长白山次生杨桦林种群空间点格局及密度制约效应
2021-04-08吴兆飞范春雨张春雨赵秀海
安 璐,吴兆飞,范春雨,张春雨,赵秀海
北京林业大学国家林业和草原局森林经营工程技术研究中心, 北京 100083
植物种群的空间分布格局及其形成机制一直是生态学研究的热点问题[1],国内学者对长白山地区不同森林类型的空间分布格局开展了大量研究,但生境异质性及密度制约效应如何作用于空间格局的形成过程尚不清楚。长白山次生杨桦林中绝大多数树种的空间格局并未表现出明显的空间异质性[2],并且环境因子对种群空间变异的解释能力较低[3]。与之相反,宋厚娟等[4]发现长白山阔叶红松林中样地小环境和地理位置是导致群落结构出现显著差异的主要因素。郝珉辉等[5]也发现生境异质性与大多数物种分布格局之间都存在显著的空间相关性。目前生境异质性与林分空间分布之间的关联性还未形成较为统一的认识。因此,在开展林分空间点格局分析时,检验并及时剔除生境异质性带来的潜在影响是十分必要的。
密度制约是发生在相邻木之间的一种调节机制,使得存活个体在空间上分布更加均匀,并且聚集程度将随着径级的增加而降低[6- 7]。目前,密度制约效应对林分空间分布格局影响研究主要集中在热带森林[8- 9]和亚热带森林[10- 12],温带森林的密度制约研究相对较少。例如,长白山阔叶红松林及其次生林负密度制约对幼苗存活具有显著的效应[13-15]。阔叶红松林群落幼树及大树在一定程度上存在密度制约效应[16-17],并且负密度制约在次生杨桦林群落构建过程中发挥了重要作用[18]。
次生杨桦林属于长白山阔叶红松林演替的早期阶段,生境异质性导致的生境过滤过程对其群落结构形成具有较大影响。利用生态模拟方法控制生境异质性的潜在影响,通过空间点格局方法量化密度制约在不同生活型及不同生长阶段空间格局形成中的作用,有助于进一步深化我们对演替早期群落构建过程的认识。因此,本文以吉林长白山5.2 hm2次生杨桦林样地为研究对象,开展了如下研究:1)评估研究样地是否存在生境异质性;2)探讨在去除生境异质性大尺度影响后,树木种群是否仍然在特定尺度上存在聚集分布格局;3)采用随机标签零模型案例—对照设计的方法检验密度制约效应,并探讨密度制约与物种多度和同种聚集之间的关系。
1 试验地概况
