植物功能群去除对高寒草甸群落结构、多样性及生产力的影响
2021-04-08杨振安何奕忻彭长辉
姜 林,胡 骥,杨振安,詹 伟,赵 川,朱 单,何奕忻,陈 槐,3,*,彭长辉,5
1 西北农林科技大学林学院生态预测与全球变化研究中心, 杨凌 712100 2 中国科学院成都生物研究所山地生态恢复与生物资源利用重点实验室, 成都 610041 3 中国科学院青藏高原地球科学卓越创新中心, 北京 100101 4 西华师范大学生命科学学院, 南充 637002 5 魁北克大学蒙特利尔分校环境科学研究所,蒙特利尔, 加拿大 C3H3P8
群落中物种的丧失所引起的生物多样性下降对于生态系统过程和功能的影响存在多种模式,但多数研究结果显示的负效应导致了对于生物多样性丧失的担忧[1- 2]。在我国青藏高原广泛分布的高寒草甸群落中,由于人为干扰、气候变化等因素所导致的生物多样性减小普遍存在。例如,过度放牧在长时间尺度上会引起优势禾草类群的消失[3],围封禁牧可能抑制低矮杂草的生长发育[4],模拟增温对植被地上部所施加的高温胁迫会导致显著的物种丰富度下降[5],施用氮素肥料导致的生态位分化的加强会引起低竞争优势物种的丧失[6]。上述物种多样性的损失往往具有非随机性[7-8],主要来源于物种对干扰的特定响应,如过牧导致的禾草损失主要来源于牲畜的选择性采食[9];或者种间竞争关系的改变,如施肥通过增大禾草竞争优势导致杂草物种的排除[10]。由此导致的群落物种构成的变化对系统功能(如生产力)具有显著影响,并且主要取决于群落中的优势物种或功能群[11-12]。目前,群落物种损失对系统过程和功能的影响效应及作用机制尚不清晰,有必要对此问题进行深入分析。
物种或功能群的去除是近年来群落生态学中应用较多的方法,通过对特定物种或功能群的去除,能够得到更接近实际物种损失背景下的系统过程和功能的响应[13]。基于青藏高原的高寒草甸,科研人员已经开展了若干植物去除试验来探讨物种损失的效应。Li等[14]在植物功能群去除试验中发现,优势禾草的丧失会显著影响群落生产力,而杂草功能群的丧失使物种多样性的损失更大。而该区域禾草草甸和小嵩草草甸中进行的去除试验表明,优势物种的损失对群落生产力无显著影响[15]。田红等[16]的去除试验结果表明,群落物种多样性与生产力之间存在正相关性。Zhou等[17]通过去除不同的植物类群发现,物种的丧失通常会降低土壤有机质和营养元素含量,而物种多样性的减小会导致土壤有效磷含量下降。Liu等[18]通过去除特定植物类群构建了多样性梯度,其结果表明,功能性状系统发育多样性和株高的组合比单一多样性指数对生产力变异的解释更大。牲畜放牧是目前高寒草甸面临的主要干扰模式,长期的过牧会导致高寒草甸由禾草占优的群落向以莎草、杂草占优群落逆向演替,期间伴随有物种的损失过程[3]。基于此,本研究在青藏高原高寒草甸群落中构建了植物功能群的梯度去除试验,试图独立的模拟上述逆向退化过程中的物种损失模式。通过对功能群损失下群落结构、组成、多样性和生产力的监测和分析,探讨高寒草甸群落中不同植物功能群的竞争关系,以及群落多样性对生产力的决定模式。
