水稻巨大胚突变体N2-52垩白形成机制研究
2021-03-29葛鑫源刘世家李欣燕海刚王永祥傅玉双单壮壮张文伟王益华江玲
葛鑫源,刘世家,李欣,燕海刚,王永祥,傅玉双,单壮壮,张文伟,王益华,江玲
(南京农业大学农业农村部长江中下游粳稻生物学与遗传育种重点实验室/长江流域杂交水稻协同创新中心/江苏省现代作物生产中心,江苏 南京 210095)
垩白是由于稻米胚乳中淀粉颗粒排列疏松、颗粒间存在间隙,引起光的折射而产生的,根据垩白在籽粒中的部位,将其划分为腹白、心白和背白[1]。描述垩白的综合指标有2个,即垩白率和垩白度,垩白严重的水稻在加工过程中易断,整精米率降低,蒸煮后米粒断裂,影响食味品质。减少垩白率和降低垩白度是稻米品质改良的主攻目标,深入研究垩白的形成及产生垩白的分子机制是育种上克服这一不利性状的前提。垩白是一个复杂的数量性状,受环境和基因型的共同作用。已报道与水稻垩白形成有关的QTL有 82个,分布于除第4染色体外的11条染色体上[1],其中第8染色体27个,第9染色体18个(数据来源网站 http://www.gramene.org/archive/qtl)。Wx位点是影响垩白形成的主效QTL之一[2]。此外,已知多个与淀粉合成相关的基因影响垩白的形成,包括OsPPDKB[1]、SSⅢa[3]、PHO1[3]、BT1[3]和OsPKpα1[4],这些基因的突变均可导致淀粉颗粒的形态异常和垩白的产生。但是,关于垩白形成与淀粉合成之间的具体联系,目前仍不明确。
巨大胚基因(giant embryo,GE)编码1个细胞色素P450超家族的CYP78A亚家族成员[5]。GE突变会导致水稻籽粒的胚增大、胚乳缩小,并影响胚发育、茎顶端分生组织的维持和籽粒产量。GE还在短链脂肪酸代谢中发挥作用。虽然GE在胚和胚的小叶上皮中表达[6],但GE突变体胚乳出现了垩白表型[5],原因尚待阐明。
本研究室之前筛选到的1个水稻巨大胚突变体N2-52,具有明显的垩白表型。本研究分析N2-52籽粒理化性质、淀粉颗粒和造粉体形态,比较不同灌浆时期、籽粒不同部位的淀粉合成酶积累量,测定蔗糖合酶活性;通过分析N2-52胚乳淀粉合成途径的变化,探讨GE基因影响垩白形成的分子机制。
1 材料与方法
1.1 材料和农艺性状观察
在粳稻品种‘宁粳2号’经甲磺酸乙酯(ethyl methane sulfonate,EMS)化学诱变获得的M2代群体中,筛选出稳定遗传的巨大胚突变体N2-52。将N2-52与籼稻品种‘N22’杂交获得F1种子,并衍生得到F2突变基因定位群体。2018年正季将N2-52及其野生型种植在南京农业大学牌楼实验基地,分别随机选取 20株成熟植株,考察株高、穗长、结实率等农艺性状。
1.2 成熟种子横断面的扫描电镜观察及半薄切片样品制备和观察
在日立 S-3000N 型扫描电子显微镜(加速电压5 kV)下观察N2-52及其野生型成熟种子的横断面。