研究实验区位于吉林省白河林业局光明林场境内,位于东经128°07.82′,北纬42°19.17′,地势平缓,平均坡度小于5°。研究区属于温带大陆性山地气候,夏季受太平洋高气压影响,冬季有西伯利亚东部及中亚反气旋的干扰,年平均气温约为-7.3—4.9℃,年平均降水为782.37—967.28 mm。由于该区域全年气温较低,降水量大而蒸发量小,森林覆盖率高,因此空气湿度非常高,年相对湿度约70%。研究区原始植被类型为阔叶红松林,主要阔叶树种包括蒙古栎(Quercusmongolica)、白桦(Betulaplatyphylla)、紫椴(Tiliaamurensis)、山杨(Populusdavidiana)、春榆(Ulmusjaponica)、色木槭(Acermono)以及水曲柳(Fraxinusmandschurica)等,针叶树种主要为红松(Pinuskoraiensis)、臭松(Abiesnephrolepis)和鱼鳞云杉(Piceajezoensis)[19]。
2 研究方法
2.1 样地建立及树种选择
课题组2005年参照CTFS(Center for Tropical Forest Science)样地建设标准(http://www/ctfs.si.edu/),在吉林省白河林业局光明林场建立了5.2 hm2(260 m×200 m)次生杨桦林固定监测样地。以 20 m×20 m 划分连续样方,对样方内胸径≥1 cm 的木本植物进行每木调查,记录物种名称、胸径、树高、坐标等信息。2017年对该次生杨桦林样地开展的复测中,共调查到23,695株木本植物个体。根据树种可达的最大高度将其分成 3 种生活型:冠层木(h≥15 m)、亚林层木(5 m≤h<15 m)和灌木(h<5 m)。对不同生活型分别界定大、中、小个体的胸径大小范围:灌木(小1 cm≤DBH<1.5 cm;中1.5 cm≤DBH<2 cm;大DBH≥2 cm),亚林层木(小1 cm≤DBH<2.5 cm;中 2.5 cm≤DBH<5 cm;大 DBH≥5 cm),冠层木(小 1 cm≤DBH<5 cm;中 5 cm≤DBH<10 cm;大 DBH≥10 cm)。为保证足够大的样本量,从所有树种中筛选出满足大、中、小个体样本量均大于40的树种,最终本文将14个常见树种作为研究对象(表1)。
2.2 点格局分析
使用双相关函数g(r) 分析不同尺度下的树木空间分布格局。双相关函数g(r) 是由Ripley′sK(r)函数推演而来的,Ripley′sK(r) 函数是以某一任意点为圆心、r为半径的圆内,期望点数与样方内点密度的比值[20]。g函数是基于成对个体间距离的关联性函数,分析以r为半径、dw为环宽的圆环内所有个体的分布。与Ripley′sK(r) 函数相比,g函数能较敏感地判别某一尺度上点的实际分布偏离期望值的程度,是评估聚集程度的重要分析方法。