毛果杨MAP65 基因家族的扩张与表达分析
2021-04-10刘晓霞吕中睿张建国饶国栋
薛 丽,刘晓霞,罗 莹,吕中睿,张建国,2,3,饶国栋,2,3*
(1. 林木遗传育种国家重点实验室,中国林业科学研究院林业研究所,北京 100091;2. 南京林业大学,南方现代林业协同创新中心,江苏 南京 210000;3. 国家林业和草原局林木培育重点实验室,中国林业科学研究院林业研究所,北京 100091)
微管是由α、β-微管蛋白以异二聚体的形式组装而成的中空管状结构[1]。植物细胞中,微管可以快速的聚合或者解聚、迅速转变微管列阵,对细胞的正常扩张和分化具有重要作用。微管结合蛋白(MAP)是一类对微管的组装和功能具有调控作用的蛋白[2]。Changjie 等最早从烟草(Nicotiana tabacumL.) BY-2 (‘Bright Yellow-2’)悬浮培养细胞中分离得到一类与微管紧密结合且分子量约为65 kD 的蛋白,即MAP65[3]。MAP65 蛋白通常定位于一种或几种微管列阵上,是一类可与微管交联并促进微管成束的微管结合蛋白[4]。
从烟草BY-2 悬浮培养细胞中分离出小原生质体,制备胞质提取物,之后加入紫杉醇协助组装微管,再将微管和MAP 低温高速离心以去组装,经过数次组装-去组装循环,分离得到3 个烟草MAP65蛋白[5]。后用同样的方法在胡萝卜(Daucus carotaL.)悬浮细胞中分离得到3 个胡萝卜MAP65 蛋白[6]。随着拟南芥(Arabidopsis thaliana(L.) Heynh.)与水稻(Oryza sativaL.)基因组测序的完成,拟南芥和水稻中分别鉴定出了9 和11 个MAP65 蛋白家族成员[7-8]。小立碗藓 (Physcomitrella patens(Hedw.)Bruch & Schimp.)中克隆得到了5 个MAP65基因[9]。对拟南芥MAP65 蛋白家族的研究表明,9 个成员之间氨基酸相似性为28%~79%,C 端序列同源性尤其低,C 端包含与微管特定结合的结构域,决定了微管聚合的速率,不同基因的C 端微管结合结构域不同,故以不同的效率促进微管聚合[10]。ATMAP65-1和ATMAP65-2促进扩增细胞的轴向生长,但不是维持细胞营养生长和生殖生长所必需[11]。ATMAP65-3的缺失导致成膜体异常,胞质分裂不完全及植株发育异常[12]。ATMAP65-4和ATMAP65-3具有高度同源性,功能相似,交联成膜体中的相邻反向平行的微管,这2 个基因的同时缺失会导致植株生长受到严重影响[13]。ATMAP65-5在整个细胞周期中均有表达,且对微管解聚药具有更强的耐受力[14]。ATMAP65-6对微管聚合没有促进作用,但可诱导微管形成网状结构,该蛋白定位于线粒体,可能参与线粒体相关功能的实现[15]。植物中MAP65基因家族的成员数量较多,且在细胞周期的不同微管列阵中具有不完全相同的定位模式,在不同组织中的表达量亦存在差异,这预示该基因家族在植物中具有功能的分化[10,16-17]。
