春尺蠖细胞色素C氧化酶亚基I基因的克隆与系统进化分析*
2021-04-12陈龙崔阔澍单艳敏查干
陈龙 崔阔澍 单艳敏 查干
1.河套学院农学系,内蒙古巴彦淖尔
2.四川省农业技术推广总站,四川成都
3.内蒙古自治区草原工作站,内蒙古巴彦淖尔
4.巴彦淖尔市草原工作站,内蒙古巴彦淖尔
春尺蠖Apocheima cinerarius Ershoff,又称春尺蛾,是一种鳞翅目、尺蛾科杂食性害虫,被我国多个省、自治区列为重点监测范围,具有繁殖成活率高、蛹羽化速度快、多食性、暴食性等特点[1-2]。近年来,春尺蠖主要危害范围呈扩大趋势,危害对象从桑、梨等20余种林木逐步发展到玉米、小麦等农作物,危害部位多为叶片,危害区域从内蒙古、新疆、黄淮海等北方区域逐步向四川、云南等南方区域爆发迁移。本研究对春尺蠖遗传规律进行了分析,进一步探明了春尺蠖在昆虫纲Insecta进化树中的位置及其进化规律,不仅具有分子遗传学价值,而且具有预期防治意义。
细胞色素C氧化酶亚基Ⅰ(COⅠ)是线粒体DNA(mtDNA)中最保守的蛋白,是动物系统发育关系分析的条形码[3],其序列已经被广泛应用于膜翅目昆虫蜜蜂[4]、鳞翅目[5,6]、双翅目果蝇[7]等昆虫种间的分子鉴定和系统发育研究。目前研究表明,线粒体遗传密码与基因组遗传密码相比较,不仅具有高度的相似性,且更能代表遗传密码的早期形式[8],因此,利用COⅠ基因作为分子标记来分析种群的遗传结构和变异,可从分子学角度为春尺蠖的鉴定分类和系统进化提供可靠的依据[9]。虽然Liu等[10]在NCBI中已上传春尺蠖线粒体基因组序列并分析了其全基因序列,但还未见关于COⅠ基因的序列分析及系统进化关系相关的报道。本研究克隆获取了春尺蠖COⅠ基因序列,并对春尺蠖线粒体基因组COⅠ基因进行了序列分析,明确了其分子遗传特征,对其蛋白的理化性质进行了分析,预测了其编码蛋白的分子结构,提出了春尺蠖与近源主要类群间的系统发育关系,为明确春尺蠖在鳞翅目昆虫的分类地位及分子系统进化关系,以及春尺蠖种群分布与进化关系的深入研究奠定了基础。
1 材料与方法
1.1 材料
供试昆虫:内蒙古巴彦淖尔市乌拉特前旗柠条草场(E:108°45′ 23.63″ ; N: 40°46′ 4.19″ )的春尺蠖雌雄成虫。 主要试剂: TaKaRa MiniBEST Universal Genomic DNA Extraction Kit Ver.5.0、TaKaRa MiniBEST Agarose Gel DNA Extraction Kit Ver.4.0、连接载体pMD19-T Vector Cloning Kit、2×PCR Master Mix 及 TaKaRa Taq 购自大连 TaKaRa 公司;DH5α 感受态细胞购自北京天根(Tiangen)公司。
1.2 DNA 提取
将春尺蠖雌雄成虫各取3 只混合后,置于灭菌后的研钵中用液氮研磨,具体提取步骤参照TaKaRa DNA 提取试剂盒说明书。而后利用1.0%的琼脂糖凝胶电泳与Nano Photometer™P-Class 超微量分光光度计分别检测提取DNA 的质量和浓度。
