A transgenic pig model expressing a CMV-ZsGreen1 reporter across an extensive array of tissues
2021-04-09AmyDesaulniersRebeccaCederbergElizabethCarreiroChannabasavaiahGurumurthyBrettWhite
Amy T. Desaulniers, Rebecca A. Cederberg, Elizabeth P. Carreiro, Channabasavaiah B. Gurumurthy,Brett R. White,✉
1Department of Animal Science, University of Nebraska-Lincoln, Lincoln, NE 68583-0908, USA;
2Department of Pharmacology and Experimental Neuroscience, University of Nebraska Medical Center, Omaha, NE 68198-5930, USA.
Abstract Since genetic engineering of pigs can benefit both biomedicine and agriculture, selecting a suitable gene promoter is critically important. The cytomegalovirus (CMV) promoter, which can robustly drive ubiquitous transgene expression, is commonly used at present, yet recent reports suggest tissue-specific activity in the pig.The objective of this study was to quantify ZsGreen1 protein (in lieu of CMV promoter activity) in tissues from pigs harboring a CMV-ZsGreen1 transgene with a single integration site. Tissue samples (n=35) were collected from neonatal hemizygous (n=3) and homozygous (n=3) piglets and ZsGreen1 abundance was determined via immunoblotting. ZsGreen1 was detected in all tissues, except hypothalamus, kidney cortex and oviduct. The expression patterns of homozygous and hemizygous piglets were similar (P>0.05). However, quantification revealed that ZsGreen1 protein levels were tissue-specific. Within neural/endocrine tissues, ZsGreen1 abundance was highest in the anterior pituitary gland, intermediate in the cerebellum and lowest in the cerebrum, spinal cord and posterior pituitary (P<0.05). In the digestive system, ZsGreen1 was more abundant in the salivary gland than esophagus, stomach, pancreas, duodenum, jejunum, ileum, spleen, colon, gallbladder and liver (P<0.05).Interestingly, ZsGreen1 amounts also differed within an organ (i.e., the right ventricle had 3-fold higher levels than the other heart chambers; P<0.05). These results provide useful information for the use of the CMV promoter to drive transgene expression in the pig. Moreover, this swine model represents a novel resource of ZsGreen1-labeled organs and a valuable tool to advance genome editing research.
Keywords: gene expression profiling, CMV promoter, ZsGreen1, biomedical model, genome editing
Introduction
The use of genetically engineered pig models has grown dramatically in recent years[1]. Transgenic swine are increasingly utilized as animal models for human diseases since they are anatomically,physiologically and phylogenetically more similar to humans than rodents[1]. In addition to biomedical applications, genetically engineered pigs are also developed to better understand porcine physiology and enhance the agricultural industry[2-4]. Towards this end, our laboratory produced a line of swine harboring a vector that contained: 1) the human U6 promoter fused to small hairpin RNA (shRNA) specific to the porcine gonadotropin-releasing hormone II (GnRH-II)receptor and 2) the human cytomegalovirus (CMV)promoter fused to the cDNA encoding the fluorescent protein, ZsGreen1[2]. Previously, we evaluated the impact of GnRH-II receptor knockdown (70%reduced testicular mRNA levels) on reproductive characteristics of male pigs[2]. Herein, we focus on the human CMV promoter and ZsGreen1 reporter aspects of this unique swine strain.
ZsGreen1 (Clontech, USA) is derived from Anthozoa reef coral (Zoanthusspecies)[5]and modified to maximize expression, solubility and prompt chromophore maturation[6]. It is both structurally and functionally distinct from other fluorescent proteins, like green fluorescent protein(GFP). The coding sequence of ZsGreen1 is only 26%homologous to GFP[5]and ZsGreen1 exists as a tetramer, whereas GFP (derived from luminescent jellyfish) is a monomer[6]. Functionally, ZsGreen1 displays extremely bright fluorescence[7-8]. In fact,yeast cells expressing ZsGreen exhibit 8.6-fold brighter fluorescence than those expressing enhanced GFP (EGFP)[7]. In addition, ZsGreen1, exhibiting high signal-to-noise ratio[7], is highly photo-stabile and unaffected by paraformaldehyde fixation[8]. Despite these advantages, no transgenic swine lines containing a ZsGreen1 reporter have been produced.