毛果杨(Populus trichocarpaTorr. & Gray)作为第一个完成基因组测序的木本植物,为鉴定木本植物MAP65基因家族成员和分析其进化及功能提供了可能[18]。本研究在毛果杨中鉴定出9 个MAP65,通过其理化性质分析、亚细胞定位的预测、系统进化分析、基因结构和保守结构域分析、共线性分析及同义突变频率 (Ks)、非同义突变频率(Ka)分析、组织表达量以及启动子顺式作用元件分析,探究了PtMAP65基因家族的进化扩张模式,以期揭示PtMAP65基因家族的进化生物学意义,并为进一步研究PtMAP65基因家族成员的潜在功能提供新的线索。
1 材料与方法
1.1 PtMAP65 基因家族成员的鉴定及理化性质分析
在拟南芥数据库(https://www.arabidopsis.org)下载ATMAP65基因家族9 个成员的蛋白质序列,将其作为靶序列,使用BLASTP 方法在Phytozome(v12.1,https://phytozome.jgi.doe.gov/) 中的毛果杨v3.1 蛋白质组数据库中进行搜索,期望阈值设为-10,其他参数默认;候选基因再通过在线工具SMART(http://smart.embl-heidelberg.de/)和NCBICDD (https://www.ncbi.nlm.nih.gov)进行保守结构域鉴定,最后得到9 个PtMAP65 蛋白。利用Prot-Param(https://web.expasy.org/)计算PtMAP65 蛋白家族的分子量和等电点;利用Plant-mPLoc(http://www.csbio.sjtu.edu.cn/)、 LocTree3 (https://rostlab.org/)和 Wolfpsort (https://www.genscript.com/)预测PtMAP65 蛋白家族的亚细胞定位。
1.2 植物MAP65-Likes 基因系统进化树的构建
利用MAGA7.0 中的ClustalW 程序,将毛果杨、拟南芥、烟草、水稻、玉米(Zea maysL.)、巨桉(Eucalyptus grandisHill)、挪威云杉(Picea abies(L.) Karst.)、北美云杉(Picea sitchensis(Bongard)Carrière)、莱 茵 衣 藻(Chlamydomonas reinhardtiiP.A.Dangeard)等物种的MAP65-Like 蛋白序列进行多重序列比对;以莱茵衣藻作为外类群,采用邻接法(NJ)构建系统进化树,模型为p-distance,成对删除,校验参数(Bootstrap) 为1 000 次和5 000 次重复。毛果杨、拟南芥、烟草和水稻MAP65 蛋白家族系统进化树构建方法同上。
