增温增水对草地生态系统碳循环关键过程的影响
2021-04-08李博文吕汪汪姜丽丽刘培培孟凡栋张立荣张苏人李耀明斯确多吉汪诗平
李博文,王 奇,吕汪汪,周 阳,姜丽丽,刘培培,孟凡栋,张立荣,张苏人,阿 旺,李耀明,斯确多吉,3,汪诗平,3,*
1 中国科学院青藏高原研究所高寒生态重点实验室,北京 100101 2 中国科学院大学,北京 100049 3 中国科学院青藏高原地球科学创新卓越中心,北京 100101
陆地生态系统碳循环作为陆地生态系统过程的重要组成部分,是全球变化研究中的重点[1]。研究陆地生态系统碳循环是预测未来大气CO2和其他温室气体浓度变化、认识大气圈与生物圈的相互作用等科学问题的关键,也是认识地球系统水循环、养分循环和生物多样性变化的基础[2]。自工业革命以来,全球地表温度持续上升,预计在未来会以更快的速度增加[3],并且高寒区域气候变化幅度更大[4]。气候变化所带来的一系列环境问题(如全球变暖、降水时空分布格局变化等)及其对生态系统产生的重要影响越来越受到国际社会的重视[3,5]。
草地生态系统是陆地生态系统的重要组成成分,约占陆地总面积的30%[6],其碳循环过程发生微弱的变化,都可能会对陆地生态系统和大气间碳循环间的平衡产生重要影响,由此可见其在区域气候变化及碳循环过程中占有非常重要的地位,特别是高纬度冻原和高海拔高寒草地生态系统对气候变化的反应极其敏感[7-8]。近年来,温度和降水作为气候变化的重要指示指标在国内外的研究中备受关注。但是目前国内外关于增温和降水控制实验对草地生态系统碳循环的影响还存在很大的争议。比如在美国的高草草原研究表明增温能够促进碳的吸收[9],而内蒙古干旱草原的研究则认为增温对草地碳固存有不利影响[10]。同样的不一致性也出现在了模拟降水试验中[11-12]。此外,在国内外的原位增温实验中,无论是利用开放式气顶箱(open top chamber, OTC)增温还是红外加热器增温,大都是设计增温和不增温2个梯度的试验[13- 15]。然而总体上看,气候变暖是一个逐步升温的过程[3],而目前的增温试验大都设置的恒定增温,与实际情况相比有着很大的不一致性。所以研究温度逐渐升高的情境下草地生态系统碳循环的响应方式具有很重要的现实意义,但目前这一研究非常匮乏,亟待加强。另外,研究表明温度对草地生态系统碳循环的影响受到水分的调节作用[10],但是这种调节作用在不同的草地类型下的结果不同[16-17]。本文综述了国内外有关增温增水对草地碳循环关键过程(图1)的研究成果,并指出了目前控制试验所存在的不足之处,并认为在以后的试验中应加强逐步增温控制试验及与水分耦合的研究,从生态系统层次更加深入地量化草地生态系统对温度和水分变化响应,这对于更好地了解草地生态系统对全球变化响应的机理具有十分重要的意义。
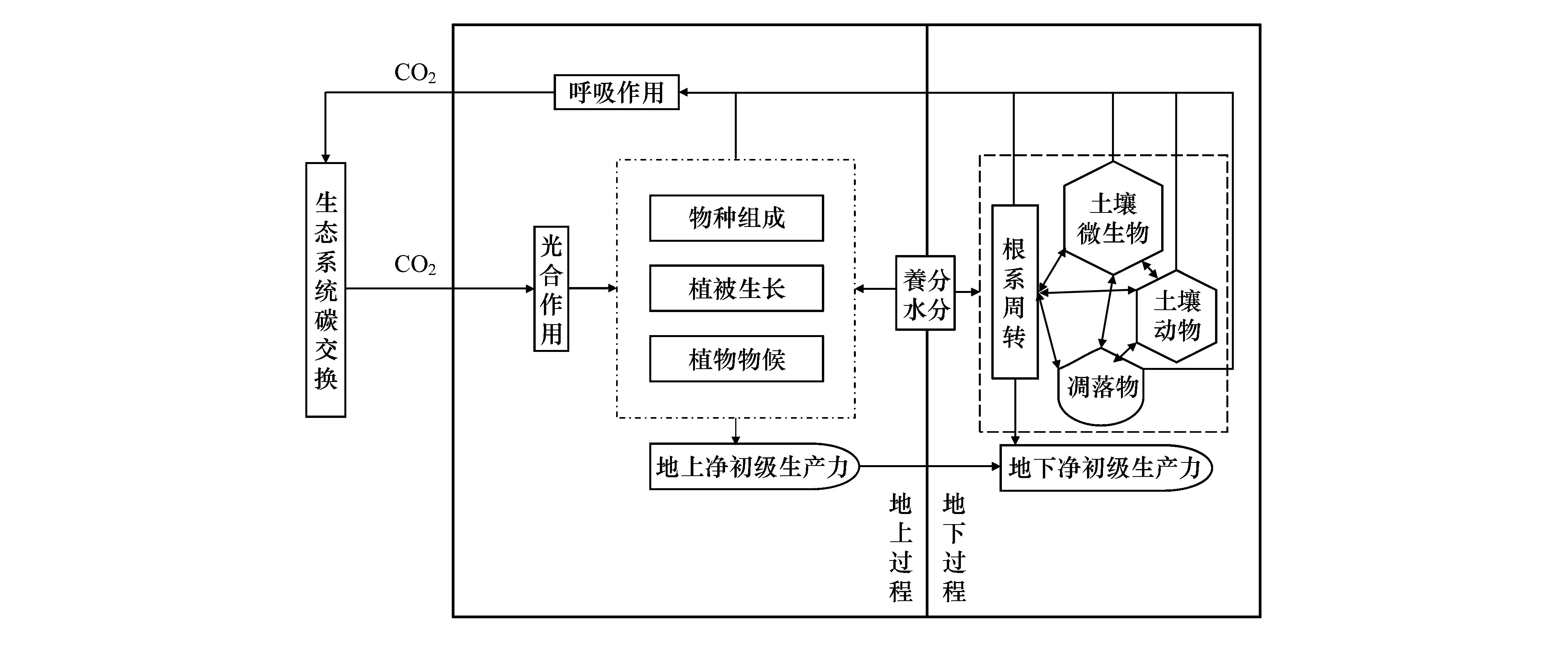
图1 草地碳循环关键过程示意图Fig.1 Schematic diagram of the key processes of grassland carbon cycle
1 增温对草地生态系统碳循环关键过程的影响
1.1 增温对植物群落组成和生产力的影响