The human CMV immediate early promoter,discovered by Boshart and coworkers[9], has been heavily utilized to drive recombinant gene expression due to its constitutive and promiscuous nature[10]. The CMV promoter, now the most commonly used and most robust ubiquitous promoter[11], is utilized to drive transgene expression in numerous vertebrate animal models including sheep[12-13], fish[11], mice[14-15], rats[16],chickens[17]and pigs[18-19]. Pigs harboring a construct containing the CMV promoter fused to GFP express the transgene in a wide range of organs, including those derived from each of the 3 germ layers: skin(ectoderm), pancreas (endoderm), and kidney(mesoderm). Despite apparent universal transgene expression[19], the CMV promoter is preferentially active in exocrine compared to non-exocrine cells of the pig[18]. The objective of this study was to quantify ZsGreen1 protein levels (in lieu of CMV promoter activity) in a wide range of porcine tissues.
Materials and methods
Animals
All animal procedures followed the guidelines for the care and use of animals established by the University of Nebraska-Lincoln (UNL) Institutional Animal Care and Use Committee. These experiments utilized transgenic swine that expressed ZsGreen1 controlled by the human CMV immediate early promoter and were generated as previously described[2].Briefly, transgenic piglets were produced by microinjection of lentiviral particles into the perivitelline space of 1-cell porcine embryos. Lentiviral particles were made using the Lenti-X pLVX-shRNA2 vector and the HTX Packaging Mix (Clontech). The vector contains 2 ubiquitous promoters, CMV (driving ZsGreen1 expression) and the human U6 promoter(controlling the expression of a shRNA specific for the GnRHR-II gene). The background strain was a Large White × Landrace composite line (Nebraska Index line) developed and maintained[20]at the UNL Eastern Nebraska Research and Extension Center swine unit (Mead, NE, USA). Animals from this background strain were transported to the UNL Animal Science Building (Lincoln, NE, USA) and used to derive the CMV-ZsGreen1 transgenic line of pigs. This genetically-engineered swine line has been expanded, maintained and studied within the confines of the UNL Animal Science Building. In hemizygous animals, one copy of the CMV-ZsGreen1 transgene is stably integrated on chromosome 14[2], and has remained transcriptionally active throughout the lifetime of the founder animal and in 10 subsequent generations.
Adult animals were housed individually withad libitumaccess to water and fed approximately 2.5 kg of feed daily. Hemizygous CMV-ZsGreen1 transgenic sows were artificially inseminated with semen from non-littermate hemizygous transgenic boars and allowed to gestate to term. Following Mendelian inheritance, these matings yielded hemizygous,homozygous (2 copies of the transgene), and nontransgenic (control) littermates in the proportions expected with a single integration site. After farrowing, piglets remained with their dam to sucklead libitum.
At 1 day of age, 3 littermate female piglets (control,hemizygous transgenic and homozygous transgenic)were selected from each of the 3 different litters; each litter was produced from a different dam and sire mating to maximize genetic diversity. Initially,transgene status was assessedviaevaluation of ZsGreen1 expression using an ultraviolet (UV) light and a Roscolux #15 filter (deep straw; Rosco, USA).
Tissue collection
Piglets were first sedated with a mixture of telazol(Zoetis, USA), atropine (VetOne, USA), rompun(Bayer, USA) and ketamine (VetOne) (TARK; 0.2 mg/kg atropine, 0.8 mg/kg rompun, 3.1 mg/kg ketamine and 0.8 mg/kg telazol) delivered intramuscularly in the neck (1 mL/18-23 kg of body weight).Animals were then euthanizedviaintracardiac injection with Euthasol (1 mL/4.53 kg of body weight;Delmarva Laboratories, USA). Following euthanasia,tissue samples (n=35) generally representing 5 anatomical regions (brain, thoracic, digestive, renal and reproductive) were collected from each transgenic piglet. Isolated tissues included: cerebrum, cerebellum,spinal cord, anterior pituitary gland, posterior pituitary gland, hypothalamus, thymus, lymph node, the four chambers of the heart, lung, muscle, thyroid, salivary gland, esophagus, stomach, duodenum, jejunum,ileum, large intestine, gall bladder, spleen, pancreas,liver, kidney (medulla and cortex), bladder, adrenal gland (medulla and cortex), ovary, uterus, oviduct,and skin. Skin samples from control littermate piglets were collected to serve as a negative control for ZsGreen1 immunoblotting. In addition, tail samples were collected from all piglets for genotyping. All samples were frozen and stored at −80 °C until analysis.