1.3 毛果杨、拟南芥、烟草和水稻MAP65 家族成员的基因结构和保守结构域分析
利用GSDS (2.0, http://gsds.cbi.pku.edu.cn/)分析毛果杨、拟南芥、烟草和水稻MAP65s的基因结构;利用MEME (v5.1.1, http://meme-suite.org/)分析其蛋白序列的保守结构域,最大结构域数目设置为15。
1.4 毛果杨染色体定位、共线性分析及Ka/Ks 计算
从毛果杨基因组注释文件中获得PtMAP65基因家族染色体定位的位置信息;在linux 系统下利用MCScanX 进行毛果杨基因组内共线性分析,并使用TBtools (v0.6696)作图。利用BioEdit Sequence Alignment Editor 比对复制基因对的CDS 序列,再用DnaSP5 计算复制基因对的Ks和Ka,根据Ks值计算进化时间(T):T=Ks/2λ,λ = 9.1 × 10-9[19]。
1.5 PtMAP65 基因家族成员启动子顺式作用元件分析
使用TBtools (v0.6696)从毛果杨基因组序列文件中获取PtMAP65基因家族成员起始密码子上游2 000 bp 的DNA 序列作为启动子,使用plantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析启动子中的顺式作用元件。
1.6 PtMAP65 基因家族成员表达特性
PtMAP65基因家族成员在不同组织中的表达数据来源于Poplar eFP Browser (http://bar.utoronto.ca/),利用TBtools (v0.6696)分析制作基因表达热图,取log scal,极限值设置为2 000,其他参数默认。
2 结果与分析
2.1 PtMAP65 基因家族成员的鉴定与理化性质分析
通过在线BLASTP 比对,从毛果杨中鉴定出14 个候选的MAP65s。SMART 和NCBI-CDD 分析结果显示:14 条蛋白序列都具有完整或不完整的MAP65_ASE1 结构域 (PF03999);经过比对建树分析,删除影响树形的序列,最后得到9 个MAP65s。根据其在染色体上的分布情况,将其命名为PtMAP65-1到PtMAP65-9(表1)。PtMAP65 蛋白家族成员的氨基酸长度变化范围为570~730 aa,分子量变化范围为64.65~83.00 kDa,等电点变化范围为5.17~8.45。综合考虑PtMAP65 蛋白家族成员用3 种方法预测的亚细胞定位,最终预测:PtMAP65-1、PtMAP65-3 和PtMAP65-9 位于细胞核,PtMAP65-8位于叶绿体,其余成员位于细胞质。