1 研究方法
1.1 试验样地概况
本研究选取了位于青藏高原东缘若尔盖区域的高寒草甸群落作为试验对象。研究样地地理坐标为32°57′59.89″ N、102°37′10.30″ E,海拔高度3475 m,在行政区划上归属四川省阿坝藏族羌族自治州红原县。区域气候为典型大陆性高原寒温带湿润-半湿润季风气候,寒冷而潮湿。试验样地临近气象站(红原站,32°47′34.92″ N、102°32′59.04″ E)的监测数据(国家气象科学数据共享服务平台,http://data.cma.cn/)显示,该区域多年(1961—2016)的年均气温为1.6(0.5—2.8)℃,年均降水量为751.5(508.1—996.3)mm,年均气压仅为665.9(664.5—667)hPa,冬半年(11月至次年4月)寒冷干燥,夏半年(5月—10月)温暖湿润,其降水量占全年的86(76—92)%。
试验样地位于河流一级阶地,临近洪积扇边缘,地势平坦。样地土壤类型为(高原)草甸土亚类,归属土壤系统分类的普通草毡寒冻雏形土亚类[19-20]。剖面调查显示,土壤有效土层深度为110 cm,包括腐殖质层(0—37 cm)、过渡层(37—60 cm)和淀积层(60—110 cm),成土母质为古河道冲积物和洪积物。试验对象亚高山草甸归属川西北高原灌丛、草甸地带下的川西北高原亚高山灌丛、草甸亚带[21],曾长期被作为牦牛等家畜的冬季草场使用,于2014年4月开始进行围封禁牧处理。对象高寒草甸群落的主要组成类群包括:禾本科(Gramineae)、莎草科(Cyperaceae)、菊科(Compositae)、毛茛科(Ranunculaceae)、蔷薇科(Rosaceae)、豆科(Leguminosae)、唇形科(Labiatae)、大戟科(Euphorbiaceae)、龙胆科(Gentianaceae)、玄参科(Scrophulariaceae)和伞形科(Umbelliferae)等。
1.2 试验设计
将高寒草甸群落中的所有植物种依据生活型划分为以下3个功能群(plant functional group, PFG):禾草功能群(Grass)、莎草功能群(Sedge)和杂草功能群(Forb)。试验设置以下6个处理:不去除任何功能群(N,对照)、去除禾草功能群(G)、去除禾草和莎草功能群(GS)、去除所有功能群(A)。为了检验去除操作所引起的地上生物量损失的潜在影响[22],设置随机去除25%(Q)和50%(H)的群落地上生物量2个处理。试验采用随机区组设计,共设置3个样区作为重复,大小为11 m×11 m。每个样区内设置6个1 m×1 m的样方,随机性对应6个处理,共18个试验样方,样方间隔2 m以减小边际效应。去除处理在2014、2015年的07月中旬进行,去除功能群时以剪刀齐地表剪去样方中目标物种的地上部分并移出,随机去除生物量时以10 cm×10 cm方格为单位,通过去除一定数量的植物地上部分实现。为监测群落特征的时间变化,我们分别于2014、2015年的7月下旬在试验样地毗邻区域进行了相同的调查,分别以C1、C2表示(对照,n=5)。
