有毛野生甘蓝(Brassica incana)抗蚜虫特性研究
2021-04-06左香君房朋朋李加纳梅家琴
左香君 房朋朋 李加纳 钱 伟 梅家琴,*
有毛野生甘蓝()抗蚜虫特性研究
左香君1,2房朋朋1,2李加纳1,2钱 伟1,2梅家琴1,2,*
1西南大学农学与生物科技学院, 重庆 400715;2西南大学农业科学研究院, 重庆 400715
甘蓝(var.)是重要的十字花科芸薹属蔬菜, 但生产中常遭受蚜虫危害, 鉴定抗虫资源并了解其抗虫机制是抗虫育种的重要基础。课题组前期征集到1份周身覆盖浓密表皮毛的野生甘蓝(), 为调查该材料是否具有蚜虫抗性, 并明确其抗性是否与表皮毛有关, 本研究利用该有毛野生甘蓝与1份无毛芥蓝()杂交, 对双亲及其F2:3家系中有毛和无毛家系进行室内蚜虫接种发现, 蚜虫在无表皮毛甘蓝上的平均产卵数(36.7枚)和存活天数(22.2 d)均显著高于有表皮毛甘蓝(平均产卵8.1枚, 平均存活13.1 d)。为进一步明确该野生甘蓝对蚜虫是否具有化学毒杀或排拒作用, 采用有毛和无毛甘蓝叶片的浸提液对蚜虫进行胃杀、触杀和拒食性调查发现, 2种浸提液对蚜虫均无胃杀和触杀作用, 但有毛甘蓝浸提液对蚜虫存在一定的趋避作用。表明野生甘蓝对蚜虫的抗性一方面源于其表皮毛的物理阻碍, 另一方面与其体内化学物质的趋避作用有关。
甘蓝; 蚜虫; 表皮毛; 抗性鉴定; 抗虫特性
甘蓝(L., CC, 2= 18)是十字花科芸薹属植物, 具有多个栽培变种, 如结球甘蓝、花椰菜、青花菜、芥蓝、苤蓝等, 均是人们非常喜爱的蔬菜, 也是重要的经济作物, 在我国栽培面积广泛[1]。甘蓝类蔬菜经连年栽培后, 有害生物逐年增多, 病虫害逐年严重, 其中蚜虫是甘蓝等芸薹属蔬菜的主要虫害之一[2-3], 导致甘蓝产量及质量下降。此外, 甘蓝的近缘物种也受到蚜虫危害, 如受蚜虫危害并经蚜虫传播的病毒病能使甘蓝型油菜(L., AACC, 2= 38)减产10%左右, 严重时可减产20%~30%[4]。
蚜虫属同翅目(Homoptera)蚜总科(Aphidoidea), 种类繁多, 繁殖速度快, 繁殖能力强。蚜虫主要多以成虫和若虫的形态聚集在叶片和幼苗背面, 然后通过刺吸式口器刺入植株的茎、叶和嫩梢中吸食植物汁液。植株受害后, 生长率降低, 发育不良, 植株矮小; 叶片变黄、枯萎、卷曲, 还会形成褐色斑点[5-6]; 严重时会导致结球甘蓝不结球。蚜虫刺吸引起的排出体外的植株汁液, 还能够招引蚂蚁, 诱发植物感染霉菌, 煤污病。此外, 蚜虫还会携带传播病毒[7], 能够传播200多种植物病毒, 是世界传播病毒种类最多的一种昆虫。目前人们对蚜虫危害的防控主要依赖于药剂防治, 不仅引起环境和食品安全问题, 并且增加了种植成本。有学者通过调整种植密度等栽培手段提高抗虫能力进行研究, 但效果十分有限[8]。
培育抗虫品种是解决农作物蚜虫问题的治本之道, 寻找有效的抗虫资源并了解其抗虫机制是推动抗虫育种的前提和根本。目前栽培甘蓝中缺乏有效抗源, 而甘蓝的野生类型(如、、等, CC, 2= 18)长期生活在严苛的野外环境下, 可能具有抗虫资源。课题组前期征集到9类野生甘蓝, 其中周身具有浓密的表皮毛。植物表皮毛是存在于植物表皮上的一种毛状结构, 具有抗生物和非生物逆境的功能。有研究表明, 某些植物的表皮毛与其抗蚜虫能力有关[9-11]。因此, 本研究对野生甘蓝进行了抗蚜虫特性的研究, 明确了其蚜虫抗性既与表皮毛结构有关, 又与其内在物质有关, 可为甘蓝及近缘物种的抗蚜虫育种提供新的抗源。
1 材料与方法
1.1 供试材料
