麻栎天然群体种子表型变异分析
2021-04-10解孝满李文清
吕 锋,解孝满,韩 彪,乔 婕,董 昕,吴 丹,陆 璐,李文清*
(1. 山东师范大学生命科学学院,山东 济南 250014;2. 山东省林木种质资源中心,山东 济南 250102)
麻栎(Quercus acutissimaCarruth.) 属于壳斗科(Fagaceae)栎属(Quercus) 植物,在我国分布十分广泛,是一种重要的珍贵用材树种[1],可作为木材、菌材和薪炭材,具有较高的经济价值和利用价值;其种子富含淀粉和脂肪油,可酿酒、制作饲料,也可食用,具有非常广阔的发展前景。种子是植物有性生殖最重要的器官,是联系上下代植物体不可替代的纽带[2]。20 世纪50 年代以来,种子的微形态特征被一些学者应用于植物系统学和分类研究,证明了种子微形态特征在科、属、种等不同层次上具有分类学意义[3-5]。已知的大多数森林树种是通过种子发育生长为生殖成熟的个体,从而周而复始的繁殖下去,生长在不同环境中的林木经过漫长的演化过程形成了对各自生境的适应性,各部分器官具有多样的形态变异,其中,种子的形态变异蕴含了许多较为稳定的可遗传变异,因此,种子是研究林木天然群体遗传变异的重要材料。
目前,对麻栎种质的相关研究主要集中在组织培养及快速繁殖技术[6],苗期生长特性[7],优良家系的选择[8]和群体遗传多样性[9-10]等,对表型多样性的研究鲜有报道。表型变异是基因型与环境相互作用的结果,是探索群体遗传分化和变异来源的重要层面。因此,本研究通过对7 个麻栎天然群体种子表型变异进行分析,以期揭示麻栎表型变异大小及规律,为促进麻栎种质资源保护和遗传改良提供科学依据。
1 材料与方法
1.1 试验材料