在水稻开花后 9 d,选取96个籽粒的胚乳,在紧邻胚下的胚乳部位,用单面刀片将胚乳切成厚度为 1~2 mm 的薄片,在4 ℃条件下于戊二醛固定液中过夜;在-20 ℃冰箱中脱水、渗透,在室温下用100% LR White树酯进行包埋,封口膜封好,60 ℃烘箱放置48 h聚合。修样后用切片机切片,样品厚度1 μm,用捞样器将切片捞至干净的载玻片上,60 ℃烤片至水分蒸发干净;在样品上适量滴加 1% I2-KI 溶液着色,显微镜下观察淀粉颗粒并拍摄照片。
1.3 脂肪、蛋白、总淀粉、直链淀粉含量的测定
将N2-52及其野生型种子去壳后碾磨成糙米粉,过150 μm 筛,烘干至恒重。称取3.000 g 糙米粉,重复3次,采用 FOSS 公司的全自动脂肪测定仪测定脂肪含量,并根据糙米重,换算成每粒糙米的脂肪含量。
使用FOSS公司KJELTEC2300型全自动凯氏定氮仪测定总氮含量,换算成蛋白质含量(转换系数为5.95),之后换算成每粒糙米的蛋白含量。
采用Megazyme总淀粉含量测定试剂盒(爱尔兰Megazyme国际有限公司),利用酶解法,并根据说明书中的方法(a)进行测定,再换算成每粒糙米的淀粉含量。
直链淀粉含量的测定按照《水稻、玉米、谷子籽粒直链淀粉国家标准:GB 7648—1987》 的方法。
1.4 米粉的尿素膨胀体积测定
配制0~9 mol·L-1尿素溶液,并用2 mol·L-1醋酸调节pH值至6.0。称取20 mg精米粉,加入1.5 mL EP管中,分别向每个EP管加入不同浓度的尿素溶液1 mL,混合均匀后在25 ℃、200 r·min-1振荡24 h;室温下8 000g离心20 min,静置1 h。3次重复。淀粉颗粒在尿素中的溶解度用膨胀体积来衡量,即膨胀体积=总体积-上清液的体积。
1.5 米粉的黏度分析
称取N2-52及其野生型精米米粉样品(过150 μm筛)各3.000 g至铝杯中,加入25 mL ddH2O(样品、铝杯预先烘干至恒重)。重复3次。采用瑞典波通公司(Perten)的快速黏度分析仪(RVA)测定米粉的黏度特性,用Excel 2019绘图分析。
1.6 基因的图位克隆
通过杂交获得N2-52(♀)/‘N22’(♂)的F2种子,去壳后挑选与突变体表型一致的籽粒提取DNA,使用本实验室已有的分子标记,用44个极端个体进行连锁分析,确定连锁区间后增加标记并从F2种子中进一步挑选极端个体750个,开发标记进行精细定位(表1)。标记的开发基于Graneme网站和Primer Premier 5.0软件。
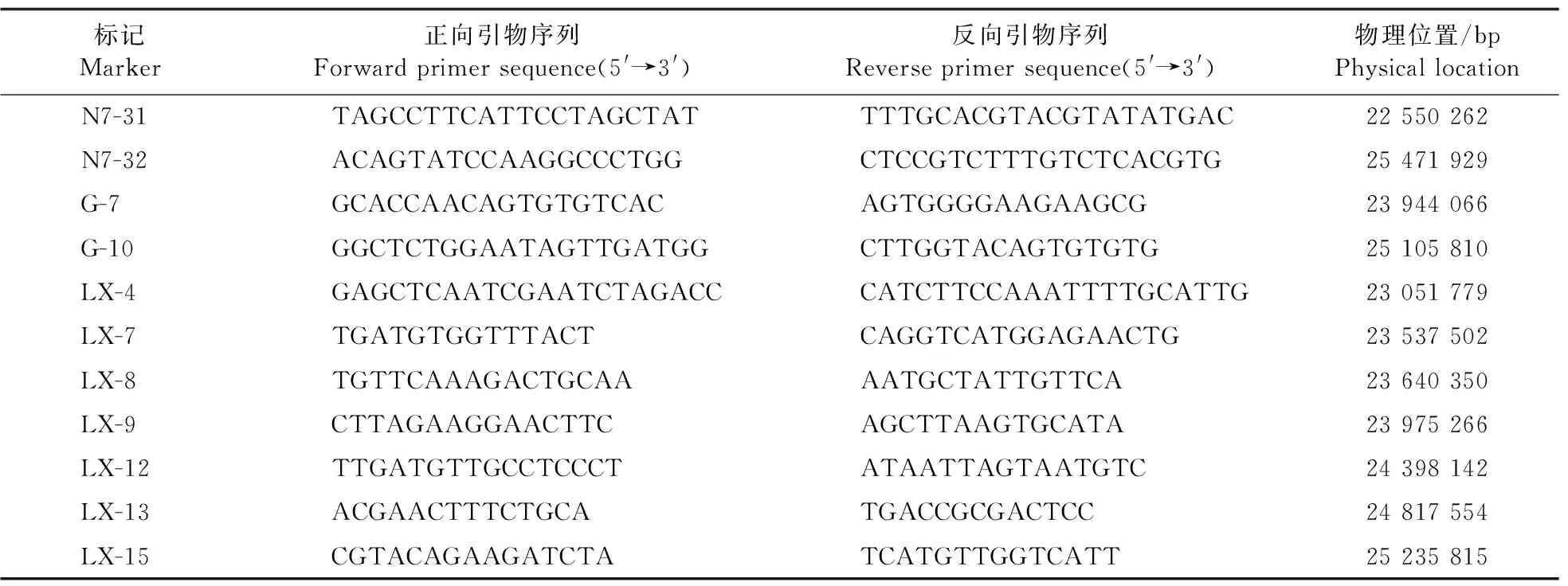
表1 N2-52突变基因精细定位所用的分子标记Table 1 Markers for fine mapping of the mutant locus of N2-52
1.7 蛋白免疫印迹分析