本研究试验材料包括有毛野生甘蓝(, 编号C01)、无毛栽培甘蓝(芥蓝, 编号C41), 以及由两者发展的F2:3家系。所有材料种植于西南大学实验基地, 采取常规田间管理。
1.2 供试虫源
从人工气候室培养的甘蓝植株上采集蚜虫, 之后转移到放有结球甘蓝(蚜虫易取食材料)的嫩叶上饲养, 饲养场所为人工气候箱, 饲养条件为: 温度24℃, 相对湿度72%, 16 h光照/8 h黑暗。选取一龄若虫作进行接种。
1.3 蚜虫接种试验
对C01、C41、21个有毛F2:3家系、14个无毛F2:3家系分别进行室内蚜虫接种试验, 具体方法如下: 在直径为35 mm的培养皿底部放1张湿润滤纸, 每个材料取1片幼嫩叶片置于滤纸之上, 用毛笔挑入一头初产若蚜, 置于人工气候培养箱中(温度24℃, 相对湿度72%, 16 h光照/8 h黑暗), 每2 d更换一次叶片。接种后每12 h观察一次, 记录蚜虫产卵数及死亡情况, 统计后及时用毛笔剔除初产若蚜, 直至成虫死亡。每个材料共计接种20头蚜虫, 每年3次重复, 整个试验重复2年。
1.4 甘蓝对蚜虫的作用方式
1.4.1 叶片浸提液制备 参考李波等[12]的方法, 分别取有表皮毛和无表皮毛的甘蓝幼嫩叶片, 洗净泥土, 滤纸吸干表面水分, 称取10 g叶片, 充分研磨后加入4 mL 95%的乙醇, 振荡, 浸泡48 h后过滤, 双蒸水定容至20 mL, 制成母液备用。对照溶液为4 mL 95%乙醇+16mL双蒸水。
1.4.2 触杀活力测定 参考巩芳娥等[13]的方法, 将20头大小一致的蚜虫接入幼嫩的结球甘蓝叶片(蚜虫易取食材料)上, 放入9cm培养皿中, 培养皿底铺上湿润的滤纸。待试虫在叶片上稳定4h后, 喷施新鲜的甘蓝提取液及对照液, 每个处理用量5 mL。将其放入人工气候箱, 处理后12、24和36h观察结果, 记录蚜虫的死亡数(死亡标准为试虫不能取食, 触之不动)。整个试验进行3次重复。
1.4.3 胃毒活力测定 参考王晶等[14]的方法, 将结球甘蓝叶片(蚜虫易取食材料)置于新鲜的提取液中浸泡1h, 取出用吸水纸吸去多余提取液, 放于培养皿中, 接入大小一致的蚜虫20头(饥饿处理4h), 每处理重复3次。将培养皿放入人工气候箱, 于接种后24h记录蚜虫的死亡数。
1.4.4 拒食活力测定 参考张建英等[15]的方法, 稍作改进。将结球甘蓝叶片(蚜虫易取食材料)打成直径为2cm的叶碟, 将叶碟放入新鲜甘蓝叶片提取液中浸湿表面, 镊子夹出后用滤纸吸干表面残余液体。将每个叶碟单独放入一个培养皿中, 接入大小一致的蚜虫20头。将培养皿放入人工气候箱, 于接种后12、24和36h统计叶碟上的活蚜数量。整个试验进行3次重复。
1.5 数据分析
采用SPSS软件对数据进行方差分析、差异显著性检验及多重比较。
2 结果与分析
经观察, 两亲本在叶和茎上存在明显差异(图1), 前期研究表明, 甘蓝表皮毛性状由1对隐性基因控制[16], 因此本试验从F2:3家系中选择了表皮毛性状无分离的株系进行蚜虫抗性鉴定。
对双亲及21个有毛和14个无毛F2:3家系进行室内蚜虫接种发现, 无毛亲本C41上的蚜虫平均产卵数为43枚, 寿命为20.5 d, 而有毛甘蓝亲本C01叶片饲养的蚜虫产卵数为1枚, 寿命9 d。对F2:3家系进行统计发现, 蚜虫在无表皮毛甘蓝上的平均产卵数和存活天数均高于有表皮毛甘蓝, 其中无毛甘蓝叶片饲养的蚜虫平均产卵36.7枚, 平均寿命为22.2 d, 而有毛甘蓝叶片饲养的蚜虫平均产卵8.1枚, 平均寿命为13.1 d (图2)。表明有毛甘蓝C01具有一定的蚜虫抗性, 这种抗性与其表皮毛有关。