供试的麻栎种子于2019 年9 月至10 月分别采自麻栎自然分布区(25°21′~39°49′ N,100°54′~121°59′ E)的7 个天然群体。麻栎林中选择优势树种作为母树,采种母树之间相距50 m 以上,每一群体选取20~30 株母树采集种子作为1 份,每份种子不少于300 粒,将采集的所有种子进行充分混合,然后将其用作群体的种子样品,为增加实验的准确性,将每个地理种源都使用了GPS 定位软件进行定位。种子来源情况详见表1。
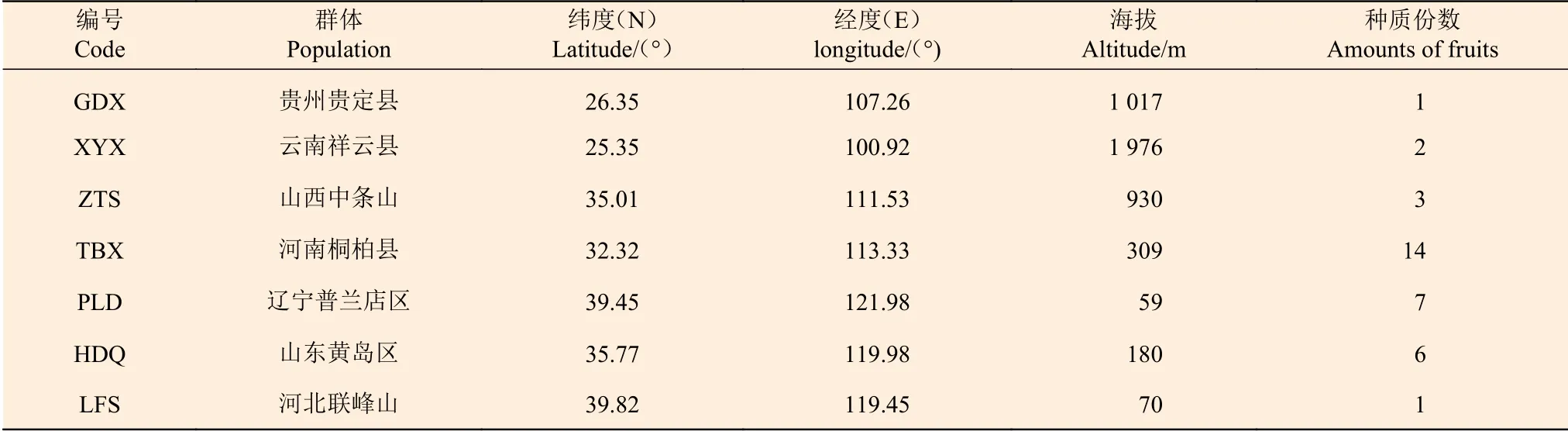
表1 麻栎群体采种母树来源Table 1 The source of the mother trees collected by the population of Quercus acutissima
1.2 表型性状测定及分析
对不同群体的种子长、种子宽、种子长宽比、表面积和体积等利用扫描仪Epson/V700 PHOTO 测定表型性状,采用winseedle Pro 2013b 软件可获得表型性状指标,使用1/100 电子天平测定千粒质量,从每组样品分别随机选取50 粒种子称质量,重复3 次,取其平均值。
采用巢式线性模型对麻栎群体间和群体内的表型变异进行方差分析。用变异系数(CV)表示表型性状的离散程度。计算表型分化系数(Vst)来表示麻栎群体的表型分化程度,即群体间方差分量与方差总量的比值。计算公式分别为:100%,其中,δ为标准差、为群体平均值,Vst=[δ2t/s/(δ2t/s+δ2s)] × 100%,其中,δ2t/s为群体间的方差分量,δ2s为群体内的方差分量[11]。采用Excel 2010 软件对数据进行统计,用SPSS 21 软件对各项指标进行分析,其中,采用Pearson 相关分析双变量相关性,并根据各性状在群体间的欧氏距离进行聚类分析。
2 结果与分析
2.1 麻栎种子性状的群体间变异
由表2 的方差分析结果可知:在群体间和群体内,种子长、宽、长宽比、表面积、体积及千粒质量均差异极显著(P< 0.01),说明麻栎种子表型性状在群体间存在一定程度的变异。