g(r) 值的大小代表了聚集强度的大小,当g(r)>1 时,表示聚集分布;当g(r)<1 时,表示均匀分布;当g(r) = 1 时,表示随机分布。L(r) 为Ripley′sK(r) 函数的变形,当L(r)>0 时,表示聚集分布;当L(r) < 0 时,则表示均匀分布;若L(r) = 0 则表示随机分布。
(1)
(2)
(3)
式中,A为样地面积;n为样地内林木株数;Ir(dij) 为指示函数,dij为圆心i和圆心j两点间的距离,当dij≤r时,Ir(dij) = 1,当dij>r时,Ir(dij) = 0;Wij为边缘矫正的权重;r为距离尺度。
2.3 生境异质性
生境异质性和植物间相互作用的影响都可能导致局部点密度升高,因此难以判别点密度升高的原因。生境异质性强度很难被量化,假定随着树木个体的生长,生境异质性对树木的影响成比例地增加,那么成熟大树基本稳定的分布格局就显示了生境异质性作用的结果,从而若所有成熟大树在大尺度范围上聚集(例如,在北方森林中半径 >10 m),则应表明存在异质性[20-21]。分别分析3种不同生活型的所有成熟大树的空间格局,划分标准为:冠层木 DBH≥20 cm,亚林层木 DBH≥10 cm,灌木 DBH≥3 cm。生境异质性效应能够导致树木的聚集程度增加,在模拟检验过程中L(r)函数的累积效应能增加所显示的检测值,而g(r) 则表现微弱,故选择L(r) 函数[22],并采用完全空间随机(complete spatial randomness,CSR)作为零假设模型。
为了研究树种空间格局的二阶特征(即不同尺度下林木间的相互关系),使用异质泊松分布(heterogeneous Poisson process,HP)作为零假设模型。异质泊松分布会在半径为R的圆形移动窗口内随机移动所有树木的原始位置,这种小的位移会破坏树木位置的小尺度格局,同时保持大尺度格局不变,从而可以选择性地研究小尺度上的空间格局[10]。
使用拟合优度 GOF(Goodness of Fit)测试[23]来评估实际点格局与零模型模拟的差异显著性,P值计算公式如下:
(4)
式中,u0是观测值与期望值之间的差异;rank[u0] 的值是在整个模拟格局加上观测值的等级(rank);m为模拟次数。比如,在本文中进行 199 次模拟,加上观测值共 200 个值,若对观测值的统计值大于每一个模拟值,则rank[u0]=200,p值为 0.005。