**和***分别表示在0.01和0.001水平显著差异。
** and *** mean significant differences at the 0.01 and 0.001 probability levels, respectively.
为调查有毛甘蓝对蚜虫的抗性是否具有化学效应, 本研究利用叶片提取液进行了蚜虫的胃杀、触杀和拒食试验。将饥饿处理4h的蚜虫接种于分别涂抹有表皮毛甘蓝浸提液(T1)、无表皮毛甘蓝浸提液(T2)、对照提取液(CK)的叶片发现, 接种后24h的蚜虫死亡率分别为1.4、1.8和1.6头皿-1, 3种处理之间无显著差异(>0.05) (表1), 表明甘蓝浸提液对蚜虫无明显胃杀作用。此外, 对蚜虫喷洒T1、T2浸提液发现, 试验组与对照组在接种后12、24、36h的蚜虫死亡数均没有显著差异(>0.05) (表1), 表明甘蓝浸提液对蚜虫也没有触杀作用。
分别将20头蚜虫接种在浸泡有毛甘蓝浸提液T1、无毛甘蓝浸提液T2和对照液CK的饲喂叶片上发现, 接种后12 h和24h, T1处理中叶片附着的蚜虫量略低于CK和T2处理, 而接种36h后, T1处理中叶片附着的蚜虫量(平均为5.2头皿-1)则显著低于CK (平均为7.6头皿-1)和T2处理(平均为8.4头皿-1) (<0.05) (图3)。表明, 有毛甘蓝叶片的浸提液对蚜虫存在一定的趋避作用。
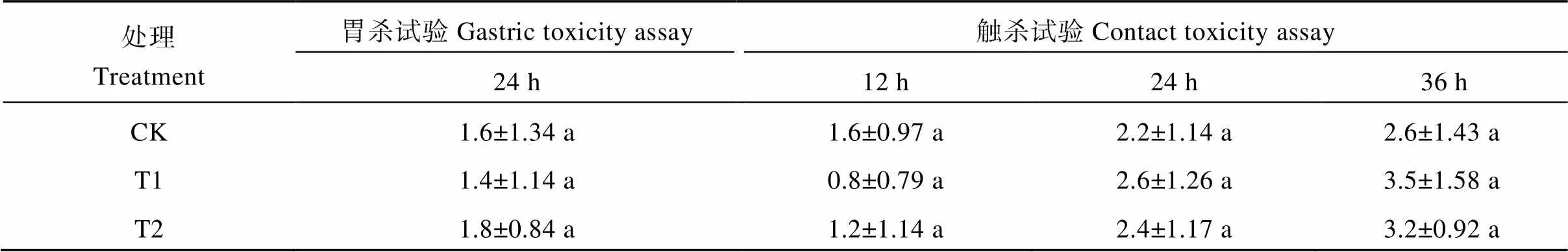
表1 不同处理中的蚜虫死亡数
CK: 对照提取液; T1: 有表皮毛甘蓝浸提液; T2: 无表皮毛甘蓝浸提液。相同小写字母表示在0.05水平无显著差异。
CK: control; T1: the extracts from hairy leaves; T2: the extracts from glabrous leaves. Values followed by the same lowercase letters mean no significant differences at the 0.05 probability level.
*表示在0.05水平显著差异。处理同表1。
* mean significant differences at the 0.05 probability level. Treatments are the same as those given in Table 1.