1.3 COⅠ基因序列克隆
由于COⅠ基因内部含AT 序列富集区,所以利用Primer Premier 5.0 软件在COⅠ基因外围设计RT-PCR扩增引物的方法来扩增春尺蠖COⅠ基因的全序列。上游引物 COⅠ-F: 5′ -ATAAAAGAGAAATATTCCC GTAAAT-3′;下游引物 COⅠ-R: 5′ -ATAGTCTGC CATATTAGAAGTTTCT-3′。PCR 反应体系共 25 μL(扩增模板1 μL,上下游引物各1 μL,Master Mix 12.5 μL,RNase-free Water 9.5 μL)。PCR 反应条件:预变性 94 ℃、5 min;94 ℃、30 s,56 ℃、30 s,72 ℃、1 min 30 次循环;72 ℃延伸10 min,4 ℃保存。利用1%浓度的琼脂糖凝胶电泳检测PCR 产物,而后经回收、亚克隆送北京六合华大公司测序。
1.4 春尺蠖COⅠ基因的生物信息学分析
利用在线预测网站ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)预测基因蛋白编码序列;使用ExPASy - ProtParam tool (http://web.expasy.org /protparam/)预测春尺蠖COⅠ编码氨基酸的基本理化性质,利用DNAMAN V6.0(Lynnon Biosoft,加拿大)分析COⅠ基因编码氨基酸的同源性和遗传距离;应用SignalIP5.0(http://www.cbs.dtu.dk/services/SignalP/) 预测COⅠ基因编码的氨基酸N 端信号肽;用TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)预测蛋白跨膜区域;用SWISS-MODEL 预测蛋白质三级结构;用MEGA6.0 中的邻接法(neighbor-joining,NJ)构建系统进化树,采用p距离(P-distance) 构建系统发育树。
2 结果与分析
2.1 春尺蠖COⅠ基因的克隆
根据NCBI 中已上传的春尺蠖CO Ⅰ基因序列信息,以COⅠ-F 和COⅠ-R为引物在目的基因外围扩增1 621 bp 的片段,经1%浓度的琼脂糖凝胶电泳检测发现有一条1 600 bp 左右的特异性条带,测序后发现其包含的1 531 bp 序列与春尺蠖线粒体基因组数据库中该基因的序列比对一致。
2.2 春尺蠖COⅠ蛋白的理化性质
春尺蠖COⅠ基因全长1 531 bp,编码蛋白区以CGA 作为起始,以碱基T 作为不完全终止,其中,A、T、 C、 G 的含量分别为 32.6%、 39.2%、 14.0%、14.2%,C+G 含量为28.2%,A+T 含量为71.8%,符合动物线粒体基因序列特征。