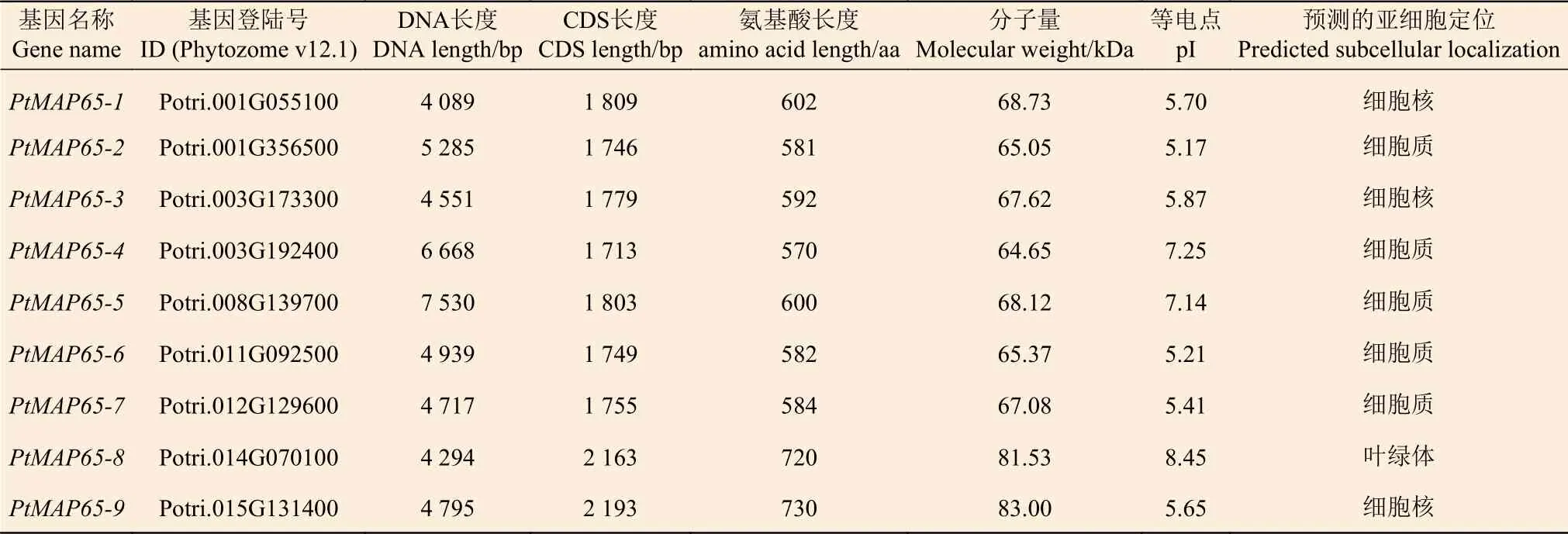
表1 PtMAP65 基因家族及其理化性质Table 1 The information of the PtMAP65 gene family
2.2 植物MAP65-Likes 系统进化分析
为研究植物MAP65-Likes 的系统进化关系,采用邻接法对毛果杨、拟南芥、烟草、水稻、巨桉、挪威云杉等物种的MAP65-Like 蛋白构建系统进化树,校验参数(Bootstrap) 分别为1 000 次和5 000 次重复,结果差异很小,图1 中系统进化树的Bootstrap 为1 000。从图1 可以看出:该植物基因家族分为Ⅰ、Ⅱ、Ⅲ、Ⅳ和Ⅴ5 个亚组,裸子植物的MAP65-Likes仅分布于Ⅰ亚组和Ⅴ亚组中,且独立地聚为一簇;被子植物的MAP65-Likes在5 个亚组中均有分布,且单子叶和双子叶植物MAP65s均匀地分布于各个亚组,这说明5 个亚组的分化发生于裸子植物和被子植物分化之后、单子叶植物和双子叶植物分化之前。在Ⅰ、Ⅱ、Ⅲ和Ⅳ4 个亚组中,单子叶植物和双子叶植物的MAP65-Likes各自聚为一簇,在Ⅴ亚组中,双子叶植物毛果杨、拟南芥和烟草的4 个MAP65-Likes聚为一簇,其他8 个MAP65-Likes与单子叶植物水稻的3 个MAP65s聚为一簇。在Ⅰ亚组中,PtMAP65-2/PtMAP65-6组成旁系同源基因对,并与ATMAP65-1/ATMAP65-2这一对旁系同源基因聚为一支;在Ⅱ亚组中,PtMAP65-4/Eucgr.J00083.1组成直系同源基因对,并与ATMAP65-5聚为一支;在Ⅲ亚组中,PtMAP65-1/PtMAP65-3组成旁系同源基因对,并与Eucgr.B03489.1聚为一支;在Ⅴ亚组中,PtMAP65-7/PtMAP65-9组成旁系同源基因对,并与旁系同源基因对Eucgr.K00145.1/Eucgr.A02875.1聚为一支,另外,PtMAP65-8/ATMAP65-4组成直系同源基因对;在Ⅳ亚组中,PtMAP65-5/Eucgr.G02071.1组成直系同源基因对,这表明MAP65 家族中毛果杨与巨桉的亲缘关系最近,这也与二者是多年生木本植物相一致。
2.3 毛果杨、拟南芥、烟草和水稻MAP65 家族成员的基因结构和保守结构域分析