3 讨论
野生甘蓝分布于西西里群岛海岸, 长期生活在严苛的野外环境下, 其周身覆盖有浓密的表皮毛, 可能对其优异的生存能力有重要贡献。前人研究表明,具有菜粉蝶和甘蓝根蝇的抗性[17], 这种抗虫特性很可能与其表皮毛有关[18-20]。本研究中,及其有毛后代对蚜虫均表现出抗性, 表明的抗虫特性与其表皮毛有关。植物表皮毛一方面可能通过物理防御作用, 使蚜虫在取食、繁殖时受到阻碍, 另一方面也可能存在化学作用, 有表皮毛的植株能分泌一些化学物质, 对蚜虫具有杀灭作用或是趋向性作用。众所周知, 十字花科植物的表皮毛并不具有分泌化学物质的功能, 因此推测的表皮毛对蚜虫具有物理障碍作用。
尽管如此, 本研究还对另一种有表皮毛的野生甘蓝(地理分布与相近)进行蚜虫抗性鉴定发现,对蚜虫的抗性较弱(本文未展示), 这暗示对蚜虫的抗性很可能不止源于其表皮毛的物理障碍。本研究随后对有毛、无毛甘蓝叶片的浸提液进行研究发现, 有毛甘蓝提取液对蚜虫虽然没有灭杀作用, 但是可发挥一定的趋避作用, 这表明了虽然不产生可以毒杀蚜虫的化学物质, 但可能存在某种(些)能影响蚜虫趋向性的成分。我们猜想, 这种(些)成分虽不是由表皮毛分泌产生, 但也许与表皮毛性状存在一定相关性(比如控制两者的遗传位点存在一定程度的连锁等)。具体是何种物质对蚜虫产生趋避性及其与表皮毛的关系还有待研究。
植物抗虫机制主要分为排拒作用、抗生作用和耐害作用, 其中排拒作用指的是植物本身固有的物理形态(如颜色、形态、体表毛状物、表面蜡质、组织厚度等)以及生化特性(产生的次生代谢产物)对害虫产生不良作用, 影响的害虫取食、产卵、寄生及生活周期等的防御行为; 抗生作用是指植物中含有对害虫有毒的物质或植物中害虫所必需的营养物质含量低, 能影响害虫的生长、发育、繁殖等; 耐害作用是指植物被蚜虫取食后表现出的较强的补偿能力, 能减少害虫对植物正常生长的影响[15]。综合分析, 本研究中对蚜虫具有排拒作用, 该作用既源于其表皮毛的物理阻碍, 也与其体内物质对蚜虫的趋避作用有关。
甘蓝在自然界存在多种野生和栽培类型, 遗传基础广泛, 不同甘蓝类型间存在较大的遗传差异, 甚至存在一定生殖障碍[21-22]。尽管如此, 研究发现与栽培甘蓝的遗传关系相对于其他野生甘蓝类型而言最近[23],与各种栽培甘蓝杂交普遍能结实, 且结实率甚至为其自由授粉结实率的1.2倍[24]。结合良好的抗虫性和抗病性[25-26], 野生甘蓝对现有栽培甘蓝的改良具有较大的应用潜力。
4 结论
野生甘蓝具有一定程度的蚜虫抗性, 其抗性一方面源于其表皮毛的物理阻碍, 另一方面源于体内化学物质的趋避作用。
[1] 金珂旭, 王正银, 樊驰, 刘辉, 何德清. 不同钾肥对甘蓝产量、品质和营养元素形态的影响. 土壤学报, 2014, 51: 1369–1377. Jin K X, Wang Z Y, Fan C, Liu H, He D Q. Effects of different potassium fertilizers on the yield, quality and nutrient element forms of cabbage., 2014, 51: 1369–1377 (in Chinese with English abstract).
[2] 陆裕华. 甘蓝主要虫害的发生为害及防治. 福建农业科技, 2014, (7): 56–59. Lu Y H. Occurrence and control of main pests in cabbage., 2014, (7): 56–59 (in Chinese with English abstract).
[3] 黄毅. 甘蓝蚜的识别与防治. 农业灾害研究, 2014, 4(5): 1–3. Huang Y. Identification and control of cabbage aphids., 2014, 4(5): 1–3 (in Chinese with English abstract).
[4] 蒋卫宁, 李萌. 秋冬之际要注意防治油菜病毒病. 四川农业科技, 1999, (6): 3–5. Jiang W N, Li M. Attention should be paid to preventing rape virus disease in autumn and winter.,1999, (6): 3–5 (in Chinese).