在COⅠ蛋白的理化性质分析结果中显示,该蛋白的预测分子式为C2660H4021N623O685S29,共包含510 个氨基酸,蛋白质分子量为56.62 kD,等电点为5.92;半衰期和脂肪指数分别为1 h、113.96,负电荷残基(Asp + Glu)总电荷的24 高于正电荷残基(Arg+ Lys)总电荷的15;该蛋白的亲水性为0.705,为疏水性蛋白。不稳定系数为34.62,小于参考值40.00,故COⅠ为稳定蛋白。SignalP 5.0 Server 预测结果显示,COⅠ在N端不含信号肽(信号肽预测见图1);通过预测该蛋白的跨膜区和三维结构,结果显示,COⅠ蛋白含12个跨膜区(跨膜结构预测见图2),而三维结构模型(图3)中绝大部分为α 螺旋,数量与图2 跨膜区一致。
2.3 春尺蠖COⅠ基因的同源性比对及系统进化关系分析
将春尺蠖COⅠ基因的氨基酸序列与NCBI 中搜索得到的其它昆虫序列进行比对,结果表明春尺蠖COⅠ基因氨基酸序列与其它昆虫的序列有很高的一致性,均高于93.00%。与同为鳞翅目尺蠖蛾科的茶尺蠖COⅠ和同为鳞翅目的草地贪夜蛾COⅠ氨基酸序列一致性最高(95.69%),其次为草地螟COⅠ和菜粉蝶COⅠ,一致性分别为95.29%和93.14%。
图1 春尺蠖CO I基因编码蛋白信号肽预测图
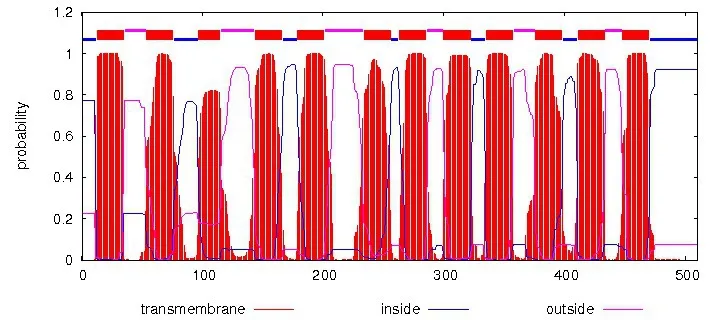
图2 春尺蠖CO I基因编码蛋白跨膜结构预测
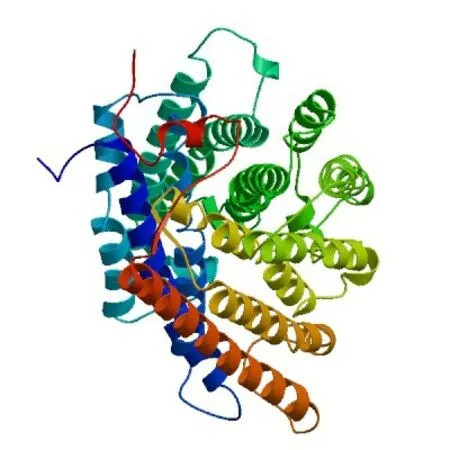
图3 春尺蠖CO I蛋白三维结构模型
通过NCBI 搜索其他目昆虫已上传的COⅠ的氨基酸序列与春尺蠖COⅠ序列信息结合构建系统进化树。结果表明,进化树中的鳞翅目、鞘翅目、双翅目和膜翅目分别聚在不同分支上;在鳞翅目分支上春尺蠖COⅠ与同为鳞翅目尺蠖蛾科的茶尺蠖COⅠ聚为一类,二者亲缘关系最近,遗传距离为0.022(表1);与鞘翅目、双翅目和膜翅目昆虫的亲缘关系依次变远,遗传距离范围在0.042~0.080 之间。这表明COⅠ基因在鳞翅目和其它目昆虫中具有相对保守的进化关系,与诸多有关鳞翅目DNA条形码研究的结论一致。
3 讨论
3.1 春尺蠖COⅠ基因信息特征分析
虽然,鳞翅目昆虫种类超过18 万种,遗传规律复杂多变,但有研究表明鳞翅目昆虫mtDNA 具有显著的碱基偏倚性。如Liao 等[11]在对鳞翅目多种昆虫的研究中发现,平均A+T 含量在77.0%以上,在基因编码控制区可达90.0%以上。杨兆富[12]等对草螟科野螟亚科的总COⅠ基因研究中发现,野螟亚科COⅠ基因A+T 含量为70.0%。呼晓庆[13]等对鳞翅目昆虫中国草地螟的研究中发现,中国草地螟COⅠ基因长度为658 bp,A+T含量为70.6%。除鳞翅目昆虫外,在鞘翅目昆虫茄十二星瓢虫和赤拟谷盗的COⅠ基因中也有相同的特征[14]。 本 研 究 克 隆 获 得 的 春 尺 蠖COⅠ基因全长为1 531 bp,其中,A、T、C、G的含量分别为32.6%、39.2%、14.0%、14.2%,A+T 含量为71.8%,远高于C+G 的含量28.2%,符合昆虫mtDNA蛋白编码序列A+T 含量都较高的典型特征[15]。根据NCBI已有春尺蠖COⅠ基因信息,通过RT-PCR技术克隆获得测序后发现它包含春尺蠖COⅠ基因,经与春尺蠖线粒体基因组数据库中该基因的序列比对,发现两者序列一致,结合该基因的生物信息学、同源基因序列比对和后续系统发育树等分析,进一步确认该基因是COⅠ基因。