通过斯皮尔曼相关系数(Spearman′s rho statistic)来评估物种多度与同种聚集最大强度、物种多度与密度制约强度以及同种聚集最大强度和密度制约强度之间的相关性。
2.4 双变量随机标签零模型
使用双变量g(r) 函数随机标签(Random labeling,RL)零模型和案例—对照设计[10]检验种群的密度制约效应,以红松的空间分布格局为例,来说明如何用这一方法来分析红松种群的密度制约效应(图1)。以大树作为对照(格局 1),代表生境异质性的作用,小树和中树作为案例(格局 2),g21(r) 表示案例周围对照个体的分布强度,g22(r) 表示案例周围案例个体的分布强度。如果案例相对于对照没有表现出额外的格局,那么符合随机标签零模型,实际分布的值落在置信区间内,即g21(r)=g22(r)。如果在某一尺度r上g21(r)-g22(r) 的值在置信区间的下方,则表示案例的点出现在案例周围的频率高于对照的点出现在案例周围的频率,说明小树或者中树有独立于大树聚集的格局。在这种聚集的格局里,树木生长在适合的生境里,可以排除生境异质性对树木的影响。从小树到中树聚集格局的降低,表示存在密度制约效应,聚集格局降低的程度代表密度制约效应的强度。用下式来检验密度制约的统计分析:
d(r)=dj(r)-ds(r)
(5)
式中,dj(r) 是以中树为案例时g21(r)-g22(r) 在尺度r上的值,ds(r) 是以小树为案例时g21(r)-g22(r) 在尺度r上的值,从小树到中树,如果d(r)>0 表示密度制约效应存在。rs是以小树为案例时,ds(r) 低于置信区间的尺度,rj是以中树为案例时,dj(r) 低于置信区间的尺度,rthin是d(r)>0 时的尺度,即密度制约发生的尺度,rmax为密度制约强度达到最大值时的尺度,dmax表示密度制约效应的最大强度。
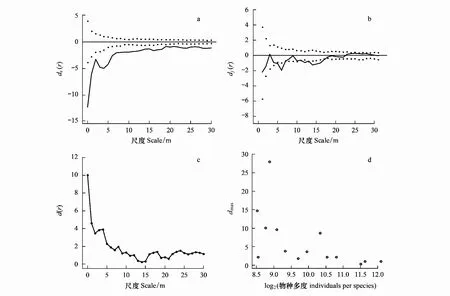
图1 同种密度制约分析Fig.1 Analysis of conspecic density dependence图a—c 为示例树种红松空间分布格局的密度制约效应;图d为所有树种的物种多度与dmax 值之间的关系,dmax 表示密度制约效应的最大强度;黑色实线为实际点格局,虚线之间代表置信区间
2.5 数据处理与分析
本文点格局分析使用 Programita Febrero 2014 软件,并采用 Microsoft Excel 2016 软件和 R 3.5.1 (http://www.r-project.org)进行数据处理和作图。通过199次Monte-Carlo 随机模拟过程构建对应点格局函数的95%置信区间。