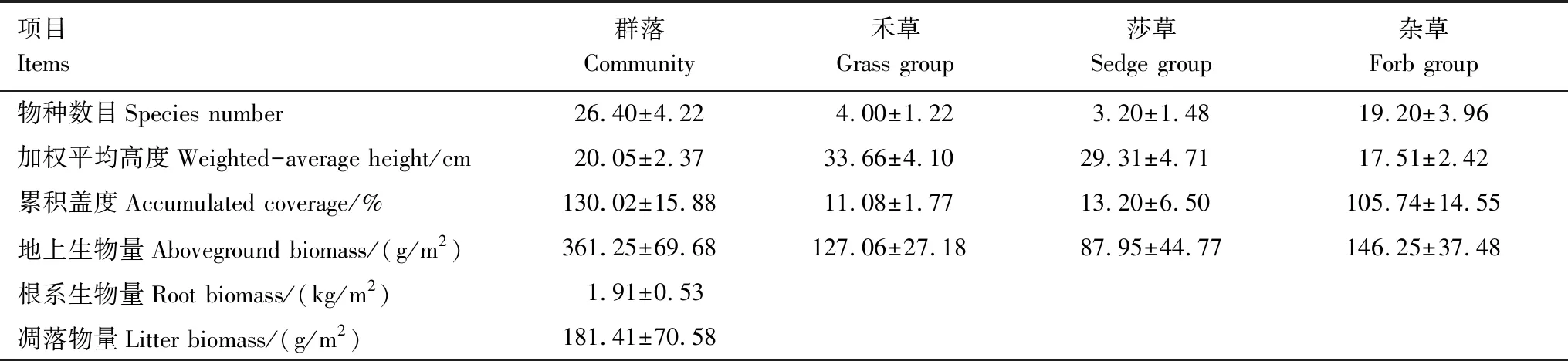
表1 试验对象高寒草甸群落及功能群特征(2014年)
1.3 样品采集与指标测试
植物群落的调查和样品采集于2016年的7月中下旬进行。将50 cm×50 cm的钢制框架置于试验样方的居中位置,框架以细线均分成10×10个方格。调查时,记录框架内出现的所有物种,通过多人目测取平均值的方法估算各物种盖度,对每个物种随机选择其中5株测量其株高并取平均值作为物种高度。完成群落调查后,按照禾草、莎草和杂草3个功能群用剪刀齐地表剪取植物地上部,并收集框架内的凋落物。用直径5 cm不锈钢土钻钻取0—10 cm深度的表层土壤并将完整土柱带回,将土柱置于孔径1 mm的筛中用流动水冲洗至无土壤颗粒作为根系样品。将上述植物样品分别置于对应纸质样品袋,于电热鼓风干燥箱(DHG- 9246A,精宏,上海)中杀青(85 ℃,0.5 h)并烘干至恒重(65 ℃)。以电子天平(BSA224S, Sartorius, Germany)称取植物样品干重,精确至0.001 g。
本研究中,我们以植物高度和盖度来表征群落结构,这也是针对草地群落结构特征研究中常用的指标体系[23]。计算过程中,群落或功能群平均高度为所包含物种平均高度的加权平均值(以各物种盖度为权重),群落或功能群累积盖度为所包含物种盖度的累积值。参照前人对物种多样性指数的介绍与讨论[24- 26],我们选取了以下四类物种多样性指数,包括Patrick物种丰富度指数(r, 丰富度)、Shannon-Wiener物种多样性指数(h,SW多样性)、Simpson物种优势度指数(d, 优势度)和Pielou物种均匀度指数(j, 均匀度)。各多样性指数的计算公式如下所述:
Patrick丰富度指数:
r=S
(1)
Shannon-Wiener多样性指数:
(2)
Simpson优势度指数:
(3)
Pielou均匀度指数:
(4)
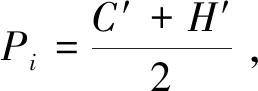
1.4 数据统计及分析
应用单因素方差分析(one-way ANOVA)检验去除处理对群落结构、多样性和生产力等指标的影响,并应用Duncan检验法进行多重比较。相关性分析通过计算指标间的Pearson线性相关系数获得。显著性水平设置为P<0.05。统计分析在SPSS 22.0软件中进行。
2 结果分析
2.1 群落结构和功能群组成对去除处理的响应
由图1可见,群落累积盖度在不同处理间没有显著差异,而去除处理对群落加权平均高度具有显著影响。其中,对照群落(C1、C2和N)的高度伴随试验进行显著增加,而去除操作不同程度的降低了群落高度。其中,去除全部功能群(A)的降幅最大且显著。除莎草外,禾草和杂草功能群的优势比均受到显著影响。在试验周期内,对照群落中禾草的优势比显著增加而杂草优势比显著降低。相比于对照(N),随机去除25%生物量(Q)显著降低了禾草优势比;同时,去除禾草和莎草(GS)导致杂草优势比显著增大。

