岩溶与非岩溶区典型植物最适光合模型和光合特征研究
2021-04-06倪隆康何文黄科朝顾大形黄玉清
倪隆康, 何文, 黄科朝, 顾大形*, 黄玉清
岩溶与非岩溶区典型植物最适光合模型和光合特征研究
倪隆康1, 何文1, 黄科朝1, 顾大形1*, 黄玉清2*
(1. 广西喀斯特植物保育与恢复生态学重点试验室,广西壮族自治区中国科学院广西植物研究所,广西 桂林 541006;2. 北部湾环境演变与资源利用教育部重点实验室,广西地表过程与智能模拟重点实验室,南宁师范大学,南宁 530001)
为探讨不同生境典型植物光合特征对主要环境因子的响应,选择Ethier & Livingston、Ellsworth和Sharkey等3种光合生化模型拟合我国西南岩溶与非岩溶区8种典型植物的A-Ci曲线。结果表明,用Ethier & Livingston模型拟合的A-Ci曲线要优于Ellsworth模型和Sharkey模型,拟合方程的决定系数(2)为0.98~0.99 (<0.01)。从Ethier & Livingston模型拟合的Vcmax和Jmax来看,岩溶区和非岩溶区乡土植物间的差异不显著,但都显著低于外来速生植物尾巨桉(×)。岩溶区和非岩溶区乡土植物的光合特征参数(Vcmax,25和Jmax,25)均与空气温度和土壤水分含量呈显著相关(<0.05)。除非岩溶区乡土植物的Jmax,25外,所有植物的光合特征参数与土壤水分拟合方程的2均低于其与环境温度的,表明环境温度对典型植物Vcmax,25和Jmax,25的影响强于土壤含水量。
岩溶;光合生化模型;光合特征;环境因子
植物通过光合作用生产有机物并储存能量,进而驱动生态系统的物质循环和能量流动。植物光合过程的关键特征参数,如最大羧化速率(Vcmax)、最大电子传递速率(Jmax),是植被气体交换生理过程模型的重要驱动参数,其精度和环境响应特征对植被生产力估算及其气候变化响应格局模拟具有重要影响。因此植物光合特征参数是了解植被生态功能、预测植被在气候变化下群落演替的关键依据[1]。不同植物间的资源利用能力、环境适应能力等的差异导致物种间光合能力及其环境响应差别较大[2]。除种间因素外,光合特征拟合模型的选择对植物光合能力估算也有重要影响[3]。
CO2作为光合作用的主要碳源,是光合作用的主要限制因子[4]。通过拟合光合-二氧化碳响应曲线(A-Ci曲线)可以估算植物的Pmax、Vcmax、Jmax等一系列表征植物碳同化能力的特征参数。常用的A-Ci曲线拟合模型有经验模型和生化模型。1980年Farquhar等建立的C3植物电子传递和酶动力学生化模型, 极大地推动了植物光合系统对CO2浓度变化响应机制和估算模型的研究[5]。随后,Ethier等在Farquhar模型的基础上增加了二氧化碳导度以提高Vcmax的估算精度[6],该模型可拟合出如Rp、Rd、CO2补偿点和饱和点等参数,又可直接算出Vcmax和Jmax。Ellsworth模型用于研究不同物种间Vcmax和Jmax的差异及其对温度的依赖程度,但无法计算其他生理参数[7]。Sharkey模型作为广泛应用的生化模型,充分考虑A-Ci曲线3个阶段[5]对拟合结果的影响,即核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)、1,5-二磷酸核酮糖(RuBP_regen)、三碳糖磷酸利用率(TPU)限制阶段[8]。有研究表明Rubisco酶羧化能力、光合电子传递能力和CO2的扩散过程均随温度的变化而改变[9]。目前,国内对该领域的相关研究还相对较少,对各生化模型的拟合差异缺乏深入的研究和系统的认识。