[5] 方燕, 乔格侠, 张广学. 蚜虫寄主植物与取食部位的多样性. 动物分类学报, 2006, 31(1): 31–39. Fang Y, Qiao G X, Zhang G X. The diversity of host plants and feeding parts of aphids.,2006, 31(1): 31–39 (in Chinese).
[6] 郑竹胜, 邢鲲, 赵飞, 王建民, 赵凌瑞. 甘蓝蚜虫田间防治研究. 中国果菜, 2015, 35(4): 59–61. Zheng Z S, Xing K, Zhao F, Wang J M, Zhao L R. Study on field control of cabbage aphids., 2015, 35(4): 59–61 (in Chinese with English abstract).
[7] 高德良, 庄占兴, 庄治国. 几种新颖的杀虫剂对甘蓝蚜虫防治效果研究. 生物灾害科学, 2018, 41(4): 299–302. Gao D L, Zhuang Z X, Zhuang Z G. Study on the control effect of several novel insecticides on cabbage aphids., 2018, 41(4): 299–302 (in Chinese with English abstract).
[8] 陈红琳, 陈尚洪, 沈学善, 蒋梁材, 刘定辉. 种植密度对四川盆地丘陵区移栽油菜农艺性状和产量的影响. 中国农学通报, 2012, 28(30): 142–145. Chen H L, Chen S H, Shen X S, Jiang L C, Liu D H. Effect of planting density on agronomic characters and yield of transplanted rape in hilly area of Sichuan basin., 2012, 28(30): 142–145 (in Chinese with English abstract).
[9] 尚宏芹, 刘建萍, 戴洪义, 高昌勇, 赵丽兰, 孙显明. 辣椒体表茸毛与抗蚜虫关系的研究. 莱阳农学院学报, 2004, 21(4): 293–295. Shang H Q, Liu J P, Dai H Y, Gao C Y, Zhao L L, Sun X M. Study on the relationship between pepper body surface hair and aphid resistance., 2004, 21(4): 293–295 (in Chinese with English abstract).
[10] 何俊平, 陈发棣, 陈素梅, 房伟民. 不同菊花品种抗蚜虫性鉴定. 生态学杂志, 2010, 29: 1382–1386. He J P, Chen F L, Chen S M, Fang W M. Identification of aphid resistance of different chrysanthemum varieties., 2010, 29: 1382–1386 (in Chinese with English abstract).
[11] 郭宝德, 姜艳丽, 冀丽霞, 黄穗兰.基因导入形态抗蚜棉研究. 山西农业科学, 2013, 41: 1171–1173. Guo B D, Jiang Y L, Ji L X, Huang H L. Study ongene transfer morphological resistance cotton., 2013, 41: 1171–1173 (in Chinese with English abstract).
[12] 李波, 赵永田, 高甜甜, 金洪, 陶科, 侯太平. 5种植物乙醇提取物抑菌杀虫活性. 福建农业学报, 2019, 34: 451–457. Li B, Zhao Y T, Gao T T, Jin H, Tao K, Hou T P. Antibacterial and insecticidal activity of 5 kinds of plant ethanol extracts.,2019, 34: 451–457 (in Chinese with English abstract).
[13] 巩芳娥, 张进德, 贾星宏. 核桃外果皮提取液对蚜虫杀虫活性试验. 林业科技通讯, 2017, (2): 42–44. Gong F E, Zhang J D, Jia X H. Test on the insecticidal activity of walnut exocarp extract against aphids., 2017, (2): 42–44 (in Chinese with English abstract).
[14] 王晶, 张莹, 周春相, 王艺卉, 邓颖, 袁海滨. 野艾蒿精油化学分离各组分对小菜蛾的杀虫活性. 吉林农业大学学报, 2018, 40: 675–680. Wang J, Zhang Y, Zhou C X, Wang Y H, Deng Y, Yuan H B. Insecticidal activity of the chemically separated components of the essential oil ofagainst., 2018, 40: 675–680 (in Chinese with English abstract).
[15] 张建英, 杨贵军, 于有志, 杨乐. 11种植物提取物对枸杞蚜虫的拒食和毒杀活性测定. 农业科学研究, 2007, 28(1): 21–23. Zhang J Y, Yang G J, Yu Y Z, Yang L. Determination of antifeedant and toxic activity of 11 plant extracts against wolfberry aphids., 2007, 28(1): 21–23 (in Chinese).