过去10多年来,国内外在生态系统水平上开展了很多温度控制试验,发现增温对植物生产力的影响存在不一致的结果[13-14, 18]。增温可以通过影响土壤湿度和养分有效性而影响植物的生长、繁殖和资源竞争及分配策略[19],不同的植物或功能群的反应可能不尽相同,从而导致了物种组成和多样性的变化,进而影响到植物生产力的变化[13-14, 20]。如在北美半干旱草原的研究发现,温度升高改变群落物种组成(C3和C4植物的比例)[9]。Peng等[21]的研究发现,增温使高寒草甸植物功能群向禾草类和杂草类转变,而莎草类植物相对减少。Wang等[14]也有类似的发现。究其原因可能是,增温导致了土壤含水量下降,资源有效性降低,相比于莎草科植物,禾本科植物的根系较深且其能够被更多的丛枝菌根真菌浸染在面对干旱胁迫环境时,更加有利于自身获取水分和其他资源[22-23]。
研究发现生长季的温度和地上净初级生产力密切相关[24]。Shi等[25]在美国中部草原的八年增温试验表明,增温对草地生产力没有显著影响。但在美国的高原草原的研究则表明,增温降低草地生产力[26],而在高寒草甸则发现增温能够促进植物的生产力[14]。其原因可能是增温通过增加蒸散作用,从而减少植物水分的可利用性,进而影响到植物的生产力,而在不同的草地生态系统,由于其所处环境不同,所以导致温度对植物生产力的影响具有很大不确定性[24]。此外,研究还发现增温增加了土壤氮的可利用性[27],但是因研究地点和生态系统不同,所以增温对土壤氮的可利用性及植物生产力的影响出现不一致的结论[13-14]。特别是在高寒生态系统,植物利用小分子有机氮的能力更强,且随着不同季节土壤氮可利用性的变化而改变有机氮的利用能力以及与微生物竞争能力[28]。还有研究认为,只有在土壤氮有效性和水分不是制约因子时,增温才会提高植物生产力[29]。
1.2 增温对植物物候的影响
植物物候的变化受多种环境因子的调控,而在这众多的影响因子中,温度被认为是关键因子[13, 18, 30]。亚北极植物物候对温度的敏感性是非线性关系,即在温度较低时增温比温度高时增温效果明显;同时增温具有延迟效应,如果前一年或者前两年的温度较高,会显著延长当年的物候期,因为亚北极植物在开花前一年或多年发育叶原基[31];而在北美干旱草原,由于增温导致的土壤水分受限会使得植物的生长季缩短[32]。对于高寒草原而言,虽然生长于此的植物已经适应了低温环境,但是它们仍然受到低温环境的限制,即使小幅度的增温也能够显著地促进植物生长[33]。此外增温还能够促进一些地下过程(如凋落物分解过程),从而为植物返青提供充足的养分,促进植物生长[34]。目前关于增温对高寒草甸植物物候的影响,大多数的长期地面观测和增温控制试验的结论大致都一样:返青期和初花期随着持续增温而显著提前,枯黄期显著延迟,从而延长了整个植物生长季[18, 30, 35]。特别发现与其它物候期相比,结果期则保持相对稳定,而早期物候和晚期物种则基本保持同步性变化[36],增温通过延长开花期从而延长繁殖期和生长季[37],且中花植物对增温更敏感[38]。学者还认为个体植物物候很难预测群落物候的变化,因为群落物候还受到因温度变化诱导的植物开花功能群变化的影响[35, 39],且其非线性的变化与开花植物功能群多样性有关[40]。另外,与多年生植物相比,增温对一年生植物物候序列的影响很小,其主要受降水或土壤水分状况的影响,但增温增加了一年生植物的种子生产[41]。
1.3 增温对植物地下生产的影响