岩溶区面积约占我国国土面积的三分之一,也是我国主要的生态脆弱区域[10]。岩溶强烈发育地区大量的出露裸岩和特殊的水土二元结构导致岩溶区与同区域的非岩溶区迥异的生境条件。生境的差异性可能对植物的碳同化过程产生重要影响。对岩溶和非岩溶区典型植物选择合适的拟合模型估算其光合生理参数并据此判断植物光合特征的环境响应特征,对理解植物的生境适应机制和植被气体交换过程模拟具有重要作用。目前对A-Ci曲线拟合模型的研究材料多集中在树木幼苗或离体测量, 对成熟森林树木的原位研究数据则相对有限,植物幼苗期的生理指标推广到其它生育时期可能会导致较大误差[11]。因此,本文以广西岩溶和非岩溶区典型植物为研究对象,利用Li-6400便携式光合仪野外原位测量成熟植株冠层叶片光合速率对CO2的响应过程,筛选拟合典型植物光合特征的最适生化模型,并分析不同生境下典型植物光合特征及其对主要环境因子的响应,为岩溶和非岩溶区广泛分布的我国西南地区森林碳同化过程的模拟研究提供数据支撑。
1 材料和方法
1.1 材料
广西桂林是典型的西南岩溶和非岩溶地貌分布区,为开展复杂地貌条件下植被研究提供了条件。本研究依托会仙岩溶生态试验站(25°06′03″ N,110°13′46″ E)和处于非岩溶区的桂林园林植物园生态定位研究站(25°01′46″ N,110°17′03″ E)、黄冕桉树人工林生态站(24°29′02″ N,109°45′13″ E)完成, 3个实验观测站的气候条件相同。选取典型亚热带岩溶植物九龙藤()、雀梅藤()、柞木()、马甲子()和非岩溶植物尾巨桉(,我国南方主要外来人工造林树种)以及亚热带典型地带性植物黧蒴锥()、锥栗()、荷木()为研究对象。
1.2 方法
环境因素测量 每个实验站点均建有观测塔作为冠层实验平台并安装有冠层气象观测系统。环境温度运用Li-6400 (Li-COR Inc, Lincoln, USA)便携式光合作用仪的温度传感器测量。土壤体积含水率利用经过校正的土壤水分传感器(SM150, Delta- T Devices Ltd, UK)在30 cm深度的土壤进行连续测量,用数据采集仪(CR1000, Campbell Scientific Inc. USA)记录每10 min的平均值。
光响应曲线测量 利用Li-6400便携式光合作用仪(Li-COR Inc,Lincoln,USA), 在晴朗无云的早晨9:00-10:00开始测量,叶片经过自然光的充分诱导,再利用Li-6400自带光源对被测叶片进行活化15 min,活化光强设置为1000mol/(m2·s),以确保植物完全活化。测量时采用仪器红蓝光源控制光合有效辐射强度,光强梯度依次为1 500、1 000、800、500、300、200、150、100、50、20、0mol/(m2·s),流速500mol/s,CO2浓度为400mol/mol,采用自动测量程序进行测量,最小等待时间120 s,最长等待时间200 s。光响应曲线只进行单次测量,主要目的是利用光响应曲线拟合的饱和光强进行A-Ci曲线测量的参数设置。饱和光强等参数的拟合均用经验模型计算得出[12]。非岩溶区植物尾巨桉、黧蒴锥、锥栗和荷木的饱和光强分别为1 600~1 900、800~900、900~1 000和700~800mol/(m2·s),岩溶区植物九龙藤、雀梅藤、柞木和马甲子分别为1 000~1 100、1 000~1 200、900~1 200和1 000~ 1 200mol/(m2·s)。
二氧化碳响应曲线测量 选取长势一致的冠层阳生叶片进行A-Ci曲线的测量,每种植物重复6次。根据光响应曲线拟合得到的饱和光强设置相应植物的光合有效辐射,CO2浓度梯度依次为50、100、150、200、300、400、600、800、1 000、1 200、1 600、2 000mol/mol。流速500mol/s,最小等待时间60 s,最长等待时间300 s。Vcmax和Jmax等指标均通过生化模型拟合软件计算得出[6–8]。
1.3 数据分析