DNA extraction
Genomic DNA was isolated from tail samples using enzymatic digestion and phenol/chloroform extraction.Briefly, a tail slice (approximately 1 mm) was incubated overnight in a lysing solution (50 mmol/L Tris [pH 8.0], 150 mmol/L NaCl, 10 mmol/L EDTA,1% sodium dodecyl sulfate [SDS], 0.1 mg/mL Proteinase K [Fisher Bioreagents, USA], and 1% βmercaptoethanol) at 55 °C. The samples were extracted twice with phenol: chloroform: isoamyl alcohol (25:24:1) solution and once with chloroform: isoamyl alcohol (24:1) solution before precipitating the DNA with isopropanol. DNA was subsequently resuspended in nuclease-free water.
Genotyping
Since we previously identified a single transgene integration site on chromosome 14 (aligning with clone NW_003612067.1 with 99% identity and matching identities 448 946-448 372)viasequencing analysis of inverse PCR products for this model[2],copy number was evaluatedviaconventional PCR using 3 primers. The forward (F) and reverse (R)primers were designed to flank the insertion site of the transgene, and an additional reverse primer aligning the transgene (RT) was included (F: 5′-GCAAC CTCTTCGACACTCCA-3′; R: 5′-AGCTACCAGGG AACAAAGCC-3′; RT: 5′-GGTTTCCCTTTCGCTT TCAAGT-3′). An MJ Research PTC-200 Thermocycler(USA) was used for PCR reactions with the following conditions: 1×TaqBuffer A (Fisher Bioreagents) with a final concentration of 2.0 mmol/L MgCl2,200 μmol/L dNTPs, 300 nmol/L of each primer and 1.25 U ofTaqDNA Polymerase (Fisher Bioreagents).Cycling conditions were 95 °C for 2 minutes,followed by 35 cycles of 95 °C for 30 seconds, 56 °C for 30 seconds and 72 °C for 60 seconds and a final extension of 72 °C for 10 minutes. The resultant PCR products were subjected to electrophoresis on 1%agarose gel. Products generated from the F and R primer pair were 627 bp in length and reflected a wild type chromosome 14. Products from the F and RT primer pair resulted in an 847 bp product, indicating that the transgene was present.
Protein preparation
Protein extraction and sample preparation were performed as described previously[21]. Protein was extracted by homogenization of tissue samples in radioimmunoprecipitation assay (RIPA) buffer(1 mL/100 mg; 20 mmol/L Tris, 137 mmol/L NaCl,10% glycerol, 1% NP40, 0.1% SDS, 0.5% deoxycholic acid, 2 mmol/L EDTA, 1 mmol/L PMSF, 1% protease inhibitor cocktail and 1% phosphatase inhibitor cocktail) using a Biospec Tissue Tearor (USA).Protein concentrations were quantified using a bicinchoninic acid (BCA) assay (Pierce Biotechnology, USA) according to the manufacturer's instructions. Extracted protein was mixed with 4×loading dye (2% Tris [pH 6.8], 28% glycerol, 20%SDS and Orange G) containing 100 mmol/L DTT and frozen at −80 °C until immunoblotting.
Immunoblotting
Immunoblotting was performed as described previously[21]. Tissue samples were segregated by anatomical regions (brain, thoracic, digestive, renal and reproductive) and protein (10 or 20 μg) was separated using SDS-PAGE (15%) before transfer to an Immobilon-FL polyvinylidene difluoride membrane(Millipore, USA) pre-soaked in methanol. After electrophoretic transfer, nonspecific binding was blocked by incubation of blots in Odyssey Blocking Buffer(Cat. No. 927-40100; LI-COR Biosciences, USA) for approximately 3 hours at room temperature. Membranes were then incubated with a mouse monoclonal primary antibody directed against ZsGreen1 (1:1000;Cat. No. 632598; Clontech), diluted in Odyssey Blocking Buffer with 0.05% Tween-20 and shaken at 4 °C overnight. The following morning, membranes were rinsed in Tris-buffered saline with 0.05%Tween-20 (TBS-T) prior to incubation with a goat anti-mouse secondary antibody (1:15 000; IRDye 680;Cat. No. 926-32220; LI-COR Biosciences) in Odyssey Blocking Buffer plus 0.05% Tween-20 and 0.025%SDS for 1 hour at room temperature. Blots were rinsed with TBS-T to remove excess secondary antibody and briefly rinsed in TBS to remove Tween-20 prior to imaging.