3 结果与分析
3.1 生境异质性对空间格局的影响
各生活型成熟大树的空间格局如图2 所示,g函数和L函数偏离包迹线可以表明存在生境异质性,L函数比g函数的聚集尺度和强度更为强烈。对于冠层木和灌木的成熟个体,L函数都表现出明显的整个尺度上的聚集分布,甚至在r> 30 m 时也是如此;对于亚林层木,L函数在 20 m 处由聚集分布转为随机分布。
此外,本研究分析了每个树种成熟个体的空间格局(表 1)。L函数分析中,所有树种的成熟个体在 0—30 m 尺度范围内都表现出强烈的聚集分布。g函数分析中,各树种均为聚集分布,其中青楷槭(Acertegmentosum)、色木槭、稠李(Prunuspadus)、假色槭(Acerpseudo-sieboldianum)在个别尺度上存在随机分布。一般而言,小尺度上的聚集是植物之间相互作用引起的,而大尺度上的聚集可以解释为由生境异质性引起的[20],这充分说明生境异质性显著影响各树种的空间分布。这一发现为长白山次生杨桦林存在大尺度范围的生境异质性提供了证明,这表明在研究空间格局时,应该考虑生境异质性对结果的影响。
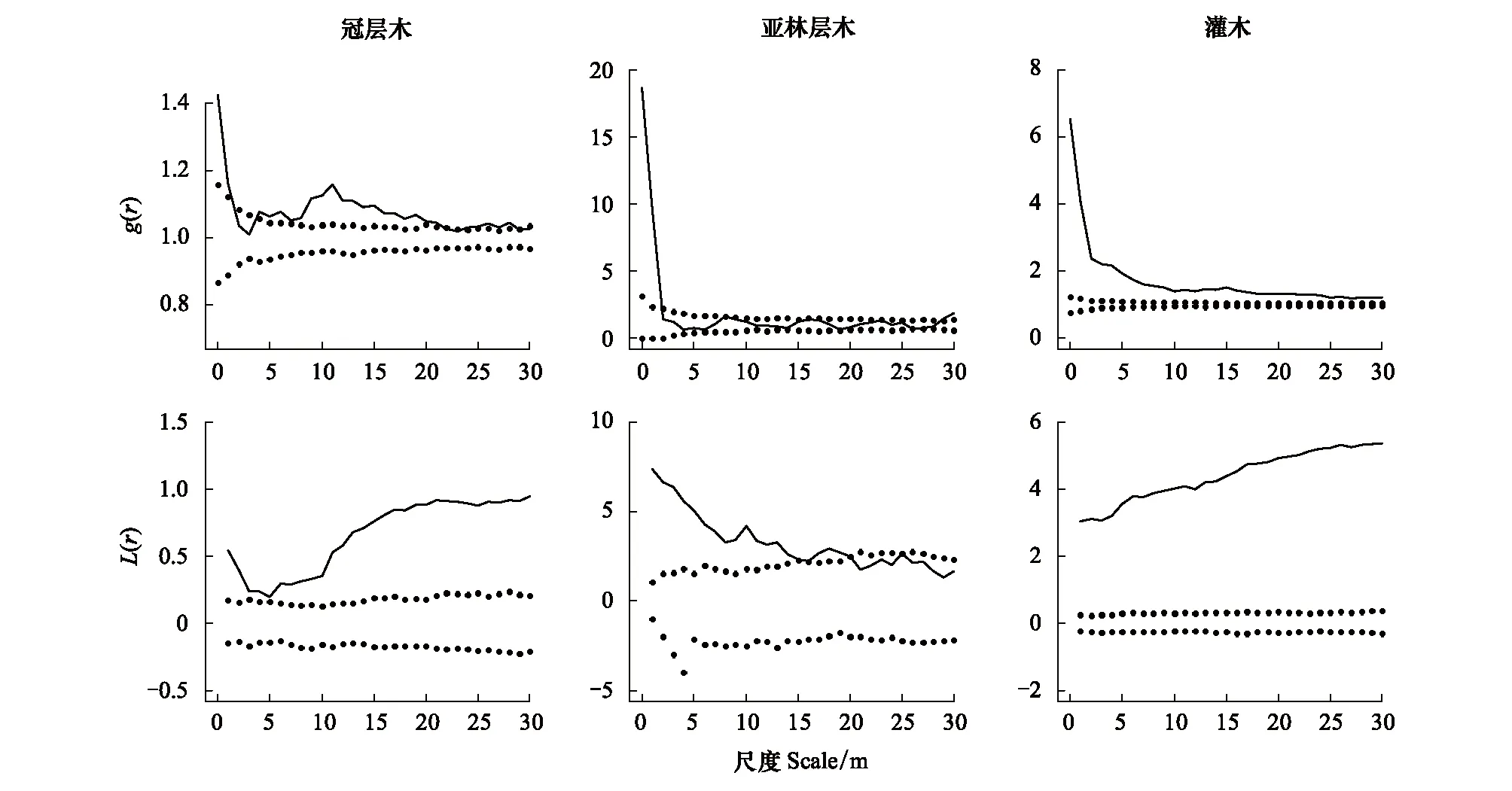
图2 所有成熟大树的点格局分析Fig.2 Analysis of point pattern of all old adult trees完全随机零模型下用双关联函数g(r) 和L(r) 函数进行点格局分析;黑色实线为实际点格局, 虚线之间代表置信区间。
3.2 去除生境异质性后的林分空间格局
与异质泊松分布零模型进行对比的 14 个树种中有 12 个树种(85.7%,除了蒙古栎和鱼鳞云杉)在小尺度上显示出显著的聚集(即GOF 检测的P值小于 0.05),其中有 5 个树种(35.7%)在个别尺度上(15—30 m 范围内)表现出均匀分布(表 1)。
同种聚集的最大强度(gmax,g(r) 在尺度上的最大值)与物种多度无相关性,而亚林层木和灌木树种的gmax值与物种多度呈显著地负相关(Spearman′s rho=-0.94,S=68,P= 0.0048)。