分别提取开花后9 d和成熟时的种子、N2-52垩白部位及其野生型对应部位的总蛋白进行SDS-PAGE。使用ExpressPlus预制胶(4%~20%)进行电泳,电泳完成后使用eBlot L1蛋白转膜仪将蛋白转印到PVDF膜上(Minipore 0.45 μm),使用封闭液(5%脱脂奶粉,PBST溶解)封闭1 h。转入含有一抗(1∶10 000)的新封闭液中孵育1 h。PBST漂洗PVDF膜3次,每次10 min。再将膜转入含有二抗(1∶5 000)的封闭液中孵育 1 h,重复漂洗。使用ECL发光液检测。所用抗体为ADP葡萄糖焦磷酸化酶小亚基Ⅱb(ADP-glucose pyrophosphorylase small subunit Ⅱb,AGPS Ⅱb)、颗粒结合淀粉合酶Ⅰ(granule bound starch synthaseⅠ,GBSSⅠ)、淀粉分支酶Ⅰ(branching enzymeⅠ,BEⅠ)、淀粉分支酶Ⅱb(branching enzymeⅡb,BEⅡb)、淀粉合酶Ⅱa(starch synthaseⅡa,SSⅡa),由北京华大蛋白质研发中心有限公司制备,内参抗体为Anti-Actin。
1.8 蔗糖合酶活性测定
蔗糖合酶活性测定参照韩小花[7]的方法。分别取开花后9 d的种子5粒,剥壳后研磨,将粉末置于2.0 mL的EP管中,按0.1 g·mL-1的比例加入提取液,4 ℃、13 000 r·min-1离心20 min,取上清液再离心 10 min,取上清液用于酶活性测定,重复3次。蔗糖合酶活性测定步骤:在10 mL洗净烘干的玻璃管中加入50 mmol·L-1蔗糖50 μL、50 mmol·L-1MgCl250 μL、Hepes(N-2-hydroxyethylpiperazine-N-ethane-sulphonicacid)缓冲液500 μL、酶提取液50 μL,振荡摇匀后30 ℃预热10 min,加入50 mmol·L-1尿苷二磷酸葡萄糖(uridine diphosphate glucose,UDPG)50 μL、50 mmol·L-1蔗糖溶液50 μL,30 ℃水浴10 min。向各反应管中加入2 mmol·L-1NaOH 100 mL,沸水浴10 min 终止反应。待冷却至室温后,加入3.5 mL 30% HCl和1 mL 0.1%间苯二酚,80 ℃水浴10 min,冷却后使用分光光度计在480 nm处测定酶活性。100 mL酶提取液配方:50 mmol·L-1Hepes、2 mmol·L-1MgCl2、50 mmol·L-1β-巯基乙醇、12.5%甘油。
2 结果与分析
2.1 水稻巨大胚突变体N2-52表型分析
相较于野生型,突变体成熟种子具有巨大胚,同时,胚下方和籽粒腹部下方产生了垩白(图1-A)。N2-52成熟种子的粒长和粒厚与野生型种子相比无明显差异,粒宽则显著高于野生型,垩白粒率极显著高于野生型。N2-52植株的每穗粒数和结实率极显著低于野生型,千粒重显著低于野生型,株高显著高于野生型(图1-B),但穗长无明显差异(表2)。