试验数据测量于2019年1月至11月,逢单月选择晴朗无云的天气集中对成年植株冠层的成熟健康叶片进行光合-光响应曲线的原位测量,并计算饱和光强。随后选取6株长势一致的成年植株,在饱和光强下分别进行A-Ci响应曲线的原位测量,同时记录温度、湿度、光照等气象数据。采用3种生化模型分别拟合各植物的Vcmax,25和Jmax,25,并对不同模型的拟合精度(决定系数2、均方根误差RMSE、值)进行比较。随后利用筛选出的最优拟合模型对本研究植物的A-Ci曲线进行拟合。用单因素方差分析比较生长旺季不同植物的光合能力差异,用线性回归分析不同环境条件下植物光合性状与环境因子之间的关系, 各光合特征参数的分析结果用Excel和Sigmaplot 12.5整理和作图,用SPSS 20.0进行数据分析。
1.4 拟合模型
Farquhar模型是光合生化模型的基础,已被广泛应用于光合生化特征参数的拟合[13–15]。本文选择基于Farquhar模型的3个改进模型用于岩溶和非岩溶地区典型植物的最适光合模型的筛选。
(1) Ethier & Livingston模型[6,16]

式中,*为CO2的补偿点,O为叶内O2浓度,R是暗呼吸速率,K是Rubisco有限羧化反应的Michaelis-Menten半饱和常数,0是Rubisco限氧反应的Michaelis-Menten半饱和常数,V为最大羧化速率,S为二磷酸核酮糖羧化酶特异性因子。
式中,Ф是光响应曲线电子传递速率的初始斜率,I为PFD吸收,θ是光响应电子传递曲率因子,是电子传递速率,J为最大电子传递速率。
(2) Ellsworth模型[7]
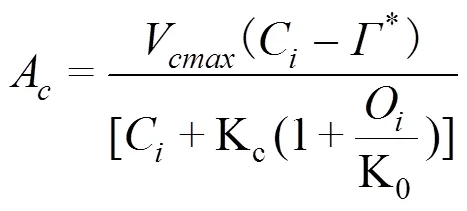
式中,C和O分别是胞间CO2和O2的浓度,*是CO2补偿点,Kc和K0均为常数,是光响应曲线的曲率(取0.90),是电子传递量子产率(固定值在0.3 mol/mol,C3植物平均光合量子产量为0.093, 叶片的吸收率为0.8。这些参数值对J的估算只有轻微的影响)[7]。
(3) Sharkey模型[8]

式中,C是二氧化碳分压的二磷酸核酮糖羧化酶,是二磷酸核酮糖处的分压,饱和光强下的为max。
2 结果和分析
2.1 不同模型的拟合结果比较
利用不同生化模型对8种岩溶与非岩溶典型植物拟合生长旺季(7月份)的A-Ci曲线见图1,在CO2浓度低于400mol/mol时,Ethier & Livingston模型和Ellsworth模型的拟合点与实测值基本一致, Sharkey模型的拟合点则明显偏离实测曲线。CO2浓度约为900mol/mol时,所有模型的拟合结果趋于一致,并接近实测值。随着CO2浓度升高,3个模型的拟合曲线逐渐与实测值偏离,其中Ethier & Livingston模型要稍高于实测值,而Ellsworth模型和Sharkey模型的拟合结果要稍低于实测值。初步推断,Ethier & Livingston模型和Ellsworth模型对A-Ci曲线的拟合结果要优于Sharkey模型。再根据拟合决定系数(2)可知,Ethier & Livingston模型的拟合结果精度最高,2为0.98~0.99,RMSE为0.71~ 1.47,所有植物的拟合结果均为极显著相关(< 0.01)。Ellsworth模型与Sharkey模型的拟合结果次之。因此,Ethier & Livingston模型是本研究8种植物A-Ci曲线拟合的最优生化模型。
2.2 岩溶区与非岩溶区植物的Vcmax,25和Jmax,25比较
为避免旱季水分亏缺的影响,本研究选取水分充足的主要生长季(5-7月)数据进行岩溶区和非岩溶区植物光合特征参数的比较。从图2可见,尾巨桉作为我国南方引进的重要速生造林树种,其Vcmax,25和Jmax,25明显高于其他植物。荷木的光合能力最弱, Vcmax,25和Jmax,25分别为41.43mol CO2/(m2·s)和80.84mol e–/(m2·s)。岩溶区光合能力最强的植物为雀梅藤, Vcmax,25和Jmax,25分别为63.95mol CO2/(m2·s)和129.94mol e–/(m2·s)。九龙藤的Vcmax,25和Jmax,25分别为46.21mol CO2/(m2·s)和101.63mol e–/(m2·s),为岩溶区光合能力较弱的植物。马甲子是岩溶区落叶乔木,Jmax,25达到116.04mol e–/(m2·s),与非岩溶区植物黧蒴锥的光合能力相当。而柞木和荷木虽然生境不同,但Vcmax,25相近。整体而言,岩溶区植物的光合能力与非岩溶乡土植物的差异不显著(> 0.05),但都显著低于尾巨桉(<0.05)。