通过计算各尺度表现出不同空间格局的树种的百分比,评估了空间格局第二阶特征的尺度制约效应。如图3所示,聚集百分比在 0—1 m、4—6 m 的范围处达到峰值(93.3%),然后随着r的增加而急剧下降,在约 18 m 处聚集比例基本保持为 0。均匀分布在 20 m 处开始出现,这种均匀分布可能是由于零模型与小尺度上的强烈聚集相作用的结果[10]。如图3左图为假色槭的空间格局,异质泊松分布把在小尺度上聚集的点随机分布在 0—30 m 的尺度上,从而产生在大于 20 m 的距离处,包迹线高于观测值的对相关函数。
3.3 密度制约效应对分布格局的影响
采用随机标签零模型和案例—对照设计分析密度制约效应,结果如表 1 所示。从中可以看出:小树作为案例时,14 个树种均表现出相对于大树的额外聚集分布,其中臭松、红松、色木槭、紫椴、假色槭和簇毛槭(Acerbarbinerve)在整个 0—30 m 尺度上均有明显的聚集,暴马丁香(Syringareticulata)和稠李在大尺度上存在部分
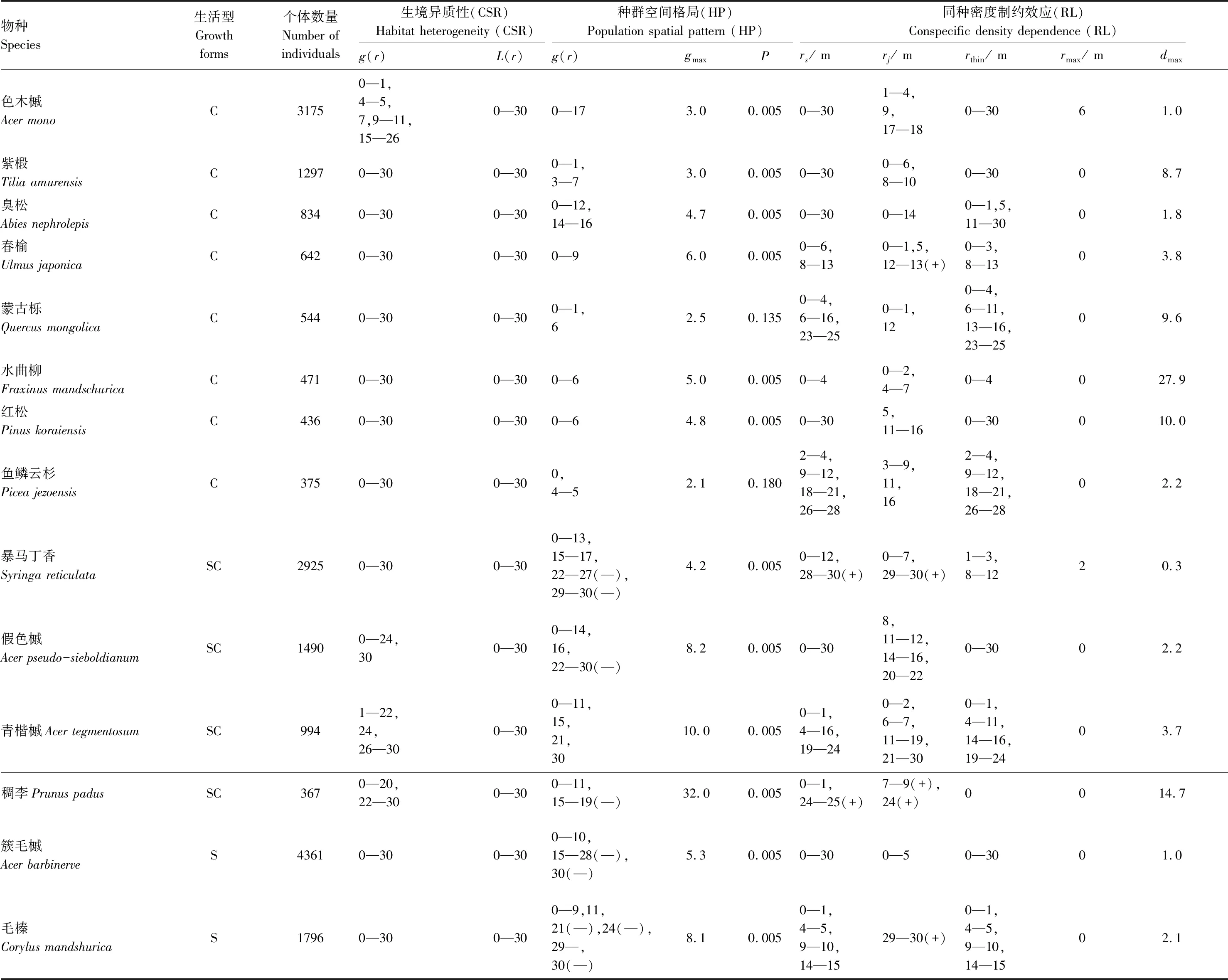
表1 分析生境异质性、种群空间格局和同种密度制约效应
均匀分布;中树作为案例时,14 个树种中有 12 个树种表现出相对于大树的额外聚集分布,4 个树种表现为部分均匀分布,分别为春榆、暴马丁香、稠李和毛榛(Corylusmandshurica)。与大树相比,从小树到中树额外聚集分布的百分比在检验尺度上明显有所下降(图4)。
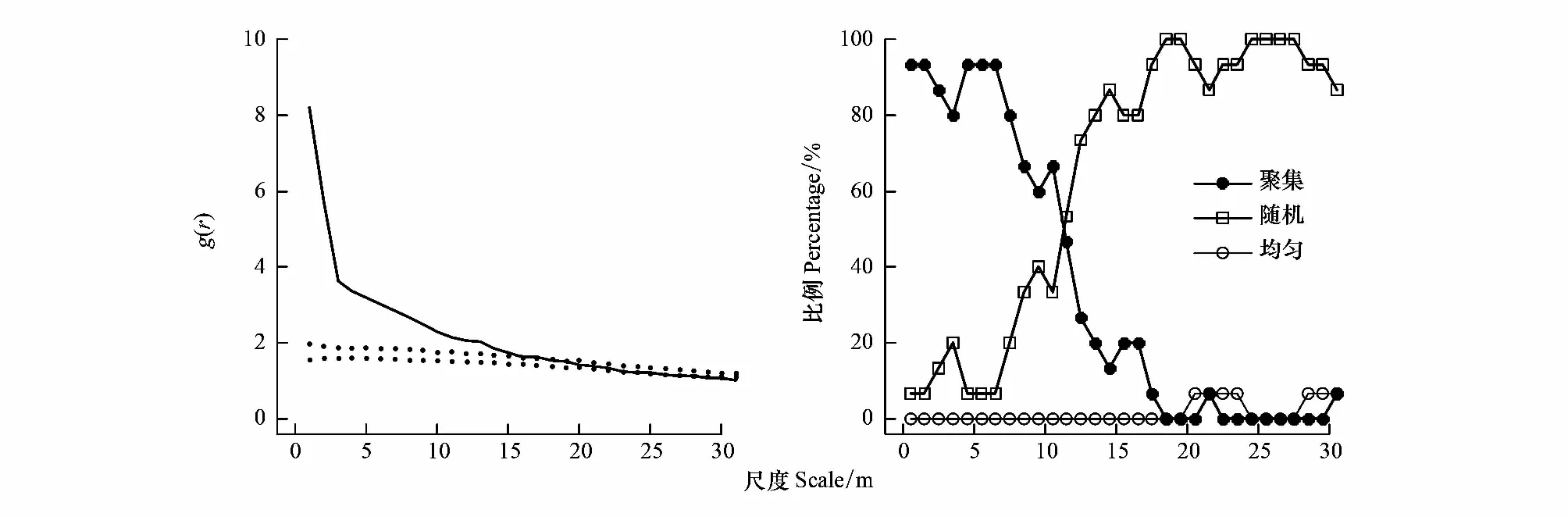
图3 去除生境异质性影响后的林木空间分布格局Fig.3 Spatial distribution pattern of trees after removing the influence of habitat heterogeneity

图4 密度制约效应的相关尺度分析Fig.4 The analysis of scale-dependence of density dependence当 g21(r)-g22(r)<0 显著时,用实心圆表示;当 g21(r)-g22(r)>0 显著时,用空心圆表示;当 g21(r)=g22(r) 时,用空心方块表示
所有 14 个常见树种的额外聚集强度从小树到中树均呈下降趋势,即均受到密度制约的影响,这表明在长白山次生杨桦林群落中林木分布格局受密度制约效应的影响十分普遍(表 1)。其中有 5 个树种(35.7%)在 0—30 m尺度上均受密度制约效应的影响,其中包括 4 个物种多度大的树种(n>1000)(色木槭、紫椴、假色槭、簇毛槭)和 1 个物种多度小的树种(n<1000)(红松)。各树种密度制约强度的最大值dmax与物种多度呈显著的负相关(Spearman′s rho=-0.90,S=863.9,P<0.0001)(图1),且与同种聚集强度最大值gmax无相关性,而亚林层木和灌木树种的dmax与gmax值为显著的正相关(Spearman′s rho=1,S=0,P<0.0001)。