根据表3 可以看出:各群体种子长度的平均值为1.63~2.32 cm,其中,最大的是云南祥云县群体,最小的是贵州贵定县群体;各群体种子宽度的平均值为1.33~1.84 cm,贵州贵定县群体最小,云南祥云县群体最大;长宽比最大的是山西中条山群体,最小的是河北联峰山群体;种子体积的平均值为1.48~4.15 cm3,其中,最小的是贵州贵定县群体,最大的是云南祥云县群体;种子表面积平均值为5.19~10.41 cm2,最小的是贵州贵定县群体,最大的是云南祥云县群体;种子千粒质量差异最大,平均变幅为1.74~4.82 kg,贵州贵定县群体最小,云南祥云县群体最大。
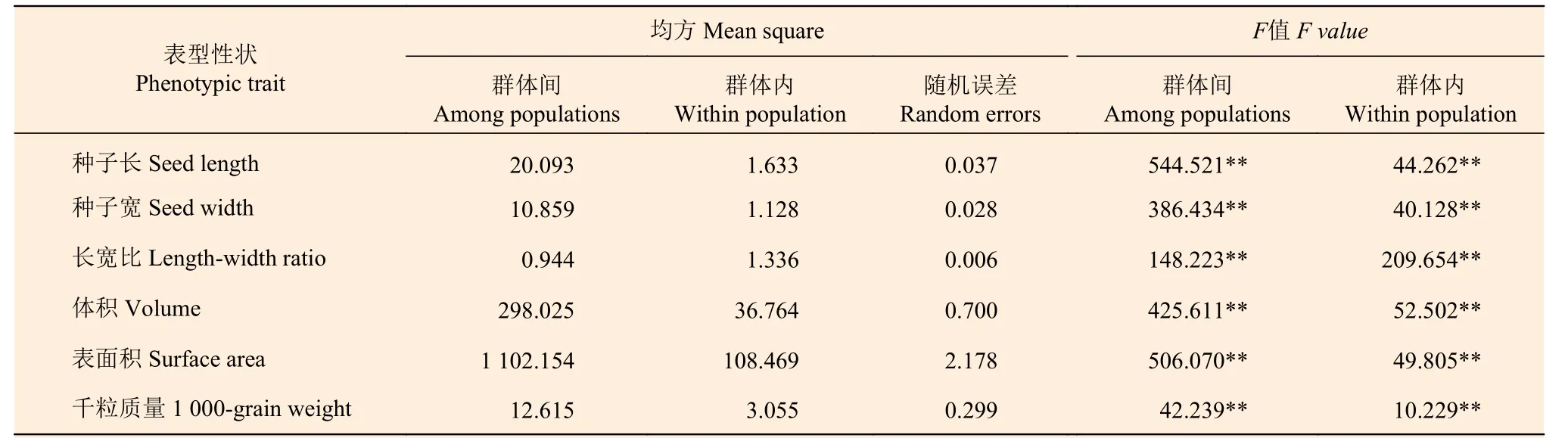
表2 麻栎群体种子表型性状方差分析表Table 2 Table of variance analysis of phenotypic traits of Quercus acutissima populations seeds
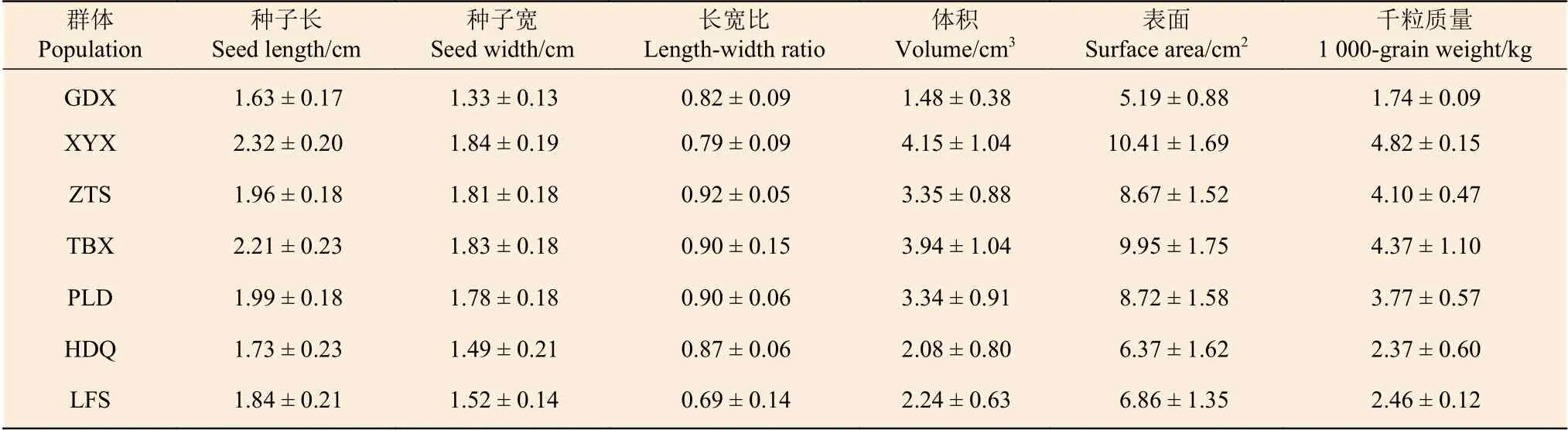
表3 麻栎群体种子的平均值和标准差Table 3 Mean and standard deviation of Quercus acutissima population seeds
2.2 麻栎种子性状的群体内变异
分析和比较不同表型性状时,性状的离散程度用变异系数来表示[12],变异系数越大,性状的变异程度越大;变异系数越小,性状的变异程度越小。由表4 可知:在麻栎的6 个表型性状中,平均变异系数最大的为种子体积,达28.18%,其次是种子表面积(18.79%),变异系数最小的为种子长(10.34%),表明种子长的变异程度最小,而种子体积这一性状的变异程度最大。
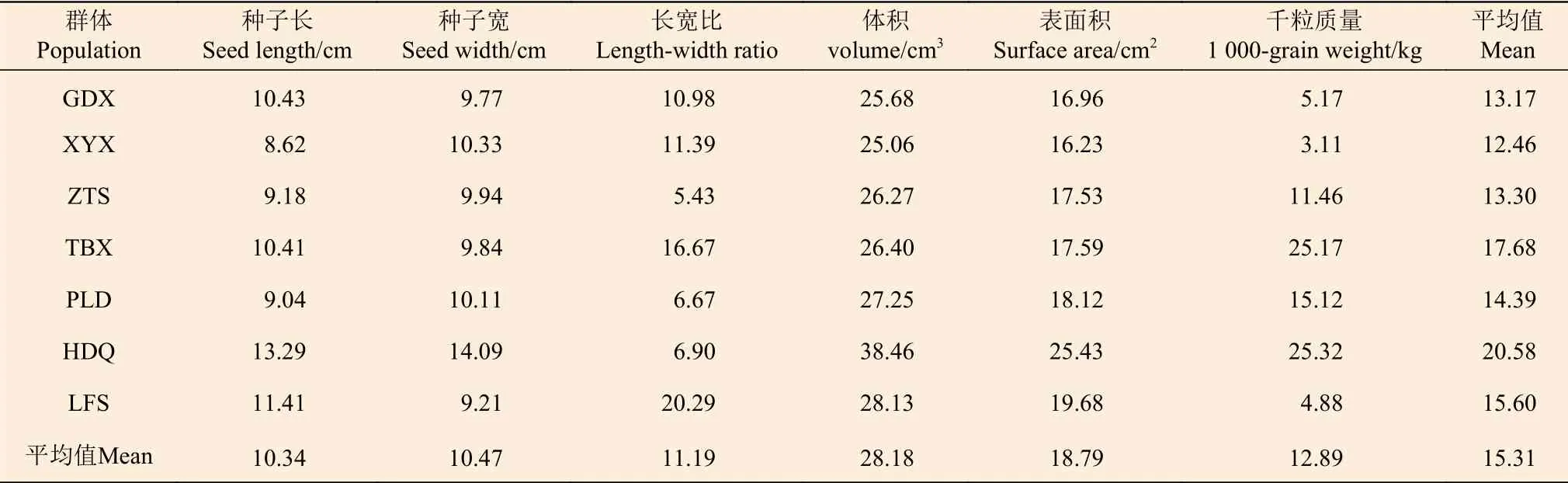