2.3 植物Vcmax,25、Jmax,25与环境因子的关系
对岩溶与非岩溶区典型植物的Vcmax,25和Jmax,25与环境因子进行相关分析,结果表明(表1),尾巨桉的Vcmax,25和Jmax,25与环境温度呈极显著相关(< 0.01),决定系数(2)分别为0.99和0.97,但与土壤含水量的相关性不显著(>0.05)。非岩溶乡土植物的Vcmax,25和Jmax,25与环境温度均呈极显著相关(< 0.01),2分别为0.67和0.56,说明环境温度对非岩溶乡土植物的光合能力有较强影响。土壤含水量与非岩溶植物的Vcmax,25和Jmax,25呈极显著相关(< 0.01),2分别为0.21和0.6,说明了土壤含水量的大小对非岩溶植物的光合能力亦有一定影响。与非岩溶植物类似,岩溶区植物Vcmax,25和Jmax,25与环境温度呈极显著相关(<0.01),2分别为0.90和0.95, 可看出环境温度和岩溶植物的关系十分密切。土壤含水量与岩溶植物的Vcmax,25和Jmax,25均呈显著相关(<0.05),2分别为0.29和0.35。
3 结论和讨论
本试验采用Ethier & Livingston、Ellsworth、Sharkey等3个光合生化模型对8种岩溶和非岩溶典型植物的A-Ci曲线进行拟合,结果表明,CO2浓度为0~400mol/mol时,Sharkey模型拟合的直线斜率高于其他模型,明显偏离实测值;CO2浓度为400~600mol/mol时,Ellsworth模型对黧蒴锥、九龙藤和雀梅藤拟合的A-Ci曲线,光合速率均比实测值低;当曲线趋于稳定时,Sharkey模型对部分植物的拟合结果比实测值低,如马甲子、雀梅藤和柞木。朱中华等[17]对干旱区葡萄(叶的光合拟合研究也有相似结果,这可能是因为模型将A-Ci曲线中Rubisico和RuBP限制阶段的转折点设置为定值,难以得出最优结果。Ethier & Living- ston模型在本研究中对植物光合特征的拟合结果最优,在我国西南典型岩溶和非岩溶区具有较强的适用性。Su等对不同生化模型的拟合结果进行了比较,也认为Ethier & Livingston模型具有较好的拟合效果[18]。基于Ethier & Livingston模型的拟合结果, 表明尾巨桉的光合能力高于其他岩溶和非岩溶乡土植物(图2)。Wullschleger[19]认为桉树具有较高的光合能力,Vcmax,25和Jmax,25分别为44mol CO2/(m2·s)和154mol e–/(m2·s), 本研究结果与其基本一致。有研究表明[20],在1 800mol/(m2·s)光合有效辐射下,荷木的Vcmax,25和Jmax,25分别为32mol CO2/(m2·s)和71mol e–/(m2·s),低于本研究结果,这可能是由于荷木在强光下光合能力将受到抑制所致[21]。通常认为岩溶植物生产潜力低于同区域的非岩溶植被[22]。但本研究中岩溶植物在生长季光合能力要高于非岩溶植物。Kirschbaum等的研究表明,岩溶植物较高的光合生产潜力与其长期在岩溶环境下的高温驯化有关[23],同时邓艳则指出岩溶区广泛存在的表层岩溶水可能是岩溶植物维持高生产潜力的水分来源[24]。因此,我们认为岩溶植物在水、热、光等环境条件合适的情况下,可以具有较高的生产潜力。

图1 不同模型拟合的8种植物A-Ci响应曲线。Pn: 净光合速率; ●: 实际值; ……: Ethier & Livingston模型; ┄: Ellsworth模型; ○: Sharkey模型。