图1 不同处理的群落盖度、高度和功能群综合优势比Fig.1 Community coverage, height and SDR of PFG under different removal treatments图中柱形表示平均值,误差线表示标准差。柱形上部不同小写字母表示处理间差异显著(P<0.05);C1、C2分别表示2014、2015年样地毗邻区域的背景值,N表示不去除、G表示去除禾草,GS表示去除禾草和莎草,Q表示随机去除25%生物量,H表示随机去除50%生物量,A表示去除所有功能群;SDR: 综合优势比Summed dominance ratio;PFG: 功能群Plant functional group
2.2 群落多样性对去除处理的响应
四种物种多样性指数中,仅有物种均匀度指数受到了去除处理的显著影响(图2)。随着试验进行,对照群落中物种均匀度指数逐渐下降,与物种SW多样性指数的变化趋势相同而与物种丰富度指数、优势度指数的变化趋势相反,但后三者的变化趋势均不显著。相比于对照(N),植物功能群的去除普遍增加了物种均匀度。其中,去除禾草和莎草(GS)、去除全部植物(A)和随机去除25%生物量(Q)处理的增加显著。植物功能群SW多样性和优势度指数均受到去除处理的显著影响。随着试验进行,对照群落中功能群SW多样性增加而优势度减小。相比于对照(N),仅去除禾草和莎草(GS)显著降低了功能群SW多样性。去除禾草、莎草群落(GS)的功能群优势度显著高于去除禾草(G)和随机去除50%生物量群落(H)。
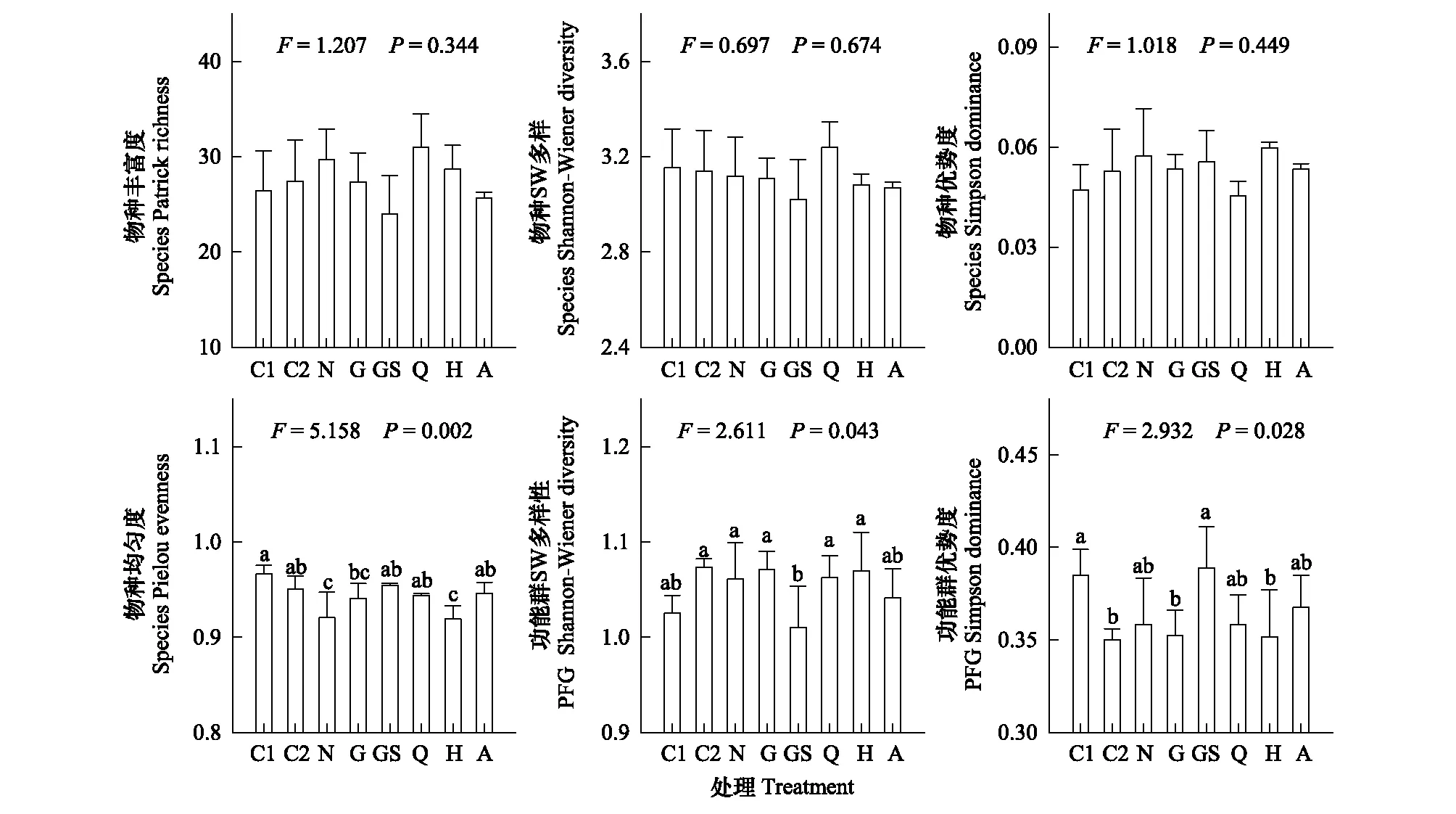
图2 不同处理的物种和功能群多样性指数Fig.2 Species and PFG diversity indices under different removal treatments

表2 不同植物功能群生物量比例及综合优势比之间的相关系数
2.3 群落及功能群生物量对去除处理的响应
由图3可见,对照群落地上生物量、根系生物量和凋落物量随着试验进行均增加,但后两者没有显著差异。仅去除全部功能群(A)显著降低了群落地上生物量。去除处理均显著降低了凋落物量,且去除全部植物(A)的降幅最大。对照群落中禾草生物量显著增加,而莎草、杂草生物量均先增大后减小,但差异不显著。功能群去除均显著减小了禾草生物量,但相互之间无显著差异。试验处理对莎草、杂草生物量的影响均不显著。总体上,禾草、莎草和杂草生物量比例的变化趋势与其生物量类似。对照群落中,禾草生物量比例逐渐增大,莎草生物量比例先增大后减小,而杂草生物量比例持续减小。

表3 群落地上生物量与功能群生物量及其比例之间的相关系数
3 讨论