随着尺度的增大,受密度制约影响的树种百分比逐渐减小。此外,密度制约强度的最大值主要在6 m以内的尺度上发生,14个树种中有13个树种在0—2 m内密度制约强度达到峰值,11个树种(78.6%)在<1 m的尺度内达到dmax,这一结果表明,密度制约效应主要发生在邻木个体之间的小尺度上(图4)。
4 讨论
4.1 种群空间分布格局
本研究中检验的树种普遍在较小尺度表现为聚集分布格局,而在较大尺度上表现为随机分布格局或部分伴随均匀分布,符合森林群落种群聚集分布的一般规律[24],在长白山次生杨桦林[25- 27]以及其他温带森林[28- 29]、亚热带森林[10- 11, 30]、热带森林[31]中都有相似的结果。然而胡艳波等[32]得到了不同的结果,他们报道了吉林蛟河阔叶红松林内树种主要呈随机分布格局,这可能主要与森林所处的演替阶段有关。
Lin等[33]的研究认为生境异质性和种子扩散限制是影响树木空间分布的主要因素。由于红松、臭松和鱼鳞云杉等针叶树种的种子较重,散落时更容易分布在母树周围,因此小径级个体更容易形成聚集分布格局。紫椴、水曲柳的种子量大,也容易分散在母树周围。此外,簇毛槭和假色槭等槭树以及毛榛都具有一定的萌蘖性,因此容易在小尺度上表现为聚集分布。随着种间竞争增强,大量小径级个体死亡,树种聚集程度下降并趋于在大尺度上随机分布。
本研究利用L(r) 函数验证了生境异质性的影响,结果表明所有 14 个常见树种均受生境异质性的强烈影响。去除生境异质性的影响有利于评估密度制约在调节种群结构中的相对重要性,例如在较大尺度上,随着径级大小的增加,聚集的减少可能是由不利生境而不是密度制约所导致的,在较小尺度上,有利生境会增加林木的聚集,从而抵消了密度制约导致的同种树木的减少[34]。Zhu等[10]用成熟树的空间格局来控制可能存在的大尺度生境异质性时,在小尺度上显示密度制约的树种比例高于在大尺度上的比例,在不考虑生境异质性的情况下,小尺度和大尺度上显示密度制约的树种比例没有差异。长白山温带次生杨桦林不同生活型或不同树种的成熟大树分布格局均表现出强烈的大尺度异质性,为了保证检测到较准确的密度制约效应,有必要考虑生境异质性的影响。
4.2 密度制约效应