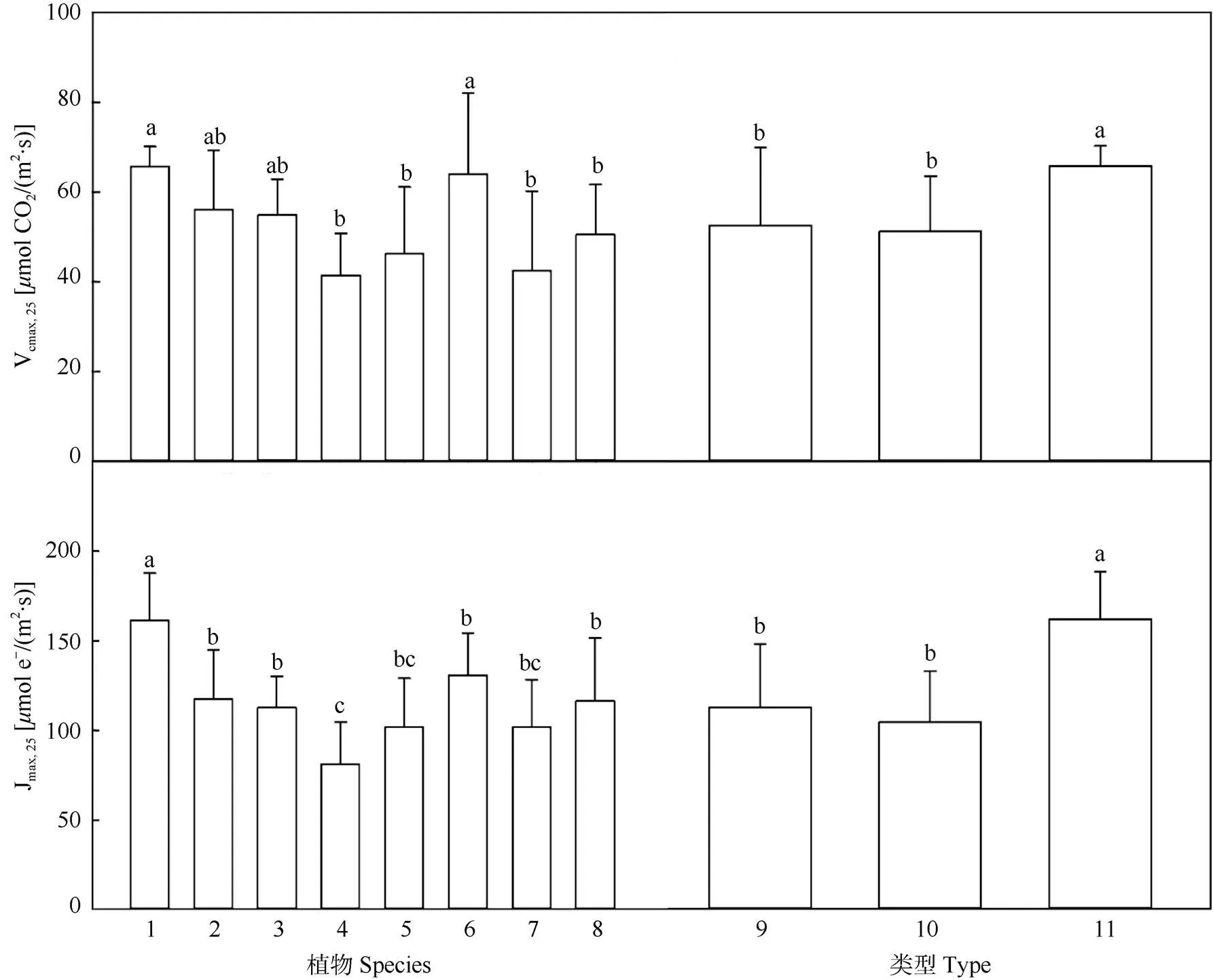
图2 生长旺季植物的Vcmax,25和Jmax,25。1: 尾巨桉; 2: 黧蒴锥; 3: 锥栗; 4: 荷木; 5: 九龙藤; 6: 雀梅藤; 7: 柞木; 8: 马甲子; 9: 岩溶植物; 10: 非岩溶乡土植物; 11: 非岩溶外来植物。柱上不同字母表示差异显著(P<0.05)。
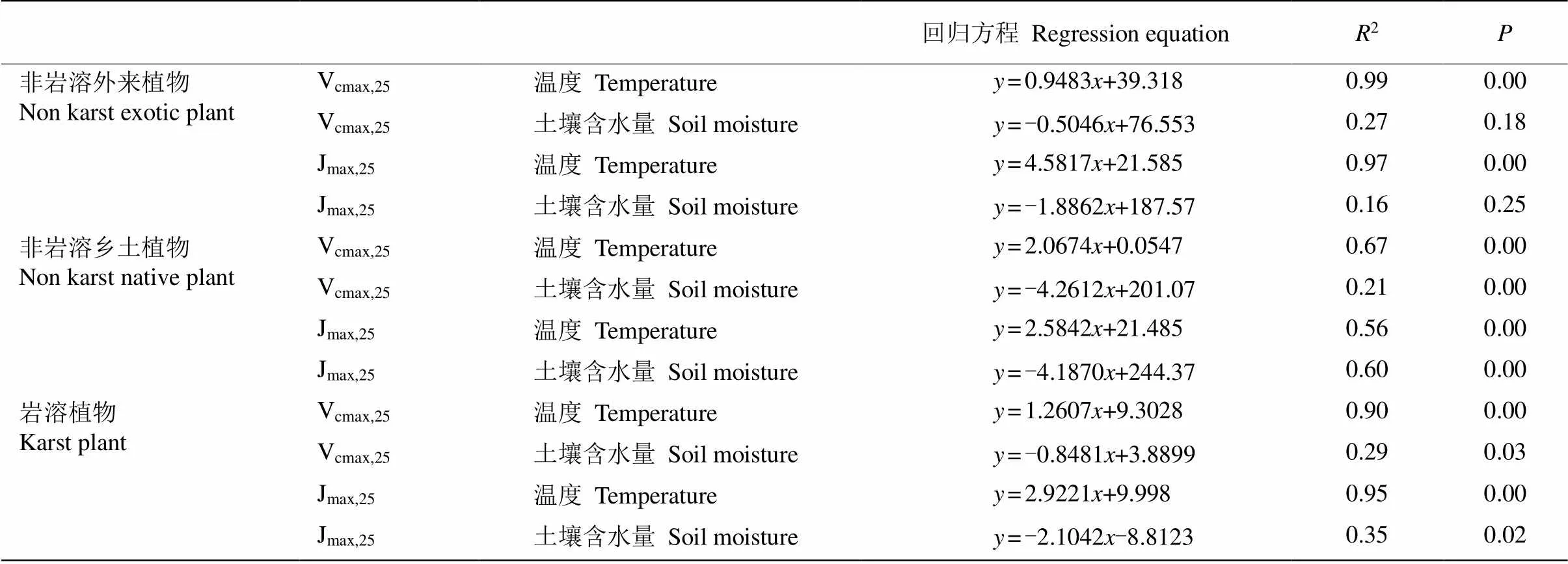
表1 不同地区植物Vcmax,25和Jmax,25与温度和土壤含水量的相关关系
植物光合过程是多个酶参与的生化反应,受环境条件尤其是温度条件的深刻影响。Vcmax是植物核酮糖1,5-二磷酸羧化酶含量和活性的最大值,受温度影响较大。Krischbaum等报道植物羧化速率随温度变化呈S型曲线[23],植物的Rubisco羧化反应随温度的升高而逐渐加速,达到一定温度时羧化反应达到最大值然后又逐渐下降,过高的温度会抑制羧化点CO2的溶解度与Rubisco的活性[25]。本研究中植物的光合能力与环境因子的相关性分析表明,无论是岩溶植物还是非岩溶植物,Vcmax,25和Jmax,25与温度均呈极显著相关(<0.01)。从表2可见,Vcmax,25与温度的拟合方程中,非岩溶乡土植物的斜率最大,说明非岩溶乡土植物的Vcmax,25对环境温度的响应最为敏感。外来种尾巨桉的Vcmax,25与温度的拟合方程斜率最小,截距最大,表明尾巨桉的光合能力受温度影响较小,在较大温度范围内保持在较高水平,显示了尾巨桉对温度的较强适应能力, 也揭示了尾巨桉在我国热带亚热带地区被广泛种植的光合生理基础。董奇妤对华南乡土树种光合能力的研究表明,尾巨桉的光能转化率处于较高水平[26],这与本研究结果一致。岩溶植物的Vcmax,25对环境温度的敏感性高于尾巨桉,但低于非岩溶乡土植物,说明岩溶植物长期生长在高温的岩溶生境中,已对干旱高温的岩溶环境产生较强的适应能力。土壤含水量与岩溶植物和非岩溶乡土植物的Vcmax,25和Jmax,25的相关性也达显著水平(< 0.05), 但整体上土壤含水量对植物Vcmax,25和Jmax,25的影响要明显小于环境温度,即环境温度是影响该地区植物光合特征的主要因素。土壤含水量与光合参数间较弱的相关性, 可能是由本研究期间土壤水分相对充足, 对植物的光合能力构成的限制较少所致。有研究指出土壤水分显著影响了植物的光合速率、气孔导度和水分利用率[27]。后续研究应加强对植物光合特征的长期监测,以获取极端干旱时期的植物光合特征响应,为极端气候变化下的植被动态研究提供依据。
本研究结果表明,Ethier & Livingston模型对我国西南典型岩溶与非岩溶区典型植物光合特征的拟合效果较好,适用于该区域植物光合特征的统一拟合估算。基于Ethier & Livingston模型的拟合结果,岩溶植物的光合能力与非岩溶乡土植物的差异不显著。非岩溶植物的Vcmax,25和Jmax,25与环境温度有显著的相关关系。相对于温度,土壤水分变化对植物Vcmax,25和Jmax,25的影响较小。
致谢 感谢中国地质调查局岩溶地质研究所会仙岩溶生态与水生态研究基地在野外试验过程中的支持。
[1] ZHANG X M, SHEN G, CHEN L, et al. Diurnal variation of photo- synthesis and response to light intensity of[J]. Guangdong Agric Sci, 2015, 42(5): 109–114. doi: 10.3969/j.issn.1004- 874X.2015.05.022.张习敏, 申刚, 陈玲, 等. 复羽叶栾树光合作用日变化及光响应特征 [J]. 广东农业科学, 2015, 42(5): 109–114. doi: 10.3969/j.issn.1004- 874X.2015.05.022.