图1 N2-52及其野生型(WT)的成熟籽粒(A)和植株表型(B)Fig.1 Phenotype of N2-52 and the wild-type(WT)mature seeds(A)and plants(B)

表2 N2-52及其WT的主要农艺性状Table 2 Main agronomic traits of N2-52 and the WT
2.2 N2-52垩白部位的扫描电镜观察
对N2-52垩白部位及其野生型成熟种子的相同位置(图中红色方框位置)横断面进行扫描电镜观察,发现N2-52籽粒的横断面淀粉颗粒排列疏松,呈不规则的球体或棒状(图2-A、B、C),而野生型籽粒淀粉颗粒排列紧密,呈规则多面体(图2-D、E、F)。

图2 N2-52及其WT成熟种子横断面扫描电镜观察Fig.2 Scanning electron microscopy observation to the cross sections of N2-52 and the WT grains A、D. 种子横断面;B、C、E、F. N2-52的垩白部位和野生型相应区域(A和D红色矩形区域)。A,D. The cross sections of grains;B,C,E,F. The chalky part of N2-52 and the counterpart of the wild-type,corresponding to red rectangle in A and D separately.
2.3 半薄切片观察
取开花后9 d种子制作半薄切片,发现N2-52胚乳中淀粉颗粒排列疏松,小淀粉颗粒明显增多,正常发育的造粉体减少,胞内局部出现空腔(图3-A红色箭头所示);野生型胚乳中的淀粉颗粒排列紧密,造粉体发育正常(图3-B)。推测N2-52胚乳中空腔是由淀粉颗粒填充不足造成的。

图3 花后9 d N2-52(A)及其WT(B)胚乳半薄切片观察Fig.3 Semi-thin sections observation to N2-52(A)and the WT(B)endosperm at 9 days after flowering箭头示空腔位置。Arrows indicate empty position.
2.4 N2-52籽粒的脂肪、总蛋白含量和淀粉成分分析
N2-52成熟籽粒的蛋白质含量比野生型高12.5%,脂肪含量降低42.4%。每粒野生型种子含淀粉 15.1 mg,而每粒N2-52种子仅有12.2 mg,但是它的直链淀粉含量比野生型高27%(表3),推测N2-52的淀粉合成受阻。
表3 N2-52及其WT种子的主要化学成分Table 3 Major chemical components of N2-52 and the WT seeds
2.5 米粉的尿素膨胀实验
在低浓度尿素溶液(0~3 mol·L-1)中,N2-52米粉膨胀体积略高于野生型;在尿素浓度为4~6 mol·L-1时,N2-52膨胀体积显著高于野生型,而在高浓度尿素溶液中(8~9 mol·L-1)野生型米粉膨胀体积极显著高于N2-52(图4)。说明N2-52基因突变改变了淀粉颗粒的结构,从而改变淀粉在尿素中的溶解度。