Quantification of ZsGreen1 abundance
Blots were scanned on an Odyssey Infrared Imager(700 channel; intensity 5.0; 169-micron resolution; LICOR Biosciences) and converted to greyscale for analysis. Band density was quantitated with Odyssey imaging software (version 2.1; LI-COR Biosciences)which automatically subtracted background signal to determine densitometry. Since the abundance of common loading controls (e.g., β-actin) differed across tissue types, total protein levels within each lane were used as the loading control according to Eatonet al[22]. After ZsGreen1 imaging, membranes were stained with GelCode Blue Safe Protein Stain(ThermoScientific, USA) for approximately 1 hour and then rinsed with 50% methanol and 1% acetic acid solution for 1 hour. Blots were reimaged (700 channel; intensity 1.0; 169-micron resolution; LI-COR Biosciences), converted to greyscale and quantitated using Odyssey imaging software to determine the relative quantity of total protein present in each sample. Data were presented as the relative band density of ZsGreen1 normalized by the total protein levels within the entire lane as described previously[21].The negative control in this experiment was wild-type porcine skin. During quantification, ZsGreen1 was considered detectable when the relative band density of the transgenic sample was greater than the nonspecific signal detected for the control, which inherently lacked ZsGreen1. ZsGreen1 was considered undetectable when values were less than or equal to the relative band density of the control sample.
Statistical analysis
Analyses were performed using the Statistical Analysis System (version 9.4, SAS Institute Inc.USA). Analysis of variance (ANOVA) was performedviathe MIXED procedure of SAS with the original model including tissue type, genotype (hemizygous or homozygous) and their interaction as the fixed effects.There was no effect of genotype or tissue type by genotype interaction (P>0.05), so these effects were removed from the statistical model; thus, homozygous and hemizygous data were grouped together for the final analysis. Litter was included in the model as a random effect andP<0.05 was considered statistically significant. Results were presented as least squares means ± the standard error of the mean.
Results
Identification of hemizygous and homozygous CMV-ZsGreen1 transgenic piglets
The presence of ZsGreen1 in the skin of neonatal transgenic piglets was detected visually (Fig. 1A) andviaimmunoblotting (Fig. 1B), indicative of CMV promoter activity. Even without the UV light, transgenic animals were identifiableviathe yellow hue of their skin (data not shown). The genotype of the animals was confirmed by conventional PCR (Fig. 1D);a schematic depicting the genotyping method is available (Fig. 1C). ZsGreen1 was not detected in the skin of control piglets (Fig. 1A and B).
ZsGreen1 fluorescence within tissues of CMVZsGreen1 transgenic piglets
ZsGreen1 fluorescence was visually detectable within internal organs of transgenic piglets under UV light; fluorescence was absent in wild-type pig organs(Fig. 2A; right panel). Fluorescence appeared robust in the intestines (Fig. 2A; middle panel), cerebellum(Fig. 2B; left panel), anterior pituitary gland (Fig. 2B;middle panel) and heart (Fig. 2B; right panel), but modest within the stomach (Fig. 2A; middle panel).ZsGreen1 was not visually detectable in the cerebrum(Fig. 2B; left panel), posterior pituitary gland (Fig.2B; middle panel), liver or spleen (Fig. 2A; middle panel). Differential fluorescent patterns were observed within organs as well. In the brain, ZsGreen1 fluorescence was intense within the cerebellum but not visually apparent in the cerebrum (Fig. 2B; left panel). In addition, the highest expression of ZsGreen1 appeared in the right ventricle compared with the other regions of the heart (Fig. 2B; right panel).
Quantification of ZsGreen1 in transgenic porcine tissues
Neural and endocrine tissues of the brain
Quantification of relative band density indicated that ZsGreen1 was present in 5 of the 6 tissues examined but only undetectable within the hypothalamus (Fig. 3A). There was an effect of tissue type on ZsGreen1 protein levels within the brain and the nervous system (P<0.0001; Fig. 3A). The abundance of ZsGreen1 was the greatest in the anterior pituitary gland, intermediate in the cerebellum and lowest in the cerebrum, spinal cord and posterior pituitary gland (P<0.05), which were not different from each other (P>0.05;Fig. 3A).