生境异质性是影响种群格局发生变化的重要因子,但去除生境异质性后,密度制约仍然是调节种群结构的重要驱动机制[10]。起初对密度制约效应的研究集中在热带森林[8-9],其中Wills等[9]在巴拿马地区得到受密度制约影响的树种比例为 79.8%。后期在亚热带森林[10-12]及温带森林[13-15,35- 36]的研究也逐渐增多,受密度制约影响的树种比例Zhu等[10]在亚热带森林得到83.0%,Lambers等[36]在温带落叶林发现等同于热带森林。在长白山阔叶红松林中,Kuang等[16]和Piao等[17]证明只有少数物种受密度制约的影响,而在长白山次生杨桦林开展密度制约的研究相对较少,且常见于幼苗水平的分析。范秀华[18]和肖翠等[37]利用广义线性混合模型以幼苗的存活状态为变量证实了负密度制约效应在长白山次生杨桦林中存在,王蕾等[38]利用一般线性模型以幼苗密度为变量得到了同样的结果。在本研究中,利用生态模拟方法控制生境异质性的潜在影响,以空间点格局的方法量化密度制约在各树种不同生活型及不同生长阶段空间格局形成中的作用,结果表明,14个常见树种受密度制约影响的比例(100%)与其他森林中的研究结果相比更高,这可能是由于该温带森林中满足样本量的树种相对于热带或亚热带森林更少,且次生杨桦林的森林群落演替阶段相比阔叶红松林群落更不稳定。总之,在后期不同纬向带森林的研究中,普遍均证明了密度制约效应的存在,且对调控物种共存机制有重要作用,那么密度制约并不是热带森林的特有效应,密度制约并不能解释为什么热带森林的物种比其他纬向带森林的物种更丰富,除非热带森林的密度制约强度更强。
对物种多度与密度制约效应关系的研究也是森林生态学研究的热点[8],已有研究证明了密度制约效应与物种多度之间为负相关关系[39-41]。在本研究中,物种多度小的树种其密度制约效应的最大强度(dmax)比物种多度大的树种更强(图1),此外,可以观察到物种多度小的树种其同种聚集最大强度(gmax)相比物种多度大的树种更强(表1),这主要体现在亚林层木和灌木树种上,冠层木树种中蒙古栎和鱼鳞云杉等树种没有体现,这一结果与Zhu等[10]的研究结果基本保持一致。
5 小结
本研究通过对长白山次生杨桦林空间分布格局和密度制约效应分析,发现在空间格局的研究中消除生境异质性的影响至关重要。同时,密度制约是调节温带森林树种种群结构的一种重要机制,密度制约强度与物种多度为负相关,与亚林层木和灌木树种的同种聚集最大强度为正相关。本研究为温带森林林分空间格局和密度制约效应的研究提供了理论支撑,对长白山地区次生杨桦林的群落结构构建和可持续经营具有重要意义。