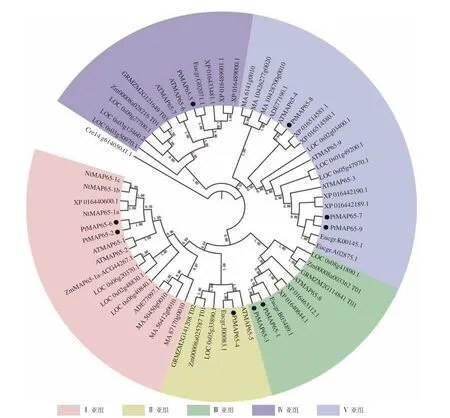
图1 植物MAP65-Likes 系统进化分析Fig. 1 Phylogenetic analysis of plant MAP65-Likes protein sequences
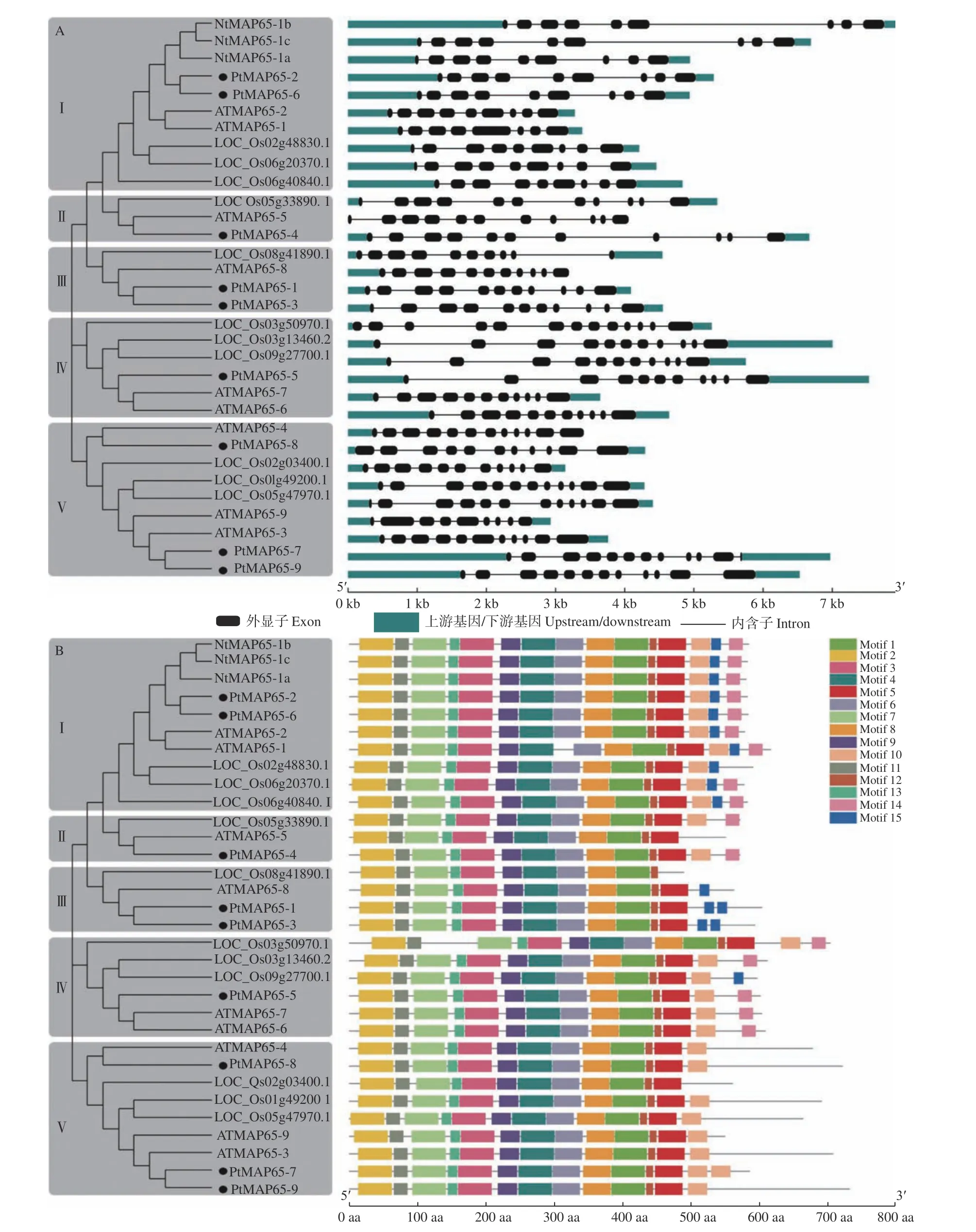
图2 MAP65 家族的基因结构和保守结构域分析Fig. 2 Gene structures and protein domains analysis of MAP65 gene family
为分析MAP65家族成员的基因结构特征,利用GSDS 绘制了毛果杨、拟南芥、烟草和水稻MAP65家族成员的基因结构图。图2A 表明:在Ⅰ亚组中,9 个MAP65s有9 个外显子、8 个内含子,仅ATMAP65-1有8 个外显子、7 个内含子;在Ⅱ亚组中,MAP65有11 个外显子、10 个内含子;在Ⅲ亚组中,3 个MAP65有11 个外显子、10 个 内 含子,LOC_Os08g41890.1有10 个 外 显子、9 个内含子;在Ⅳ亚组中,5 个MAP65有11 个外显子、10 个内含子,仅LOC_Os03g50970.1有13 个外显子、12 个内含子;在Ⅴ亚组中,MAP65的外显子个数为9~12、内含子个数为8~11。另外,在各个亚组中,外显子的长度相似,内含子的长度差异较大,而同一物种的MAP65之间的内含子长度更相似。
利用MEME 分析了MAP65 蛋白家族的保守结构域。图2B 表明:各个MAP65 蛋白家族成员具有11~15 个不等的结构域,第1、2、3、4、6、7、8、9、11、12 和15 个结构域较保守,在32 条MAP65 蛋白序列中均有分布。另外,植物MAP65 蛋白序列的C 端结构域较多变,各个亚组的MAP65 蛋白具有相似的结构域分布。
2.4 PtMAP65 基因家族的染色体定位和进化扩张
根据毛果杨基因组序列文件和注释文件,分析了9 个PtMAP65基因在染色体的定位情况。PtMAP65-1和PtMAP65-2位于Chr01,PtMAP65-3和PtMAP65-4位于Chr03,PtMAP65-5到PtMAP65-9分别位于Chr08、Chr11、Chr12、Chr14 和Chr15。毛果杨基因组内共线性分析结果显示:MAP65基因家族中共有6 对复制基因,且均为片段复制,其中PtMAP65-7、PtMAP65-8和PtMAP65-9互为复制基因(图3)。利用DnaSP5 计算复制基因对的Ka和Ks,算得的Ks值为0.234 1~1.933 8,表明复制时间最早发生于106.25 百万年前,最晚发生于12.86 百万年前;除复制基因对PtMAP65-4/Potri.001G032566的Ka/Ks 为0.617 3 外,其余复制基因对的Ka/Ks< 0.25,表明PtMAP65基因家族在进化过程中受到强纯化选择的作用(表2)。