表4 麻栎群体种子表型性状变异系数Table 4 Table of variation coefficient of phenotypic traits in Quercus acutissima population seeds %
2.3 麻栎种子表型性状的群体间分化
由表5 可以看出:在表型性状的总方差分量占比中,群体间方差分量占比51.29%,群体内方差分量占比13.64%,机误占比35.07%。从群体间表型性状的变异幅度看,群体间种子长宽比的方差分量百分比最小,为41.67%;而千粒质量的方差分量百分比最大,为72.20%。从群体内表型性状的变异幅度看,种子宽的方差分量百分比最大,其次为种子长,最小的是长宽比。种子表型分化系数(Vst)为73.17%~84.34%,其中,千粒质量的Vst最大,种子宽的Vst最小;表型分化系数的平均值为78.84%,表明麻栎群体78.84%的变异来自群体间。
2.4 表型性状间的相关分析
根据表6 可知:各种子表型性状中两两性状具有极显著的相关关系,其中,千粒质量与长宽比的相关系数较小,为0.289;而种子宽和种子体积的相关系数达0.907。说明选择的6 个表型性状进行麻栎种子表型性状分析是可行的。
2.5 麻栎群体表型聚类分析
利用欧氏平方距离系数对麻栎群体进行聚类,图1 表明:以欧氏距离10 为阈值,7 个麻栎群体可以划分为2 大类,第1 类包括山西中条山、辽宁普兰店区、河南桐柏县、云南祥云县群体,第2 类包括贵州贵定县、山东黄岛区、河北联峰山群体。
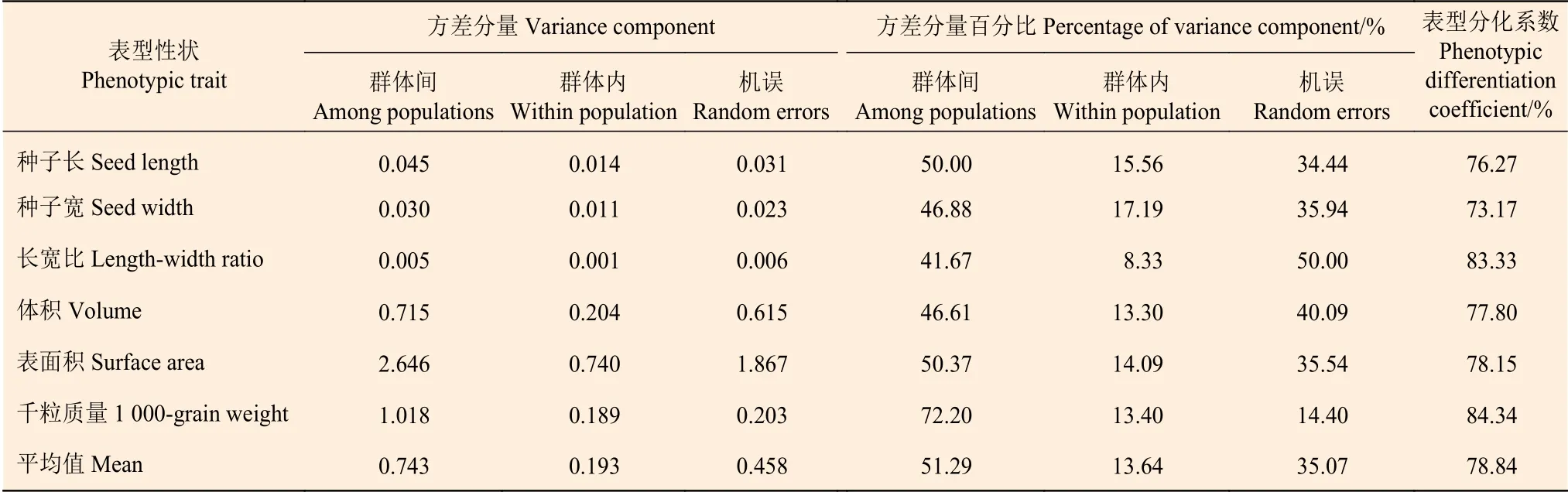
表5 麻栎群体种子表型性状方差分量及表型分化系数Table 5 Table of variance components of phenotypic traits and phenotypic differentiation coefficient of Quercus acutissima population seeds