根系是植被碳库向土壤碳库进行物质运输及能量转移的重要通道,其本身也是植被的一个重要子碳库[42]。它们的生长和周转直接影响着生态系统碳循环[43]。而根系周转的快慢在很大程度上决定了植物碳库向土壤碳库转化的速率,进而对土壤有机质动态和生态系统碳固持能力产生重大影响。比如,有研究认为根系生物量和土壤可溶性有机碳的多少相关[44]。根系的生长与土壤水分和养分可利用性以及地上/地下碳分配紧密相关,这也造成了在不同的研究地点得到了研究结果不一致的现象[45-46]。Nelson等[42]在美国的半干旱草原研究表明,增温对根系生物量没有显著影响,但是却增加了根系的分解速率,且不同的植物响应不同,这可能和凋落物的质量或者氮含量有关[47]。内蒙古温性草原的研究发现,增温降低了根系生产力、现存生物量,但是对根系周转速率却没有显著影响[48]。然而在北欧多年生草地的研究则发现,增温增加了根系生产,植物向根系分配了更多的碳[49]。Wang等[50]在青藏高原高寒草甸的研究认为,增温导致了土壤水分受限,降低了表层土壤根系的生产力,提高了根系死亡率和周转速率。
1.4 增温对土壤和生态系统呼吸的影响
土壤呼吸是生态系统与大气之间进行CO2交换的第二大过程,每年大气CO2循环的10%左右是通过土壤进行的[51-52]。模拟增温试验表明,增温增加土壤呼吸[53-54]。增温引起的土壤呼吸的增加主要和增温引起地上生物量的增加、微生物活性、增加凋落物分解相关[55-56]。但是也有研究认为增温会抑制土壤呼吸。如Fang等[57]在半干旱区的研究表明,增温增加了土壤温度降低了土壤含水量从而导致微生物活性下降,抑制了土壤异养呼吸,但是对土壤自养呼吸没有影响;Wertin等[58]在美国半干旱区草原的研究表明,增温减少了光合作用产物,从而分配到根系的光合作用产物减少,自养呼吸会受到抑制,进而降低了土壤呼吸。高寒草甸的研究表明当增温引起土壤含水量的下降达到一个阈值时,会对土壤呼吸产生负效应[59]。之所以会产生这种不同的响应,可能是和土壤因子不同(如土壤质地,含水量,养分含量)有关[60]。此外还有研究认为增温对内蒙古半干旱草原的土壤呼吸没有显著影响[61]。
生态系统呼吸占据着陆地生态系统碳通量的很大一部分,即使生态系统呼吸微弱的波动,都会对大气中CO2浓度产生很大的影响[62]。Meta分析表明增温显著增加生态系统呼吸[63]。但是不同的试验地点又会有不同的结果。在美国高草草原,增温显著增加了生态系统呼吸[9],而在温性草原增温则没有显著影响[52];对于高寒草甸,有研究发现增温没有显著影响生态系统呼吸[64],但在此区域还有研究认为增温增加生态系统呼吸[65]或者降低了生态系统呼吸[66],这是增温引起自养呼吸和异养呼吸的改变不同导致的[67]。另外,增温还能够通过增加凋落物分解和土壤有机质的分解速率,从而增加土壤呼吸和异养呼吸[68- 70]。植物呼吸和植物生产力、土壤温度有关,土壤异养呼吸通过消耗微生物生物量碳、不稳定碳或土壤湿度来对增温产生响应。而增温对生态系统呼吸没有影响可能和增温引起土壤含水量下降,进而导致植物生产和微生物活性降低有关[71]。
1.5 增温对生态系统净碳交换的影响