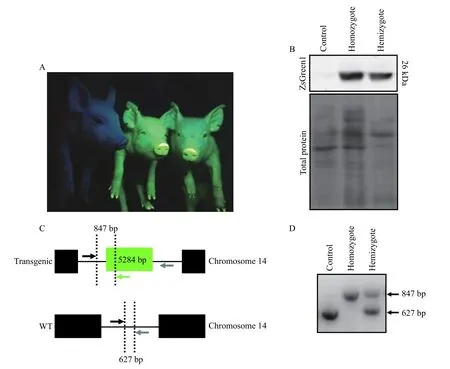
Fig. 1 Production of CMV-ZsGreen1 in hemizygous and homozygous transgenic piglets. A: Representative photograph of control piglet (left) with homozygous (middle) and hemizygous (right) transgenic siblings. ZsGreen1 was visualized with an ultraviolet light and a Roscolux #15 filter. B: Representative immunoblot of ZsGreen1 in the skin of control piglets (n=3) as well as hemizygous (n=3) and homozygous (n=3) transgenic littermates. Lower panel represents total protein levels within each lane. C: Schematic demonstrating the genotyping method to distinguish among control, homozygous and hemizygous piglets. Detection of monoallelic or biallelic transgene integration was performed by conventional PCR using 3 primers. The forward primer, denoted by the black arrow, amplifies a non-coding region upstream of the transgene integration site. A reverse primer (grey arrow) aligns a non-coding region downstream of the transgene and a second reverse primer (indicated in green), detects the transgene. If a transgene is present, the resulting PCR product will be 847 bp;however, in the absence of the transgene, the PCR product will be 627 bp. D: PCR detection of transgene integration and copy number. The presence of only an 847 bp PCR product indicates a homozygous transgenic animal whereas a 627 bp product indicates a control animal. If both 847 and 627 bp products are detected, the genotype is hemizygous transgenic.
Thoracic tissues
ZsGreen1 was detected by immunoblotting in all 9 thoracic tissues evaluated. An effect of tissue type was observed among the thoracic organs of CMV-ZsGreen1 piglets (P<0.0001;Fig. 3B). The right ventricle and skeletal muscle both produced more ZsGreen1 than the other tissues within the thoracic region (P<0.05),but did not differ from one another (P>0.05;Fig. 3B).Levels of ZsGreen1 in the thyroid, right atrium, left atrium, left ventricle, lung, thymus and lymph node were similar to each other (P>0.05;Fig. 3B).
Digestive tissues
ZsGreen1 was detected in all 11 digestive organs examined, but protein levels differed among tissue types (P<0.0001;Fig. 4). The ZsGreen1 level in the salivary gland was greater than those in the esophagus,stomach, pancreas, duodenum, jejunum, ileum, spleen,large intestine, gall bladder and liver (P<0.05;Fig. 4).Specifically, ZsGreen1 was 75-fold more abundant in the