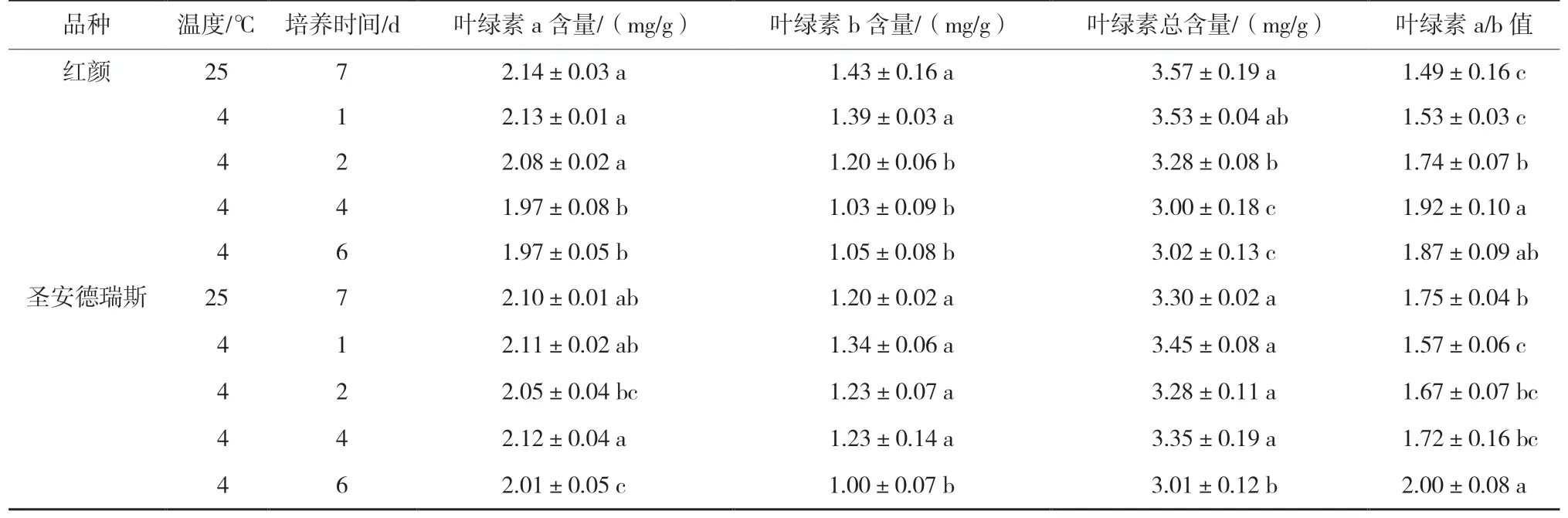
表2 低温胁迫对2 个草莓品种叶片叶绿素含量的影响
3 讨论与结论
通过JIP-test 分析的各参数值可以反映植物光系统Ⅱ对光能的利用情况[10]。逆境中Fo上升,FJ、FI和Fm下降,说明电子在PSⅡ受体侧的传递受到阻碍,QB和PQ 被还原的能力受到限制,电子传递活性降低[11-13],光化学效率(Fv/Fm)下降,表明光合机构被逐渐破坏。本研究表明,随低温胁迫时间延长‘圣安德瑞斯’各参数值变化较大,受到的影响更严重。K 点出现是和供体侧特别是放氧复合体受到伤害有关。从Wk的变化看出,2 个草莓品种叶片PSⅡ反应中心供体侧没有受到明显伤害,这和孙山等[14]在低温弱光胁迫下‘金太阳’杏结果一致,任丽丽等[15]也认为一定条件的低温弱光胁迫不会对放氧复合体造成伤害。但PSⅡ向下游电子传递链传递电子的能力(ψo)不断下降,意味着受到伤害的是PSⅡ受体侧的电子传递链。
大多数情况下,低温导致有活性反应中心的减少[14-15],但本试验中RC/CSo值增大,有活性反应中心单位密度反而增加,Rachid 等[3]发现在干旱胁迫下藜麦也有类似结果,认为有活性的RC转化为钝化的RC,为了适应逆境,通过增加单位面积有活性的反应中心密度来提高光能转化为激发能的效率,这是一种适应低温能力[16]。郑毅[17]认为部分钝化的PSⅡ仍具有启动热耗散保护机制的功能。从孙山等[14]分析中不难发现,PSⅡ反应中心在某些逆境下易形成能量陷阱,能吸收光能但不推动电子传递,本试验很好地印证了这一论断:低温胁迫后期,‘圣安德瑞斯’的RC/CSo减少,单位反应中心吸收的光能(ABS/RC)多于‘红颜’,但并没有将吸收的光能用于推动电子传递,只是较多用于热耗散,形成能量陷阱,反观‘红颜’,虽吸收的光能少,但多用来推动电子传递,低温胁迫下电子传递效率降低是导致耐寒性差异的主要原因[18]。有人认为植物在受到环境威胁时,反应中心利用光能的效率会降低,体内过剩光能对细胞造成损伤,热耗散(DIo/RC)是植物应对氧化胁迫的重要防御机制。但笔者更倾向于高玉录等[19]的分析,认为热耗散的增加是因为PSII 活性受到了抑制。
性能指数是反映光化学反应效率的重要指标,能更好地反映胁迫对光合机构的影响[20]。在对30 个大豆品种抗寒性研究发现,低温下PIabs指数既反应不同大豆品种抗寒能力差异,又显示反应中心吸收光能的能力差异[7]。本试验中2 个草莓品种的光能利用能力(PIabs)支持上述观点,与‘圣安德瑞斯’相比,‘红颜’胁迫下PIabs下降幅度较缓,抗性强。最大光化学效率(Fv/Fm)下降是PSII 发生光抑制的重要指标,但Fv/Fm是有活性和无活性反应中心光化学效率的平均值[21],对环境变化有一定的不敏感性,比起Fv/Fm,PIabs更能敏感地反映叶片光合效率的变化[22]。本试验结果进一步论证,随着低温胁迫时间的延长,PIabs确实比Fv/Fm变化敏感,在胁迫初期光合机构就有所响应。
植物均有低温适应的能力,但不同品种间具有一定的差异。多数学者通过电导率及生理指标的测定分析,综合评价作物在低温环境中的适应性。本试验中利用JIP-test 分析2 个草莓品种各参数值的变化与低温胁迫下电导率及光合色素含量变化分析结果一致:与‘圣安德瑞斯’相比,一季品种‘红颜’在低温状态下具有更强的适应性。但低温胁迫对植物叶绿素荧光特性的抑制作用是环境胁迫和植物响应恢复能力共同作用的结果,有关低温胁迫时间延长后2 个草莓品种恢复能力如何,仍需进一步研究。