生态系统净碳交换(Net Ecosystem C Exchange,NEE)是从生态系统到大气的CO2通量,它是生态系统通过光合作用吸收二氧化碳,并经过呼吸作用释放二氧化碳的一个平衡[72-73]。增温会通过促进植物生长和微生物活性来增加草地生态系统光合作用和呼吸作用,其中对处于温度受限的生态系统有更明显的影响[74-75]。Day等[76]对南极苔原研究表明,增温促进生态系统呼吸,同时增加了优势种的盖度,增加了总初级生产力(Gross Primary Production,GPP),但是后者比前者增加要大,所以表现为NEE增加,使得生态系统固定了更多的碳;而在内蒙古干旱草原的研究结果则认为,短期增温会使植被蒸腾加强,降低土壤水分,降低了总初级生产力,但是对生态系统呼吸影响不显著,所以会减少NEE,不利于碳固存[10]。但是Lu等[63]通过Meta分析发现,增温对NEE无显著影响。此外NEE对增温的响应还和群落组成有关,如Niu等[9]在美国高草草原研究发现,增温显著增加以C4植物为主的植物群落的生态系统呼吸,轻微提高总初级生产力,CO2的吸收降低;而以C3为主的植物群落,在增温条件下,生态系统呼吸和总初级生产力均受到抑制,生态系统呼吸对增温更敏感,受抑制程度更高,净效应表现为碳固定增加。在青藏高原高寒草甸的研究发现,增温增加NEE,促进生态系统碳的吸收[77]。
2 增水对草地生态系统碳循环关键过程的影响
2.1 增水对植物群落组成和生产力的影响
降水的改变同样会使物种多样性[78]和丰富度[79]发生变化。在青藏高原高寒草甸的模拟降水实验表明,增加50%降水增加了物种多样性[80]。同时,降水对植物地上部分净初级生产力(Aboveground Net Primary Production,ANPP)影响很大,在温度受限制的高寒区域,水分亏缺能够对ANPP产生一定的限制作用,使得生态系统对土壤水分有效性的改变极其敏感[81]。增水使得土壤含水量和土壤无机氮含量升高有利于ANPP的生产,尤其是杂草类的生产力对土壤含水量更敏感[82]。Yahdjian等[83]发现,在南美草原随着增水量的增加,ANPP和增水量表现出一种线性关系,但是Zhang等[82]在内蒙古草原的研究则表明ANPP对梯度增水量的响应是非线性的,这可能是前者的增水量没有达到使其出现负效应的阈值所致。此外Wu等[84]通过Meta分析发现ANPP对增水的敏感性要大于减水的敏感性。
2.2 增水对植物物候的影响
除了温度,降水也是影响物候的重要因素。有人认为水分有效性可能比温度更重要[85],因为持续时间较长的温度对植物物候影响作用是渐变的,而降水变化则会引起植物物候的突变,能够很大程度上决定物候期的早晚[86]。吕新苗等的研究发现,植物物候的年际变化对降水量的季节分配更敏感,降水的提前使得高寒草甸物候期提前20天[87]。非生长季降水的变化也会显著地影响植物的返青期[88]。此外,水分含量的增加也会显著地延迟晚期物候[89]。
2.3 增水对植物地下生产的影响
水分含量对地下根系的垂直分布有着很重要的影响。和地上生物量相比,地下生物量对水分变化的反应有很大的不确定性。Xu等[90]在内蒙古半干旱草原的研究发现,增加降水会显著增加地下根系生物量,因为在水分限制的区域,增加降水使得土壤含水量增加,土壤水分有效性的增加,有利于光合作用,从而会向地下分配更多的光合作用产物[10, 48],另外长期的降水增加会增加土壤氮损失,在降水增加的情况下,植物增加地下生物量,从而吸收更多的氮。Zhang等[82]在青藏高原高寒草甸的研究发现,虽然增水没有显著改变地下生物量,但是地下生产力与地上地下总生产力的比例却随着降水量的增加有降低的趋势。不过在半干旱草原的研究则发现地上地下的生物量分配对增水没有显著的响应[91]。虽然目前对于这一分配机制并没有研究清楚,但很可能是因为资源(比如水分、养分、光照)的相对重要性改变所致[92-93]。此外,在不同的生态系统下不同的物种组成同样会影响地下生物量对增水的响应[90, 94]。