salivary gland compared to the pancreas. The protein amounts in the esophagus were higher than those in the pancreas, ileum, spleen, gall bladder and liver (P<0.05), but not within the stomach, duodenum,jejunum and large intestine (P>0.05;Fig. 4). ZsGreen1 levels in the other tissues (stomach, pancreas,duodenum, jejunum, ileum, spleen, large intestine,gall bladder and liver) were similar to each other(P>0.05;Fig. 4).
Renal and reproductive tissues
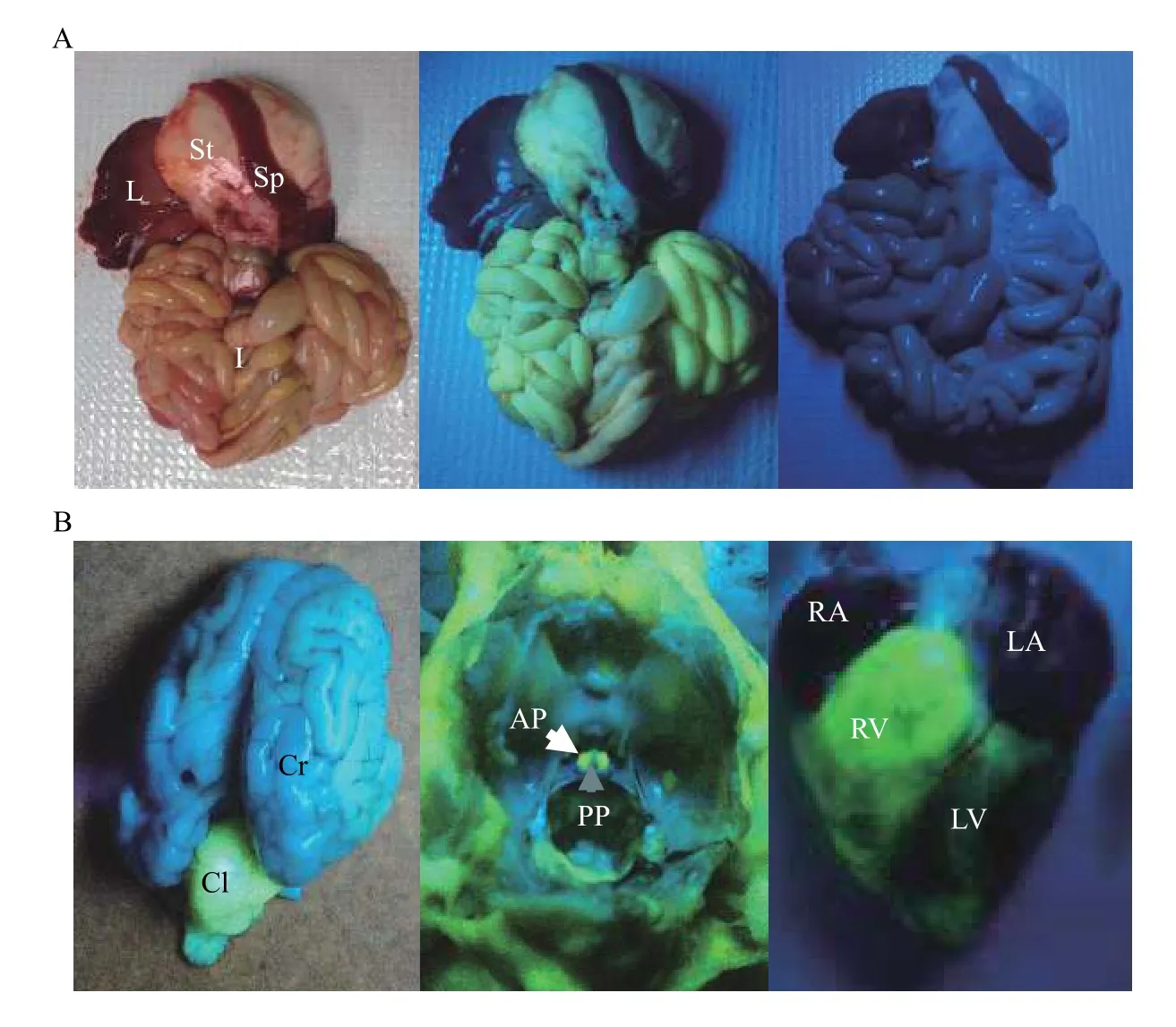
Fig. 2 Visualization of ZsGreen1 fluorescence within internal organs of transgenic piglets. A: Representative image of ZsGreen1 fluorescence in the internal organs of control and transgenic piglets. The left panel represents internal organs from a transgenic piglet. The middle panel represents a fluorescent image of internal organs from a transgenic piglet; and the right panel is a UV image of wild-type piglet organs. B: Representative fluorescent images of a transgenic piglet brain (left panel), anterior and posterior pituitary glands (within the sella turcica of the skull; middle panel) and heart (right panel). Anterior pituitary gland and posterior pituitary gland are indicated by white and grey arrows, respectively. ZsGreen1 expression was abundant within the right ventricle of the heart (right panel). ZsGreen1 fluorescence was detected with UV light and a Roscolux #15 filter. L: liver; St: stomach; Sp: spleen; I: intestines; Cr: cerebrum; Cl: cerebellum; AP: anterior pituitary; PP: posterior pituitary; RA: right atrium; RV: right ventricle; LA: left atrium; LV: left ventricle.
An effect of tissue type was observed among renal tissues that were screened (P=0.0001), with 4 out of 5 tissues expressing ZsGreen1. We were unable to detect ZsGreen1 within the kidney cortex (Fig. 5A).The medulla region of the adrenal gland produced more ZsGreen1 than the adrenal cortex, kidney medulla and bladder (P<0.05;Fig. 5A). Abundance of ZsGreen1 was higher in the adrenal cortex than the kidney medulla (P<0.05), whereas the level in the bladder was similar to that in both tissues (P>0.05;Fig. 5A). Within female reproductive tissues, we detected ZsGreen1 protein in the ovary and uterus, albeit at reduced levels, but not in the oviduct (Fig. 5B).However, no differences in ZsGreen1 abundance were detected between the ovary and uterus of female transgenic piglets (P=0.4016;Fig. 5B).