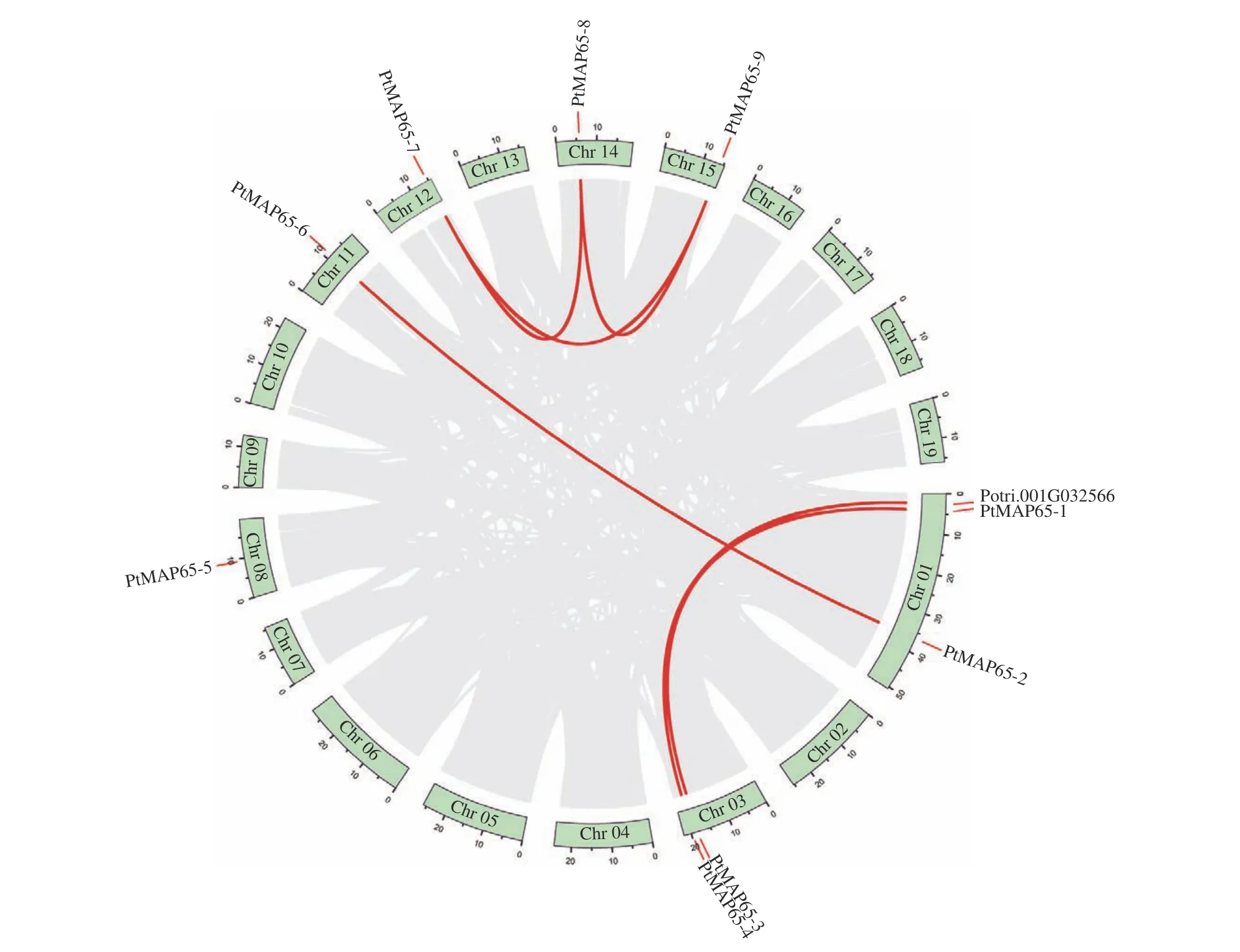
图3 PtMAP65 基因家族染色体分布与复制基因对Fig. 3 Chromosomal locations and duplicated gene pairs of PtMAP65 gene family
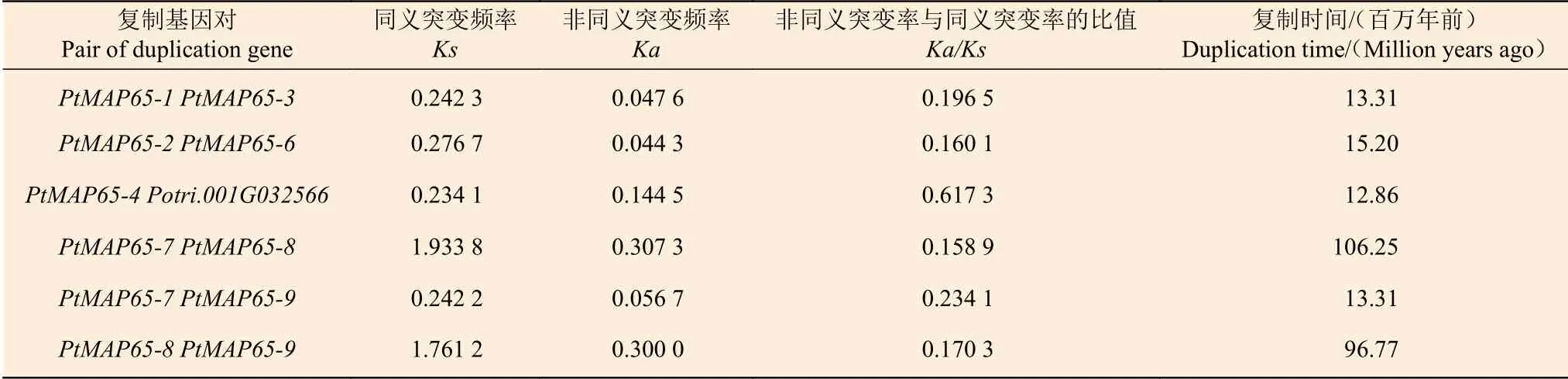
表2 复制基因对的Ks、Ka 分析Table 2 Ks and Ka analysis of duplicated gene pairs
2.5 PtMAP65 基因家族的启动子顺式作用元件分析
为了解转录调控机制,利用PlantCARE 数据库分析PtMAP65基因家族启动子的顺式作用元件。根据顺式作用元件的功能,将其分为光响应元件、激素响应元件、逆境胁迫响应元件和植物生长发育元件。图4 表明:在PtMAP65基因家族中,光响应元件和激素响应元件的个数较多,逆境胁迫响应元件和植物生长发育元件的个数较少;PtMAP65-3中的激素响应元件远多于其他基因,PtMAP65-4、PtMAP65-7和PtMAP65-8中未预测到植物生长发育元件。
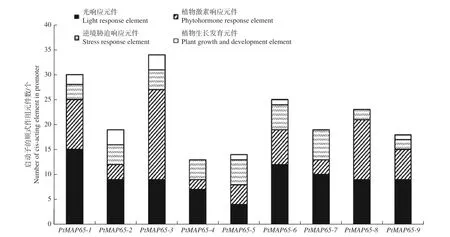
图4 PtMAP65 基因家族启动子的顺式作用元件分析Fig. 4 Cis-elements analysis of PtMAP65 gene family promoter
2.6 PtMAP65 基因家族的组织表达模式分析
从Populus eFP browser 数据库下载得到PtMAP65基因家族成员在成熟叶、幼叶、根、雌花、雄花和木质部中的表达数据,绘制热图。图5表明:复制基因对PtMAP65-2/PtMAP65-6在幼叶、根和木质部均有高度表达;复制基因对PtMAP65-7/PtMAP65-8在雌花和雄花中高度表达;PtMAP65-3在木质部中表达量最高;PtMAP65-1和PtMAP65-4主要在幼叶和雌花中表达;PtMAP65-5在幼叶和根中表达水平较高;PtMAP65-9在各个组织表达水平较低。
3 讨论