[2] NI L K, GU D X, HE W, et al. Research advances in plant ecological adaptability in karst area [J]. Chin J Ecol, 2019, 38(7): 2210–2217. doi: 10.13292/j.1000-4890.201907.033.倪隆康, 顾大形, 何文, 等. 岩溶区植物生态适应性研究进展 [J]. 生态学杂志, 2019, 38(7): 2210–2217. doi: 10.13292/j.1000-4890.201907.033.
[3] CHEN W Y, CHEN Z Y, LUO F Y, et al. Comparison between modified exponential model and common models of light-response curve [J]. Chin J Plant Ecol, 2012, 36(12): 1277–1285. doi: 10.3724/ SP.J.1258.2012.01277.陈卫英, 陈真勇, 罗辅燕, 等. 光响应曲线的指数改进模型与常用模型比较 [J]. 植物生态学报, 2012, 36(12): 1277–1285. doi: 10.3724/ SP.J.1258.2012.01277.
[4] PENG C L, LIN Z F, SUN Z J, et al. Response of rice photosynthesis to CO2enrichment [J]. ActaPhytophysiol Sin, 1998, 24(3): 272–278.彭长连, 林植芳, 孙梓健, 等. 水稻光合作用对加富CO2的响应 [J]. 植物生理学报, 1998, 24(3): 272–278.
[5] FARQUHAR G D, VON CAEMMERER S, BERRY J A. A biochemical model of photosynthetic CO2assimilation in leaves of C3species [J]. Planta, 1980, 149(1): 78–90. doi: 10.1007/BF00386231.
[6] ETHIER G J, LIVINGSTON N J. On the need to incorporate sensi- tivity to CO2transfer conductance into the Farquhar-von Caemmerer- Berry leaf photosynthesis model [J]. Plant Cell Environ, 2004, 27(2): 137–153. doi: 10.1111/j.1365-3040.2004.01140.x.
[7] MEDLYN B E, DREYER E, ELLSWORTH D, et al. Temperature response of parameters of a biochemically based model of photo- synthesis: II. A review of experimental data [J]. Plant Cell Environ, 2002, 25(9): 1167–1179. doi: 10.1046/j.1365-3040.2002.00891.x.
[8] SHARKEY T D, BERNACCHI C J, FARQUHAR G D, et al. Fitting photosynthetic carbon dioxide response curves for C3leaves [J]. Plant Cell Environ, 2007, 30(9): 1035–1040. doi: 10.1111/j.1365-3040.2007. 01710.x.