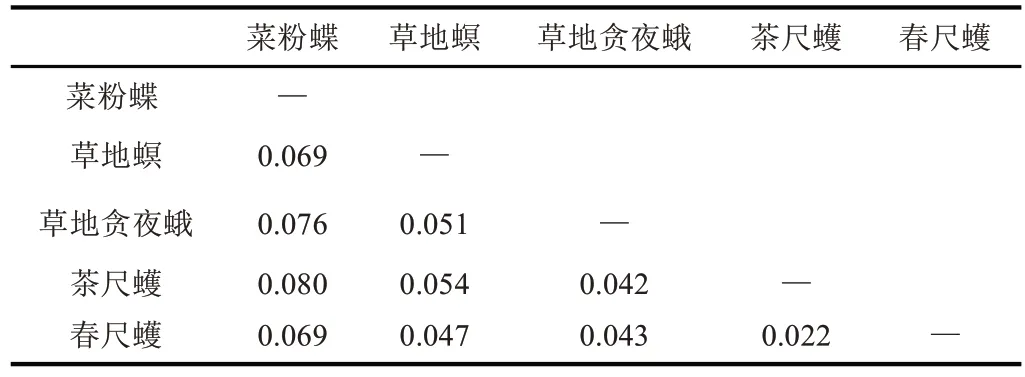
表1 鳞翅目昆虫COⅠ基因遗传距离
3.2 春尺蠖COⅠ基因与其近缘种起始密码子、终止密码子序列的比较
mtDNA 与细胞核基因相比,其基因结构简单,共编码13种蛋白,与其他编码蛋白相比,COⅠ基因进化缓慢,具有较高的保守性,其起始密码子与终止密码子也较为特殊,蛋白编码系统会因生物不同的进化而产生差异,这种差异同样出现于细胞核基因和细胞器内[16]。目前,COⅠ基因还未发现通用的起始密码子,在已报道的5 种鳞翅目昆虫线粒体基因中,除茶尺蠖(起始密码子为ATG)外,其余4 种昆虫5’端高度同源,均以CGA 作为起始密码子[17]。研究发现,与春尺蠖亲缘关系较近的家蚕[18]、稻纵卷叶螟[19]、米蛾[20]、竹纹枯叶蛾[21]、青黄枯叶蛾[22]和文山松毛虫23]的COⅠ基因起始密码子也均为CGA,这可以作为起始密码子判断的一个重要依据。本研究对春尺蠖COⅠ基因进行测序,发现其编码蛋白区同样以CGA 作为起始,这也验证了鳞翅目昆虫线粒体基因COⅠ基因进化缓慢,具有高度保守性这一特性,填补了国内对春尺蠖COⅠ基因研究的空白。但即便众多鳞翅目昆虫的起始密码子均为CGA[24],在多序列比对的研究中,也有学者发现,线粒体基因起始密码子是ATN、GTG 与TTG 等,这表明COⅠ基因虽为mtDNA 中最保守的基因,其起始密码子序列却具有复杂性及多样性,并非严格的保守单一序列,造成这种现象的原因是什么,CGA 是否为真正的密码子值得进一步深入研究。
长期以来,无脊椎动物线粒体基因终止密码存在一定争议性,通常无脊椎动物线粒体基因存在两种形式的终止密码子,即TAR 和以碱基T 的不完全终止。部分研究学者提出应遵循一些识别基本原则来对复杂的终止密码子进行多项条件比对后确定。目前,一些研究发现线粒体基因组在高级阶元系统发育关系研究中存在碱基替换饱和、长枝吸引、类长枝效应、位点速率异质性和分组策略选择等问题[25-27],在本研究序列分析中发现,春尺蠖COⅠ基因以碱基T的不完全作为翻译终止,在小蜡螟、大蜡螟与米蛾的基因序列中也有相同的现象[28]。这源于mtDNA 功能与结构的简约,这种现象在后生动物mtDNA 中普遍存在,这种基因的3’端终止密码子不完整,一般3’端都与tRNA紧密相连,完整的终止密码子由其mRNA 转录后形成的 poly(A)结构来补充完整[29,30]。
3.3 春尺蠖COⅠ基因保守性与亲缘分析