MAP65 蛋白最早在烟草中发现,是一类具有稳定微管功能的蛋白[3]。本研究在毛果杨中鉴定出9 个PtMAP65基因,保守结构域分析发现,MAP65_ASE(pfam03999)结构域普遍存在于拟南芥、毛果杨、水稻和烟草MAP65 蛋白家族中[16],暗示MAP65_ASE 为植物MAP65 家族行使共同功能的主要结构域。研究发现,植物MAP65的C 端包含一个高度分化的微管结合结构域[20],这个结构域参与微管聚合、成束,且对其亚细胞定位有一定影响[21],在有丝分裂中期,C 端蛋白的磷酸化可及时地下调MAP65与微管的结合[22-23]。进一步分析发现,毛果杨PtMAP65 蛋白的C 端同样包含一个高度分化的微管结合结构域,这可能与毛果杨MAP65家族的功能分化相关。
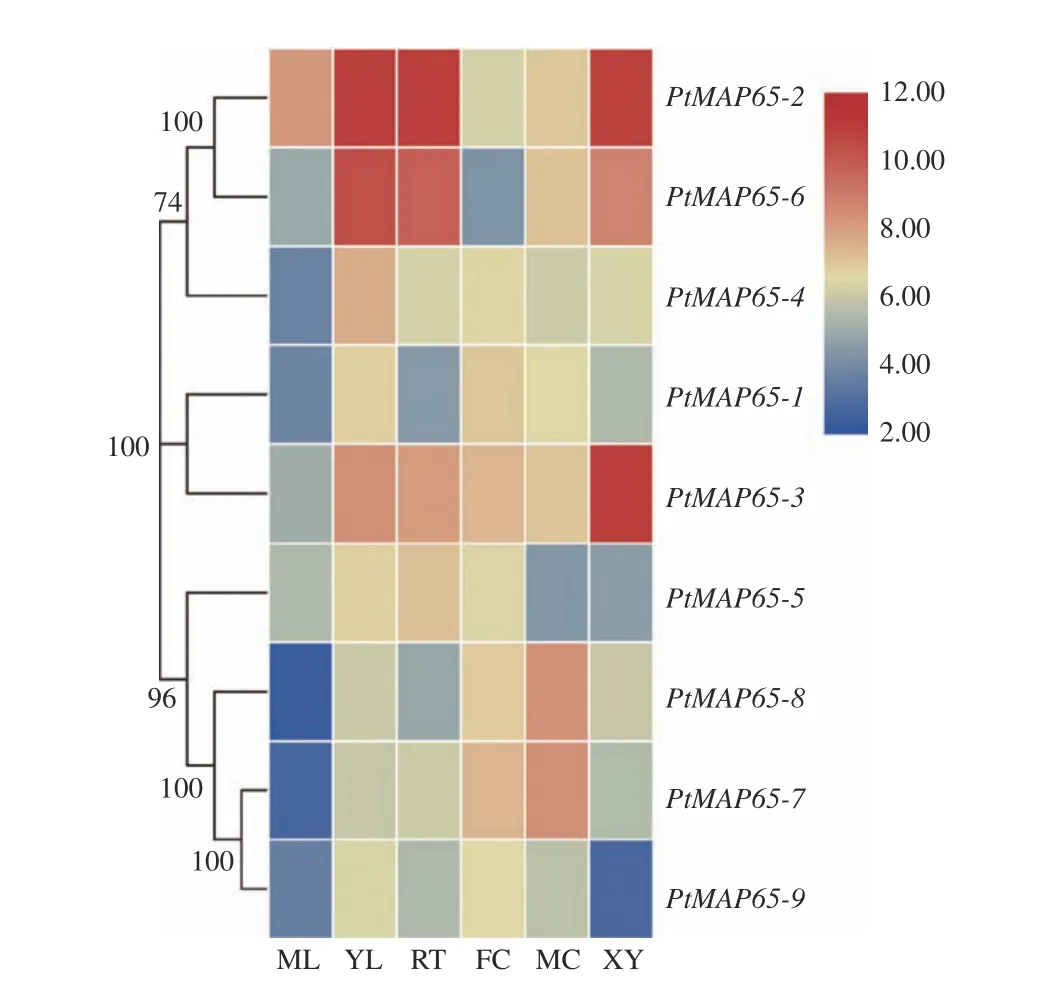
图5 PtMAP65 基因家族在不同组织中的表达模式Fig. 5 Expression profiles of PtMAP65 gene family in different tissues
片段复制和串联复制是基因家族扩张的主要动力[24]。毛果杨基因组内共线性分析发现,PtMAP65基因家族中共有6 对复制基因,且均为片段复制,这说明片段复制是PtMAP65基因家族进化扩张的主要动力。毛果杨经历了3 次全基因组复制事件,分别是古老的复制事件、100~120 百万年前的真蔷薇分支复制事件和60~65 百万年前的杨柳科复制事件[18]。分析PtMAP65复制基因发生时间发现,PtMAP65-7/PtMAP65-8、PtMAP65-8/PtMAP65-9的复制时间与真蔷薇分支复制事件接近,PtMAP65-7/PtMAP65-9的复制大约发生于为13 百万年前,由此推测,位于Chr14 的PtMAP65-8经过2 次片段复制或者三倍化复制得到位于Chr12 的PtMAP65-7和位于Chr15 的PtMAP65-9。最新研究表明,被子植物在20 百万年前亦发生了1 次全基因组复制事件,当时气温和CO2浓度均较低,保留下来的基因多与盐胁迫、低温胁迫、干旱胁迫等非生物胁迫有关[25]。分析发现,PtMAP65-1/PtMAP65-3、PtMAP65-2/PtMAP65-6、PtMAP65-4/Potri.001G032566.2,PtMAP65-7/PtMAP65-9这4 对基因的复制时间均与此次全基因组复制事件的时间相吻合。
通过对植物MAP65系统发育关系的研究发现,PtMAP65多与ATMAP65或巨桉MAP65聚为一支,这说明毛果杨和拟南芥、巨桉的亲缘关系最相近。前人研究表明:ATMAP65-1定位于除萼片和花药以外的所有植物器官和组织,且在根表皮细胞、子叶和下胚轴细胞中发现ATMAP65-1蛋白与微管相结合,在体外ATMAP65-1在微管之间形成25 nm 的交联桥,促进微管成束[26];ATMAP65-2在调节细胞的轴向生长方面具有重要作用;在ATMAP65-1和ATMAP65-2双突变体中,没有观察到明显的膨胀、变形和其他形态学缺陷的细胞,但植株生长显著迟缓[11]。