2.4 增水对土壤和生态系统呼吸的影响
有研究表明呼吸的年际间变化是和降雨量的变化密切相关[95-96]。土壤含水量和土壤呼吸呈正相关,除此之外还和地下生物量以及微生物生物量、植物盖度呈正相关[97]。增水通过增加土壤含水量,增加了群落的盖度,从而增加了光合作用碳的吸收、根的生长、微生物的活性,进而增加了土壤呼吸[98]。同时增水引起的土壤呼吸的改变还和土壤本身的水分状况有关[99]。前人研究结果表明,改变降水会对土壤呼吸产生很大影响,而且土壤呼吸随着降水梯度的改变也会产生很大的变异。随着降水量的增加,土壤呼吸会达到一个峰值,之后会随着降雨量的增多而下降[100]。这可能是因为增加的降水会增加土壤淋溶,影响氮的有效性,从而导致土壤呼吸对增加降雨的非线性反应[101]。另外,在不同的生态系统土壤呼吸对降水变化的响应也有差异[10, 102- 104]。Wu等[84]综合分析31个降水实验研究结果发现,增加降水显著刺激了植物的生长、微生物的活性和生态系统呼吸,相反减少降水抑制了植物的生长、微生物的活性和生态系统呼吸,而生态系统呼吸对降水增加的敏感性均高于降水减少,并且不同的生态系统降水改变对生态系统呼吸的影响差异性很大。
2.5 增水对生态系统净碳交换的影响
半干旱草原对降雨的响应非常敏感,当增加土壤水分的可利用性时,生态系统总生产力会迅速加强,GPP与土壤温度的相关性也会进一步增加。因为植被生物量的增加会给根系生长、微生物活动、土壤呼吸提供更多基底,进而促进生态系统呼吸[10, 75]。在内蒙古干旱草原的研究表明,增加降水会对NEE产生抑制作用,这是因为增水促进了生态系统呼吸所致[11]。但是Chen等[12]的研究则认为,植被光合作用对土壤水分的敏感性要高于生态系统呼吸,因此对GPP的促进作用会比生态系统呼吸的更大,所以会使得NEE增加,促进二氧化碳的吸收。但是在相对比较湿润的环境下,增水对NEE的变化影响不显著[105-106]。耿晓东等[107]在对青藏高原高寒草甸的增水试验表明,增水促进了GPP的生产,但是对生态系统呼吸无显著影响,从而表现出促进了CO2的吸收,增大了NEE。有人认为青藏高原高寒草甸土壤干旱可能是在增温条件下限制生态系统碳摄取的关键因素[108]
3 增温和增水的耦合效应
温度和水分的变化会对生态系统过程和功能造成很大的影响。通过Meta分析发现,增温和增水的耦合效应要比单一因素的影响要低[84]。而Luo等[109]等通过模型模拟发现增温和增水的交互作用对NPP以及生态系统呼吸有显著的正影响。Ma等[80]在青藏高原的研究也认为,增加50%的降水能够增加群落的生物量而且在有增温的协同作用下增水的效应会更加明显。温度对植物的碳获取同样会随着土壤水分的变化而变化,比如连续的降水事件之后,增温会对生态系统过程产生正的影响,有利于碳吸收,但是在持续干旱,土壤水分含量较少情况下,会产生负效应[10, 110]。有研究发现,同时增温和增水对土壤温度以及土壤含水量并没有显著的改变,这是因为一方面通过增温,土壤温度增加,但是增水又会使得植物群落盖度增加,从而缓解了增温效应,而增温引起的土壤水分降低又会通过增水来补偿[75]。比如有研究认为,增温引起的土壤含水量下降对植物生产产生的负效应会被增水引起的土壤含水量上升对植物生产的正效应所抵消,从而表现为增温和增水对植物生产没有显著影响[15]。