[9] TANG X L, CAO Y H, GU L H, et al. Advances in photo-physiological responses of leaves to environmental factors based on the FvCB model [J]. Acta Ecol Sin, 2017, 37(19): 6633–6645. doi: 10.5846/stxb2016 07161450.唐星林, 曹永慧, 顾连宏, 等. 基于FvCB模型的叶片光合生理对环境因子的响应研究进展 [J]. 生态学报, 2017, 37(19): 6633–6645. doi: 10.5846/stxb201607161450.
[10] YUAN D X. Challenges and opportunities for karst research of our country under the new situation [J]. Carsol Sin, 2009, 28(4): 329–331. doi: 10.3969/j.issn.1001-4810.2009.04.001.袁道先. 新形势下我国岩溶研究面临的机遇和挑战 [J]. 中国岩溶, 2009, 28(4): 329–331. doi: 10.3969/j.issn.1001-4810.2009.04.001.
[11] KOLB T E, MATYSSEK R. Limitations and perspectives about scaling ozone impacts in trees [J]. Environ Pollut, 2001, 115(3): 373–393. doi: 10.1016/S0269-7491(01)00228-7.
[12] YE Z P, YU Q. A comparison of response curves of winter wheat photosynthesis to flag leaf intercellular and air CO2Concentrations [J]. Chin J Ecol, 2009, 28(11): 2233–2238. 叶子飘, 于强. 光合作用对胞间和大气CO2响应曲线的比较 [J]. 生态学杂志, 2009, 28(11): 2233–2238.
[13] DE PURY D G G, FARQUHAR G D. Simple scaling of photosynthesis from leaves to canopies without the errors of big-leaf models [J]. Plant Cell Environ, 1997, 20(5): 537–557. doi: 10.1111/j.1365-3040.1997.00094.x.
[14] WALKER A P, BECKERMAN A P, GU L H, et al. The relationship of leaf photosynthetic traits-cmaxandmax-to leaf nitrogen, leaf pho- sphorus, and specific leaf area: A meta-analysis and modeling study [J]. Ecol Evol, 2014, 4(16): 3218–3235. doi: 10.1002/ece3.1173.
[15] FAN Y Z, ZHONG Z M, ZHANG X Z. Determination of photo- synthetic parameterscmaxandmaxfor a C3plant (spring hulless barley) at two altitudes on the Tibetan Plateau [J]. Agric For Meteorol, 2011, 151(12): 1481–1487. doi: 10.1016/j.agrformet.2011.06.004.
[16] HIKOSAKA K, NIINEMETS Ü, ANTEN N P R. Canopy Photosyn- thesis: From Basics to Applications [M]. Dordrecht: Springer, 2016: 3–22.
[17] ZHU Z H, HAN T, LIU J Q, et al. Biochemically-based model for photosynthetic parameter estimation using Bayesian method and its application in grapes in arid region [J]. Chin J Eco-Agric, 2017, 25(6): 876–883. doi: 10.13930/j.cnki.cjea.160967.朱中华, 韩拓, 柳金权, 等. 基于贝叶斯方法的光合作用生化模型参数估计及其在干旱区葡萄上的应用 [J]. 中国生态农业学报, 2017, 25(6): 876-883. doi: 10.13930/j.cnki.cjea.160967.
[18] SU Y H, ZHU G F, MIAO Z W, et al. Estimation of parameters of a biochemically based model of photosynthesis using a genetic algorithm [J]. Plant Cell Environ, 2009, 32(12): 1710–1723. doi: 10.1111/j.1365- 3040.2009.02036.x.
[19] WULLSCHLEGER S D. Biochemical limitations to carbon assimi- lation in C3plants: A retrospective analysis of theA/Ccurves from 109 species [J]. J Exp Bot, 1993, 44(5): 907–920. doi: 10.1093/jxb/44.5.907.
[20] SUN G C, EHLERINGER J R. Gas exchange in, a sub- tropical monsoonal forest tree [J]. Photosynthetica, 1986, 20: 158–163.
[21] ZHANG J Z, LIN G Z, LIN Z F, et al. Response of photosynthesis to growth light intensity in some south subtropical woody plants [J]. J Trop Subtrop Bot, 2005, 13(5): 413–418. doi: 10.3969/j.issn.1005- 3395.2005.05.009.张进忠, 林桂珠, 林植芳, 等. 几种南亚热带木本植物光合作用对生长光强的响应 [J]. 热带亚热带植物学报, 2005, 13(5): 413–418. doi: 10.3969/j.issn.1005-3395.2005.05.009.
[22] TANG R B, FU M C, WANG L, et al. Spatial differences and it’s environmental factors of vegetation primary productivity and soil respiration in karst and non-karst areas [J]. Earth Environ, 2020, 48(3): 307–317. doi: 10.14050/j.cnki.1672-9250.2020.48.038.唐荣彬, 付梅臣, 王力, 等. 喀斯特岩溶、非岩溶区植被总初级生产力与土壤呼吸的空间差异及其环境因子分析 [J]. 地球与环境, 2020, 48(3): 307–317. doi: 10.14050/j.cnki.1672-9250.2020.48.038.
[23] KIRSCHBAUM M U F, FARQUHAR G D. Temperature dependence of whole-leaf photosynthesis inSieb.ex Spreng [J]. Aust J Plant Physiol, 1984, 11(6): 519–538. doi: 10.1071/pp 9840519.