上世纪末,线粒体COⅠ基因首先被应用于昆虫纲的系统发育研究,此后,陆续被应用于不同阶元类群的昆虫系统发育分析。已有学者对鳞翅目昆虫及其亲缘关系较近的双翅目、鞘翅目、膜翅目昆虫COⅠ基因进行了系统进化分析并取得了许多创新性突破。如张永红[28]等首次预测了小蜡螟COⅠ基因编码信息,并与其他3种螟蛾科昆虫COⅠ基因编码区进行了多序列比对,研究发现其起始密码子为CGA,与其遗传距离较近的大腊螟COⅠ基因起始密码子TAA 有显著不同,这一发现体现了线粒体基因的多态性和异质性;史妍茹[31]等首次测定了螟蛾总科米蛾及巢蛾总科旋纹潜叶蛾线粒体全基因组序列,并分别进行了注释,预测了米蛾及旋纹潜叶蛾tRNA 为典型的三叶草结构,通过分子生物学验证了该科昆虫tRNA 的理论结构。本研究通过序列同源性分析,发现春尺蠖COⅠ基因的氨基酸序列与其他近源昆虫相似度高达93.0%以上,有较高的同源性和一致性,进一步验证了春尺蠖COⅠ基因与其他线粒体基因相比具有更好的保守性[32]。在本研究对系统发育树的研究结果中也发现了相似的结果,表明在COⅠ整个系统进化过程中,春尺蠖与同为尺蛾科的茶尺蠖亲缘关系最近,和同为鳞翅目的昆虫草地贪夜蛾亲缘关系较近,与来自不同目的昆虫COⅠ基因亲缘关系差异也较为明显,其中,鳞翅目与鞘翅目亲缘关系最近,其次为双翅目,与膜翅目的亲缘关系最远,这与其他昆虫COⅠ基因的系统进化研究中得出的结果类似。由此说明,COⅠ具有保守性的同时还具有差异性,从而使得它能在不同物种和不同的生活环境中不断提高适应性,起到独特的生物学功能。
虽然本文对春尺蠖COⅠ基因的序列及进化关系进行了全面分析,但是还有一些问题有待进一步明确。首先,线粒体基因的遗传信息是典型的母性遗传,它与细胞核基因相比基因片段与进化程度在分析近缘及种内群体的进化关系上更具有先天的优势,但任何单一基因标记都具有优势和缺陷,种间的遗传进化分析都需要结合多种分子标记去研究,与COⅠ基因相比,线粒体基因组还有其他保守的基因序列(如COⅡ和CYTB 等),将它们用于进化分析,所得结果是否相同。其次,细胞核基因在不同组织、雌雄虫体和不同龄期中的表达是不同的,作为细胞质基因的COⅠ基因是否同它们一样,有组织、性别及龄期的表达异特性。第三,在草地贪夜蛾鉴定分子标记鉴定中,常结合核基因组Z 染色体上的磷酸甘油醛异构酶基因(Tpi)作为分类措施,在春尺蠖分子鉴定中是否需要其他基因协同鉴定以区分不同亚型。以上问题均需要进一步研究。
4 结论
本研究克隆了春尺蠖COⅠ基因,明确了春尺蠖COⅠ基因氨基酸的理化性质及分子系统进化关系,并预测了其蛋白质结构模型,为研究春尺蠖的起源与演化提供理论基础。结果表明,春尺蠖COⅠ基因的蛋白编码区以CGA 为起始密码子,以碱基T 作为不完全终止,基因全长为1 531 bp,编码区包含510 个氨基酸,其分子量与等电点分别为56.62 kD 和5.92;N 端未发现信号肽,具有12 个跨膜结构;预测分子式为C2660H4021N623O685S29,不稳定系数为34.62,总平均亲水系数为0.705,为疏水性的稳定蛋白。遗传距离结果发现,春尺蠖与茶尺蠖遗传距离最近(为0.022)。同源序列比对和系统进化分析表明,春尺蠖与茶尺蠖同源性最高,氨基酸一致性为95.7%。