4 问题与研究展望
目前国内外的原位增温实验中,无论是利用OTC增温还是红外加热器增温,基本上都是设计增温和不增温2个梯度的实验[13- 15]。但这些研究所呈现的只是一种恒定增温的模式(如与对照相比,温度恒定增加2℃或4℃),这显然不太符合自然条件下气候变化的现象。全球变暖在大的时间尺度上是一个逐步升温的过程,如过去50年来,以每十年0.19℃的速率增加,而最近30年的增温速率则达到了每十年0.25℃[111]。有研究认为,面对突然增温,植物和生态系统可能会发生改变,如在开始年份植物会产生适应增温环境的反应,表现为根系呼吸增加[112]和生物量增加[113],但随着增温年限的延长,植物和土壤或生态系统呼吸开始表现出下降趋势,甚至回到了增温之前的水平[114-115]。研究还发现在最适温度以内,NEE也会随着温度的升高而增加[116]。值得我们注意的是,Sihi等[117]在室内的培养试验则认为,相比于恒定增温,逐步增温增加了微生物量碳,同时加速了土壤异养呼吸的速率。因此,这种恒定增温的模拟试验研究结果可能会存在很大的不确定性,不利于评估气候变化情境下的真实情况。另外增温导致了土壤水分的降低,从而使得增温的效应受到水分降低的影响[13, 118],但是增水又会缓解增温所带来的负效应,所以为了更好地认识和理解未来气候变化情景对草地生态系统碳循环关键过程的影响及其机制,有必要开展长期野外逐步增温和增水耦合试验,阐明逐步增温和水分变化的修饰作用尤为重要。
此外,地下生态学研究对生态系统和地下碳循环模型的正确建立有十分重要的作用,特别是需要厘清在气候变化下草地生态系统的这些过程如何响应与反馈。草地总生物量的60- 80%左右分配在地下,而细根生长和周转及其对气候变化的响应的研究还很缺乏[48, 119]。植物根系直接控制着水分和养分的吸收,进而影响植物的生长过程,根系的周转直接影响着土壤碳库和微生物的活性[120-121]。因此,为了降低草地碳储量及其动态变化结果的不确定性,需要进一步加强光合产物的地下分配、以及细根生产和周转等地下核心过程的研究。另外,和植物地上部分物候相比,目前对于植物根系物候的研究还很匮乏[122]。已有的知识表明,根系物候和植物地上部分物候并不同步,同时根系的生长和植物地上部分的生长也存在不同步的现象[123-124],所以利用微根窗技术以及同位素标记等技术来加强根系和根系物候的观测,能够帮助我们很好地认识根系动态这一关键的碳循环过程。同时随着微生物学研究新技术的发展,认识微生物多样性及其功能基因对气候变化的反应也是理解碳循环关键机理的必然需求。
综上所述,在未来的野外试验中,可设计逐步增温和降水耦合试验平台,进一步结合并利用同位素标记技术、分子生物学技术、高通量测序技术、功能基因芯片技术和宏基因组技术以及微根窗技术等现代技术手段,从植物-土壤-为生物等生态系统不同层次,开展逐步增温对草地生态系统碳循环关键过程特别是地下生态过程(根系物候)的影响及其机理的研究。