[16] Mei J, Wang J, Li Y, Tian S, Wei D, Shao C, Si J, Xiong Q, Li J, Qian W. Mapping of genetic locus for leaf trichome in., 2017, 130: 1953–1959.
[17] Ellis P R, Pink D A C, Barber N E, Mead A. Identification of high levels of resistance to cabbage root fly,, in wildspecies., 1999, 110: 207–214.
[18] Egea-Gilabert C, Bilotti G, Requena M E, Ezziyyani M, Vivo-Molina J M, Candela M E. Pepper morphological traits related with resistance to.,2008, 52: 105–109.
[19] Elle E, Dam N M, Hare J D. Cost of glandular trichomes, a “resistance” character inRegel (Solanaceae)., 1999, 53: 22–35.
[20] Kim J K, Han J K, Kim S, Lee H R, Shin J S, Kim J H, Cho J, Kim Y H, Lee H J, Kim B D, Choi D. Trichome density of main stem is tightly linked toresistance in chili pepper (L.)., 2011, 122: 1051–1058.
[21] Bothmer R V, Gustafsson M, Snogerup S.sect.(Brassicaceae): II. Inter- and intraspecific crosses with cultivars of.,1995, 42: 165–178.
[22] Kianian S F, Quiros C F. Trait inheritance, fertility, and genomic relationships of some= 9species.,1992, 39: 165–175.
[23] Mei J, Li Q, Yang X, Qian L, Liu L, Yin J, Frauen M, Li J, Qian W. Genomic relationships between wild and cultivatedLwith emphasis on the origination of cultivated crops.,2010, 57: 687–692.
[24] 丁一娟, 魏大勇, 刘瑶, 梅家琴, 钱伟. 不同类型甘蓝的可交配性研究. 植物遗传资源学报, 2013, 14: 342–346. Ding Y J, Wei D Y, Liu Y, Mei J Q, Qian W. Study on the mating ability of different types of cabbage., 2013, 14: 342–346 (in Chinese with English abstract).
[25] Mithen R F, Magrath R. Glucosinolates and resistance toin wild and cultivatedspecies., 1992, 108: 60–68.
[26] Mei J, Qian L, Disi J O, Yang X, Li Q, Li J, Frauen M, Cai D, Qian W. Identification of resistant sources againstinspecies with emphasis on.,2011, 177: 393–399.
Characterization of aphid-resistance of a hairy wildtaxa,
ZUO Xiang-Jun1,2, FANG Peng-Peng1,2, LI Jia-Na1,2, QIAN Wei1,2, and MEI Jia-Qin1,2,*
1College of Agronomy and Biotechnology, Southwest University, Chongqing 400715, China;2Academy of Agricultural Sciences, Southwest University, Chongqing 400715, China
contains important vegetables varieties in our daily life, however, the production ofvegetables always suffers from aphids. Identifying resistant resource and understanding corresponding resistance mechanism are of great importance for resistance breeding of crops. In our previous study, a wild relative ofwith dense trichomes on leaves and stem was obtained. In the present study, to investigate the resistance ofto aphids and to understand the role of trichomes in resistance, the two parental accessions and their F2:3lines were inoculated with aphids in lab condition. The results showed that the egg production and lifespan of aphids on hairless leaves (36.7 eggs and 22.2 days) were significantly higher than corresponding values on hairy leaves (8.1 eggs and 13.1 days). To investigate whether a chemical role contributed to the resistance, the contact toxicity, gastric toxicity and anti-feeding ability of the leaf extracts were checked between hairy and glabrous F2:3lines. None of the extract was found to be with significant contact toxicity and gastric toxicity to aphids, whereas the number of aphids on leaves treated with extract from hairywas significantly lower than that treated by extract of glabrous leaves. This study revealed that the resistance ofto aphids might attribute to both the physical inhibition from trichomes and the avoidance role of its chemical compounds.
; aphid; trichome; resistance identification; resistance characteristics
10.3724/SP.J.1006.2021.04147
本研究由国家重点研发计划项目(2018YFD0100500)资助。
This study was supported by the National Key Research and Development Program of China (2018YFD0100500).
梅家琴, E-mail: jiaqinmay@163.com
E-mail: fanta152521@163.com
2020-07-05;
2020-12-01;
2021-01-10.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20210108.1357.004.html