分析发现,PtMAP65-2、PtMAP65-6与ATMAP65-1、ATMAP65-2聚 为 一支,位于Ⅰ亚组,且其在雄花和雌花中表达量低,在其他组织中表达量均较高,这暗示PtMAP65-2和PtMAP65-6可能通过参与细胞骨架的构建进而影响植株形态发生。ATMAP65-3与有丝分裂微管列阵紧密相关,在有丝分裂过程中,该蛋白组织纺锤体形态发生和成膜体形成,ATMAP65-3基因突变体根尖、茎尖分裂能力弱,植株矮小[27];引入ATMAP65-4拷贝基因的植株显著抑制了由ATMAP65-3突变引起的细胞质分裂和幼苗生长缺陷,而ATMAP65-3和ATMAP65-4的同时突变将导致植物死亡[13]。分析发现,PtMAP65-7/PtMAP65-9与ATMAP65-3聚为一簇,PtMAP65-8与ATMAP65-4聚为一簇,位于Ⅴ亚组,暗示PtMAP65-7、PtMAP65-8和PtMAP65-9这3 个基因通过参与细胞分裂过程决定植物的生长发育情况,但这3 个基因在毛果杨的各个组织中表达量较低,其中的调控方式尚需进一步实验来明确。ATMAP65-8在与周质微管的结合较弱,可能仅结合周质微管中的某一部分[28]。分析发现,PtMAP65-1/PtMAP65-3与ATMAP65-8聚为一支,位于Ⅲ亚组,这对基因的复制发生于20 百万年前的全基因组复制事件,预测的亚细胞定位在细胞核,特别的是:PtMAP65-3启动子上的激素响应元件远远高于其他基因,且该基因在木质部高度表达,推测PtMAP65-3可能通过参与各种非生物胁迫响应而影响植物次生生长。综上,PtMAP65-1、PtMAP65-2、PtMAP65-3、PtMAP65-6、PtMAP65-7、PtMAP65-8和PtMAP65-9可能紧密结合微管,通过影响细胞分裂进而影响植物生长,具体的作用机制及生物学功能仍需进一步研究。
4 结论
本研究利用BLASTP 比对,在毛果杨中鉴定了9 个PtMAP65s;构建多个物种的MAP65s系统进化树,将其分为5 个亚组;按不同亚组将基因结构图和保守结构域图归类比较,同一亚组的MAP65s具有相似的基因结构和保守结构域;利用毛果杨基因组内共线性及Ka、Ks分析PtMAP65基因家族的进化,片段复制是其扩张的主要动力,且复制基因在进化过程中受到强纯化选择的作用;亚细胞定位预测、基于芯片数据的表达量和启动子的顺式作用元件综合分析表明:在PtMAP65基因家族中,不同亚组间的PtMAP65s已经存在功能分化,同一亚组内的PtMAP65s即复制基因对之间仍存在功能冗余。