Discussion
In accord with the ubiquitous nature of the CMV promoter, ZsGreen1 protein was detected in all porcine tissues examined, except in the hypothalamus,kidney cortex and oviduct. Our results, however,demonstrated variable levels of CMV promoter activity across organs of the neonatal piglet. Other researchers observed similar results in animals expressing a transgene driven by the CMV promoter[11,17-18,23]. Several hypotheses regarding the differential activity of the CMV promoterin vivohave arisen. The activity of the CMV promoter within a given cell type may be influenced by integration site[24]; namely, activation may only occur in transcriptionally active regions of the genome[10].Stochastic silencing can happen when a transgene is integrated near a heterochromatic region, such as a centromere[24], or when multiple copies of the transgene integrate in tandem[25], due to local formation of heterochromatin[26]. Promoter suppression could also be caused by methylation, which can silence transgenesin vitroandin vivo[12-14]. Furthet al[23]observed marked differences in CMV promoter activity across tissues in two transgenic mouse lines harboring the same transgene. Conversely, Vaseyet al[18]detected a similar CMV promoter activity profile in pigs and chickens harboring the same transgene, albeit integrated in different chromosomal regions[18-19].The activity patterns were also similar in multiple transgenic chicken lines expressing different reporter genes directed by the CMV promoter[17]. Like Vaseyet al[18], we observed widespread CMV activity throughout CMV-ZsGreen1 pigs with robust activity in the skin and salivary glands, despite different integration sites and transgenes.

Fig. 3 Protein levels of ZsGreen1 within brain and thoracic tissues of CMV-ZsGreen1 piglets. A: Representative immunoblot of neural and endocrine tissues from CMV-ZsGreen1 transgenic piglets (n=6) using an antibody directed against ZsGreen1 (the upper panel). The middle panel represents total protein levels serving as the loading control. Quantification of immunoblots revealed differences in relative ZsGreen1 protein levels between tissue types (P<0.0001; the lower panel). B: Representative immunoblot of the thoracic tissue from CMVZsGreen1 transgenic piglets (n=6) using an antibody directed against ZsGreen1 (the upper panel). The middle panel represents total protein levels serving as the loading control. Quantification of immunoblots revealed differences in relative ZsGreen1 protein levels between tissue types (P<0.0001; the lower panel). Bars in a, b, c, with alternate letters differ (P<0.05). ND: not detected.
In addition to chromatin status, the repertoire of transcription factors within the cell dictate CMV promoter activity[10]. Mella-Alvaradoet alhypothesized that CMV promoter activity was robust in the cells that were most voraciously infected by CMV and reflective of optimal transcription factor expression[11].Numerous transcription factors have been implicated in the regulation of the CMV promoter, including nuclear factor kappa-light-chain-enhancer of activated B cells[10,18,27], yin yang 1[10], activator protein 1 and its major subunits (c-Fos and c-Jun)[27], p53,cAMP response element-binding protein/activating transcription factor, retinoic acid receptor[28],specificity protein 1[9], nuclear factor I, serum response factor, Elk-1 and CCAAT/enhancer-binding protein[29]. While we detected different ZsGreen1 protein levels between the cortex and medulla within the kidney and adrenal glands, we were unable to comprehensively evaluate the CMV-ZsGreen1 reporter among different cell types in this initial study.

Fig. 4 Protein levels of ZsGreen1 within digestive tissues of CMV-ZsGreen1 piglets. Representative immunoblot of digestive tissues from CMV-ZsGreen1 transgenic piglets (n=6) using an antibody directed against ZsGreen1 (the upper panel). The middle panel represents total protein levels serving as the loading control. Quantification of immunoblots revealed differences in relative ZsGreen1 protein levels between tissue types (P<0.0001; the lower panel). Bars in a, b, c, with alternate letters differ (P<0.05).
Consistent with elevated CMV promoter activity in exocrine glands, as previously noted in CMV-GFP pigs[18], ZsGreen1 protein levels were increased in the salivary glands of transgenic piglets. In the pancreas,however, we failed to detect elevated ZsGreen1 levels,conflicting with other reports about increased CMV promoter activity within the porcine pancreas[18-19].This discrepancy could be related to age-specific activity levels of the CMV promoter, as noted in zebrafish[11]. In our study, neonatal (1 day of age)piglets were utilized, whereas others examined CMV promoter activity in the pancreases of older (2 months to 1 year of age) pigs[18-19]. Likewise, we detected robust ZsGreen1 abundance in the pancreases of 1-month old piglets (unpublished data). Interestingly,the pancreas is largely quiescent in the neonatal piglet;the production of pancreatic juices and enzymes increase dramatically after weaning[30]. In the liver, we also detected little CMV promoter activity. Others reported similar levels of CMV promoter activity in porcine hepatocytes[18]and chicken liver[17], although,GFP was robustly driven by the CMV promoter in hepatocytes of zebrafish[11]. Thus, the CMV promoter is not uniformly effective in all exocrine organs and its activity varies across species.