针对高寒草甸的研究表明,围封禁牧、施肥和(模拟)增温等措施能够增加禾草、莎草的丰度,同时降低杂草的丰度,并提高群落生产力[4,6,28]。这表明高寒草甸群落中不同生活型植物类群的共存具有竞争性,可能主要源于它们对冠层光合有效辐射和土壤养分的获取以及拓植能力的差异[29-30]。本研究发现,禾草、莎草和杂草功能群的优势比之间均存在显著的线性负相关,并且禾草、莎草的生物量比例与杂草生物量比例之间也存在极显著的线性负相关(表2),表明3种植物功能群之间确存在竞争关系。叶面积和叶位相对较大的禾草类群在不受干扰的高寒草甸中常处于竞争优势地位[31],因而随着试验进行,不去除处理下(C1、C2和N)禾草的优势比显著增加而杂草的优势比显著下降,且二者生物量比例也具有类似的趋势。相关性分析的结果还显示,禾草生物量与群落地上生物量之间存在极显著的线性正相关性(表3),表明高寒草甸生产力主要取决于优势禾草,符合质量比假说[12]。植物地上部对动物采食常表现出补偿性生长[32],而连续的损失则会降低其同化作用,减小对繁殖器官(种子、地下根)的投入,导致种群再生受限。因此,包含去除禾草的处理均显著减小了其生物量,而包含去除莎草和杂草的处理也减小了二者的生物量(图3)。此外,功能群去除在抑制自身再生的同时会影响植物类群间的竞争关系[15]。本研究中,禾草和莎草的同步去除(GS)弱化了二者的竞争效应,显著增加了杂草功能群的优势比,使其生物量达到最高(图3);但在去除全部功能群时,禾草仍具有最大的竞争优势,并对其他功能群产生抑制(图1)。
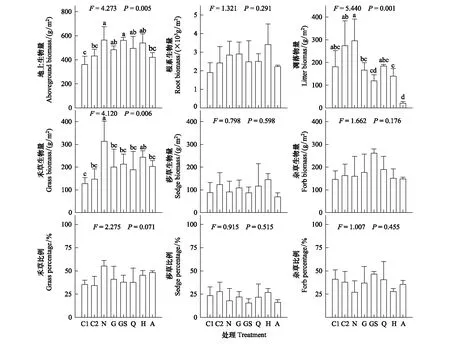
图3 不同去除处理下的群落及功能群生物量Fig.3 Biomass production of community and PFG under different removal treatment
同时,植物类群间的竞争性在受到弱化时还会导致其生长状况的补偿性[2,22]。本研究中,去除禾草和莎草降低了二者的生物量,但杂草生物量的增加补偿了群落地上生物量的损失,因而G、GS处理下总生物量与N处理无显著差异(图3)。杂草类群的类似补偿作用在McLaren and Turkington的研究结果中也存在[2],但后者的补偿较慢,较快的补偿常存在于气候相对温和的草地群落[33]。这种差异一方面可能来自于高寒草甸群落中较强的竞争关系,另一方面可能受不同的去除强度的影响[34]。当去除所有功能群(A)时,群落地上生物量显著减小,同时根系生物量也呈现减小的趋势,该结果类似于过度放牧所导致的高寒草甸生产力退化。这表明,植物地上部的连续性、高强度损失能够抑制其生长过程,从而引起系统生产力的下降[9]。此外,植物物种或者功能群的去除往往会导致直接的物质损失[9]。本研究中,群落凋落物量随着去除生物量的增多呈现显著的下降趋势,而与所去除的植物功能群类型无关(图3)。凋落物的归还作用能够直接影响有效态土壤营养元素的含量,因而长时间尺度上植物功能群的损失可能会对土壤养分具有显著地负效应[35]。相对的,高寒草甸的封育能够显著提高群落生物量和凋落物量,增加物质累积,提高系统的有效养分含量[36-37]。