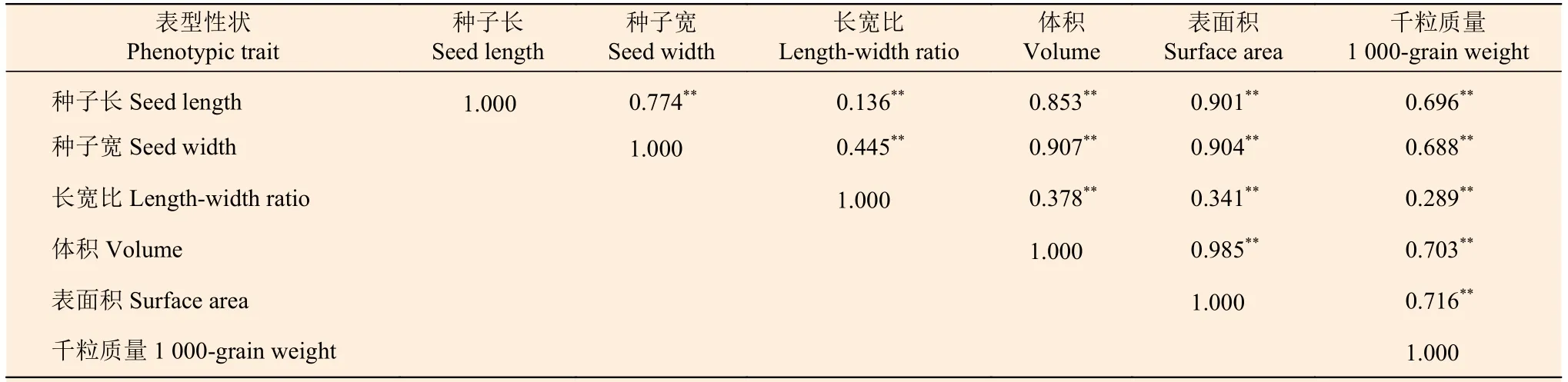
表6 麻栎种子表型性状的相关分析和差异显著性检验Table 6 Correlation analysis and difference significance test of phenotypic traits of Quercus acutissima seeds
3 讨论
3.1 麻栎群体种子表型变异多样性
表型变异既是生物遗传变异最直接的表现形式,也是遗传多样性研究的重要内容之一[13-14]。通常分布范围广泛的树种都具有丰富的表型变异,本研究以麻栎群体种子为材料进行表型性状变异的分析,可以在一定程度上反映其群体遗传多样性水平[15-16]。
由巢式方差分析结果可知:7 个麻栎群体种子表型性状在群体间和群体内均差异极显著,表明麻栎种子遗传变异较丰富,不同群体的地理环境不同使得麻栎种子性状变异差异明显,为下一步进行优良种源和家系的选择奠定了基础。麻栎群体间的变异大于群体内,这与许洋等[17]、刘志龙等[18]及张元燕等[19]的研究结论相反,可能是与遗传因素和环境因素的共同作用有关。此外,麻栎结实有明显的“大小年”现象,而且不同群体间种子的成熟期不一致,可能会造成个体的差异,所以,群体间的变异程度也可能与个体的差异有关。各个不同性状中,种子体积的变异系数最大,为28.18%,种子长的变异系数最小,为10.34%,说明种子长这一性状较稳定,这与常恩福等[20]、梁德洋等[21]及厉月桥等[22]对锥连栎和蒙古栎的研究基本上一致。7 个群体中,平均变异系数最大的是青岛黄岛区群体,为20.58%,平均变异系数最小的是贵州贵定县群体,为13.17%,说明山东黄岛区麻栎群体的生境条件相对复杂。麻栎群体平均表型分化系数为78.84%,表明麻栎群体表型变异在群体间的贡献率为78.84%,变异来源以群体间为主,群体间差异反映了地理隔离和生殖隔离的差异,群体间多样性是物种内多样性的重要组成部分[23]。因此,在麻栎种质资源保护工作中应当注意对群体间的变异进行合理利用。