Based on our visual observation of ZsGreen1 production in the skin of hemizygous and homozygous piglets (Fig. 1A), we hypothesized that an additional transgene would yield more ZsGreen1 protein in the organs of homozygous animals.However, no genotype effect was detected based upon quantification of immunoblots for ZsGreen1. These results may be due to monoallelic expression, which has been described in other transgenic animal models[25,31]. Conversely, translation of ZsGreen1 transcripts may be impaired in homozygous animals.Post-transcriptional silencing of high expressing transgenes is common in homozygous (but not hemizygous) plants and may be mediated by short interfering RNA[32]. Doubling transgene transcription may yield copious mRNA transcripts, breaching a threshold that triggers cellular processes to suppress transgene translation[33]. Nonetheless, transgene copy number did not significantly affect the production of ZsGreen1 protein in this study.
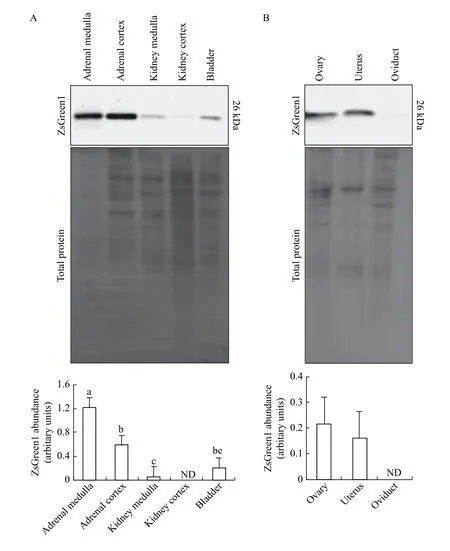
Fig. 5 Protein levels of ZsGreen1 within renal and reproductive tissues of CMV-ZsGreen1 piglets. A: Representative immunoblot of the renal tissue from CMV-ZsGreen1 transgenic piglets (n=6) using an antibody directed against ZsGreen1 (the upper panel). The middle panel represents total protein levels serving as the loading control. Quantification of immunoblots revealed differences in relative ZsGreen1 protein levels between tissue types (P=0.0001; the lower panel). B: Representative immunoblot of the reproductive tissue from CMVZsGreen1 transgenic piglets (n=6) using an antibody directed against ZsGreen1 (upper panel). The middle panel represents total protein levels serving as the loading control. Quantification of immunoblots indicated that there were no differences in relative ZsGreen1 abundance between tissue types (P=0.4016; the lower panel). Bars in a, b, c, with alternate letters differ (P<0.05). ND: not detected.
Notably, this ZsGreen1 swine model is well suited for re-engineering the transgene locus since expression of the transgene is readily detectable, the transgene is present in a single copy and the integration site has been identified[2]. This swine line could be used to test somatic cell gene editing using CRISPR/Cas9 systems or base editor nucleases by targeting the ZsGreen1 transgene sequence. In addition, the model is a useful tool for testing germline genome editing approachesin vivosuch as Genome editingviaOviductal Nucleic Acids Delivery[34]. Finally, re-engineering the transgenic CMV-ZsGreen1 locus of this pig model can make it suitable for other applications. The ZsGreen1 coding sequence can be disrupted to create a frameshift mutation in ZsGreen1, creating a model as a reporter for homology directed repair studies.Alternatively, a floxed red fluorescent protein could be inserted upstream of the ZsGreen1 cassette to serve as a Cre recombinase reporter. Re-engineering of the locus can be achievedviaEasi-CRISPR, a highly efficient genome engineering method[35].
Acknowledgments
This project was partially supported by the Nebraska Agricultural Experiment Station with funding from the Hatch Multistate Research capacity funding program (Accession No. 1011129 to BRW),an Agriculture and Food Research Initiative (AFRI)Competitive Grant (2017-67015-26508 to BRW) and an AFRI ELI predoctoral fellowship (2017-67011-26036 to ATD) from the USDA National Institute of Food and Agriculture. The funding agency had no role in study design, data collection, interpretation, or manuscript generation. The authors wish to thank Ginger Mills for pig husbandry as well as Scott Kurz,Guilherme Cezar, Kyle Regan and Amanda Lambrecht for technical assistance. The authors would also like to express gratitude to Lelanya Yates for aiding in the dissections.
杂志排行

THE JOURNAL OF BIOMEDICAL RESEARCH的其它文章
- Generating a CRISPR knockout mouse through a strong premature termination codon: a cautionary tale
- Designing and generating a mouse model: frequently asked questions
- Harnessing CRISPR-Cas system diversity for gene editing technologies
- Base editing: a brief review and a practical example
- Therapeutic gene editing strategies using CRISPR-Cas9 for the β-hemoglobinopathies
- Genome engineering technologies in rabbits