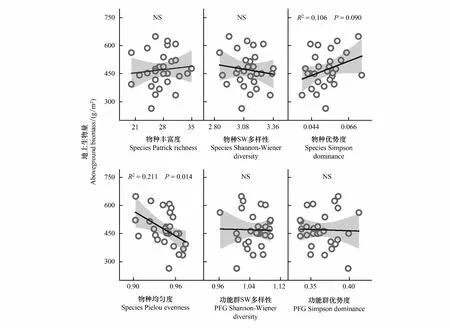
图4 群落地上部生物量与物种、功能群多样性之间的线性回归关系Fig.4 Linear regression between community aboveground biomass and diveristy indices of species and PFGNS表示回归关系不显著(P<0.05)
封育对植物物种多样性具有明显影响,同时取决于物种构成、放牧历史、环境因素等[38]。本研究中,对照群落(C1、C2和N)的物种均匀度和SW多样性逐渐减小,优势度逐渐增大。该趋势应源于封育下优势物种的竞争优势抑制了其他物种的生长发育,类似于中度干扰假说的描述[39],而丰富度的增大可能是由于中生环境中较多偶见种的存在[40]。相反的,禾草和莎草功能群的去除(G、GS)所导致的物种SW多样性的减小和均匀度的增大则源于竞争优势的弱化。上述变化趋势表明,高寒草甸植物类群间的竞争关系在一定程度上决定了物种的多样性动态,而该竞争关系同时也影响了功能群的多样性。杂草优势比在封育时的减小和去除禾草、莎草时的增大导致了功能群SW多样性和优势度相反变化。现有研究中,物种多样性与群落生产力之间常呈现正相关,少数研究发现了负相关或单峰曲线关系[41]。本研究中,群落地上生物量仅与物种均匀度指数呈显著的线性负相关,而与物种优势度指数呈不显著的线性正相关(图4),类似的关联性在放牧干扰下的高寒草甸中也存在[42-43]。这一关系可能源于优势禾草对群落生产力的较大贡献,同时也受到群落物种库和去除强度的限制[44]。因此,群落中物种的构成对生产力具有决定作用,可以作为物种多样性-生产力关系的解释[45-46]。此外,本研究中随机去除生物量与去除特定功能群所产生的影响不一致。例如,随机去除50%生物量(H)时,杂草地上部的损失导致其优势比较低,而去除禾草和莎草(GS)导致的生物量损失相当(59.5%),杂草优势比却最高(图1);类似的差异也出现在物种均匀度和功能群多样性指数的响应过程中(图2)。该结果进一步说明,高寒草甸中植物类群的特征和相互关系决定着群落对不同干扰的响应模式[22]。
4 结论
通过去除特定的植物功能群,本研究模拟了过牧干扰下高寒草甸逆向演替中的物种损失,分析了群落结构、多样性和生产力的响应。功能群的去除对群落的高度、物种均匀度和功能群多样性产生了显著影响,主要源于功能群之间竞争关系(优势)的改变。同时,功能群去除所导致的物质损失降低了群落地上生物量,尤其是禾草生物量并显著减小了凋落物量的归还。对于局域尺度的高寒草甸,物种或者功能群之间的相对优势在一定程度上决定了群落的组成,因而在群落构建及其对干扰的响应中具有重要作用。同时,物种或功能群损失的影响需要在更长时间尺度上进行连续监测,以明确各种响应的稳定性。