3.2 麻栎表型性状的地理变异
相关性分析结果表明:麻栎种子长和种子宽、千粒质量均呈极显著正相关关系,这与厉月桥等[24]对不同种源蒙古栎种子表型性状的研究结果相近,说明种子长增加,其种子宽和千粒质量也极显著的增加。根据聚类分析结果,将7 个麻栎群体划分成2 个组群,发现麻栎群体并没有完全按照地理距离远近而聚类,与地域没有明显相关性,这可能与不同的地理环境有关,此结果与栎属的其他树种白栎[25]、铁橡栎[26]和栓皮栎[27]的种子表型性状的研究基本吻合。千粒质量可以作为检测种子品质的一个指标,千粒质量越大,种子品质越好,将麻栎群体聚成了2 类,第1 类代表千粒质量大的,包括山西中条山、辽宁普兰店区、河南桐柏县、云南祥云县群体;第2 类代表千粒质量较小的,包括贵州贵定县、山东黄岛区、河北联峰山群体。不同种源麻栎种子千粒质量具有一定的地理变异模式,可能与分布区域的环境因素有关,如光照、温度、经纬度、降雨及海拔等因素相互作用造成了不同群体麻栎种子千粒质量的不同变异模式[28]。
表型变异既受遗传控制,也受环境因素的影响[29-31],表型多样性是遗传和环境影响的结果。然而,表型变异中有多少遗传成分可以被利用,需要进一步深入研究,下一步有必要选择合适的分子标记开展麻栎分子水平的变异分析,为有效保护麻栎种质资源提供可靠的理论依据,提高麻栎的利用价值。
4 结论
麻栎天然群体种子在群体间和群体内都达到了极显著的水平,其遗传变异较丰富,是遗传因素和环境因素共同作用的结果,且群体间的变异大于群体内。研究结果表明,各表型性状间呈极显著相关关系。聚类分析将7 个群体分为2 个组群,与各个群体的地理位置分布无明显相关性。麻栎天然群体表型变异的研究,为麻栎多样性种质资源保护和利用提供了理论依据。