[24] DENG Y. Coupling process between vegetation and epikarst water in karst critical zone, southwest typical peak-cluster depression area [D]. Wuhan: China University of Geosciences, 2018. 邓艳. 西南典型峰丛洼地岩溶关键带植被-表层岩溶水的耦合过程 [D]. 武汉: 中国地质大学, 2018.
[25] WALCROFT A S, WHITEHEAD D, SILVESTER W B, et al. The response of photosynthetic model parameters to temperature and nitrogen concentration inD. Don [J]. Plant Cell Environ, 1997, 20(11): 1338–1348. doi: 10.1046/j.1365-3040.1997.d01-31.x.
[26] DONG Q Y. Effects of light utilization capacity on seedlings ofMixed with native thee species [D]. Guangzhou: South China Agricultural University, 2016. 董奇妤. 尾巨桉与乡土树种苗木混交对光能利用的效应 [D]. 广州: 华南农业大学, 2016.
[27] CAI X A, SUN G C, ZHAO P, et al. The effects of soil water content on photosynthesis in leaves ofseedlings [J]. J Trop Subtrop Bot, 2004, 12(3): 207–212. doi: 10.3969/j.issn.1005-3395. 2004.03.003.蔡锡安, 孙谷畴, 赵平, 等. 土壤水分对单性木兰幼苗光合特性的影响 [J]. 热带亚热带植物学报, 2004, 12(3): 207–212. doi: 10.3969/ j.issn.1005-3395.2004.03.003.
Studies on Optimal Photosynthetic Biochemical Model and Photosynthetic Characteristics of Typical Plants in Karst and Non-karst Regions
NI Long-kang1, HE Wen1, HUANG Ke-chao1, GU Da-xing1*, HUANG Yu-qing2*
(1. Guangxi Key Laboratory of Plant Conservation and Restoration Ecology in Karst Terrain, Guangxi Institute of Botany, Guangxi Zhuang Autonomous Region and Chinese Academy of Sciences, Guilin 541006, Guangxi,China; 2.Key Laboratory of Environmental Evolution and Resource Utilization of the Beibu Gulf,Ministry of Education &Guangxi Key laboratory of Earth Surface Processes and Intelligent Simulation, Nanning Normal University,Nanning 530001, China)
The aim was to screen the optimal model in fitting photosynthetic CO2response curve (A-Ci curve) of plants in different habitats, three biochemical models, including Ethier & Livingston, Ellsworth, and Sharkey model, were used to fit the A-Ci curves of 8 typical plants in karst and non-karst regions of southwest China. Then the photosynthetic parameters and their response to main environmental factors were analyzed. The results showed that Ethier & Livingston model was better than the other two models in fitting A-Ci curve of 8 typical plants, with determination coefficient (2) from 0.98 to 0.99 (<0.01). The photosynthetic parameters (Vcmaxand Jmax) fitted by Ethier & Livingston model had no significant differences between karst and non-karst native plants (>0.05), but both of them were significantly lower than that of×. Moreover, Vcmax,25and Jmax,25of karst and non-karst native plants were significantly correlated with air temperature and soil water content (<0.05). However, the determination coefficients (2) between air temperature and Vcmax,25or Jmax,25were higher than that between soil water content and Vcmax,25/Jmax,25for all plants except of Jmax,25of non-karst native plants. Therefore, these indicated that the influence of air temperature on Vcmax,25and Jmax,25of typical plants in this area was stronger than that of soil water content.
Karst; Photosynthetic biochemical model; Photosynthetic character; Environment factor
10.11926/jtsb.4269
2020–06–22
2020–08–18
广西创新驱动发展专项(AA-17204087-9); 广西自然科学基金项目(2018GXNSFAA294023, 2019GXNSFBA245036); 国家自然科学基金项目(41830648, 31870382, 32060243, 32060369); 广西植物研究所基本业务费(18005); 广西喀斯特植物保育与恢复生态学重点实验室自主研究课题(19-A-04-03, 18-A-02-01, 19-050-6); 广西科学院基本业务费(2019YJJ1009)资助
This work was supported by the Project for Innovation Driven Development in Guangxi (Grant No. AA-17204087-9), the Natural Science Foundation of Guangxi (Grant No. 2018GXNSFAA294023, 2019GXNSFBA245036), the National Natural Science Foundation of China (Grant No. 41830648, 31870382, 32060243, 32060369), the Fundamental Research Fund of Guangxi Institute of Botany (Grant No. 18005), the Project for Independent Research of Guangxi Key Laboratory Karst Plant Conservation and Restoration Ecology (Grant No. 19-A-04-03, 18-A-02-01, 19-050-6), and the Fundamental Research Fund of Guangxi Academy of Sciences (Grant No. 2019YJJ1009).
倪隆康(1994~ ),男,硕士,研究方向为植物生态学。E-mail: 806226856@qq.com
. E-mail: gudaxing2008@163.com; hyqcoco@gxib.cn