图4 尿素浓度对N2-52及其WT米粉膨胀体积的影响Fig.4 Effects of the urea concentration on swelling volume of starch powered from grains of N2-52 and the WT
2.6 快速黏度分析仪(RVA)分析淀粉的黏度特性
相较于野生型,N2-52淀粉的黏度曲线比较平缓,总体上低于野生型(图5)。在黏度曲线的特征值方面,N2-52淀粉峰值黏度、崩解值、热浆黏度和冷胶黏度均显著高于野生型,而消减值则显著低于野生型(表4),说明N2-52基因突变改变了突变体淀粉的黏度特性。

图5 N2-52及其WT淀粉的黏度特性分析Fig.5 Viscosity profile analysis of N2-52 and the WT starch

表4 N2-52及其WT淀粉的黏度特性Table 4 The pasting properties of N2-52 and the WT starch
2.7 N2-52突变基因的精细定位
在N2-52(♀)/‘N22’(♂)的F2群体中,一共鉴定到与突变体相似表型性状的种子1 127粒,而正常的种子有3 621粒,经卡方分析χ2=2.057,符合3∶1的理论分离比例。利用图位克隆的方法,挑选出44个与突变体相似表型性状的F2个体进行定位,将突变基因连锁于第7染色体长臂N7-31与N7-32标记之间(图6-A)。进一步利用750个极端个体将基因定位于标记LX-12与LX-13之间,该区间物理距离为42 kb(图6-B)。根据水稻数据库(http://rice.plantbiology.msu.edu/)数据,该区间共有5个开放阅读框(open reading frame,ORF,图6-C),测序发现只有ORF 3即Os07g0603700在其编码区第2个外显子的第1 529个碱基由C突变为A(图6-D),导致翻译的提前终止。通过序列(https://blast.ncbi.nlm.nih.gov/Blast)比对,确定突变基因Os07g0603700为之前报道的水稻巨大胚基因GE[5-6,8]。

图6 N2-52突变基因的图位克隆Fig.6 Map-based cloning of the mutated gene from N2-52 A. N2-52突变基因与标记N7-31和N7-32连锁;B. 利用750个极端个体将N2-52基因定位于42 kb区间内;C. N2-52的基因预测(红色箭头为突变基因);D. Os07g0603700的基因结构,N2-52中发生T→A的单碱基突变。A. The N2-52 gene was linked with the markers N7-31 and N7-32;B. Using 750 individuals,the N2-52 gene was located in the 42 kb interval;C. The prediction of N2-52 candidate genes(the red arrow indicates the mutant gene);D. The gene structure of Os07g0603700,a single base mutation of T to A in N2-52.
2.8 N2-52籽粒淀粉合成酶的蛋白免疫印迹分析和酶活性测定
对N2-52开花后9 d种子以及成熟种子(整粒和垩白部位)淀粉合成酶的蛋白量进行免疫印迹分析。N2-52成熟籽粒中GBSSⅠ和BEⅠ蛋白量上升,而BEⅡb、SSⅡa蛋白量下降;其垩白部位的BEⅠ、BEⅡb、SSⅡa下降而GBSSⅠ上升,且没有检测到AGPSⅡb。在N2-52开花后9 d种子中,BEⅠ、BEⅡb和SSⅡa蛋白量均下降,而GBSSⅠ上升(图7-A)。对开花后9 d的N2-52和野生型种子的蔗糖合酶活性进行测定,发现N2-52的蔗糖合酶活性显著上升(图7-B)。

图7 N2-52及其WT淀粉合成酶的免疫印迹分析(A)和酶活性测定(B)Fig.7 Western blot analysis(A)and relative activities(B)of starch synthetases of N2-52 and the WT
3 讨论
胚乳是水稻籽粒的营养贮藏部位,其品质也是影响水稻经济的主要因素。胚乳垩白影响水稻籽粒的外观以及整精米率,是水稻品质育种最主要的改良目标之一[9],其遗传特性复杂,垩白形成的原因和调控机制还不明确[1]。
本文通过对巨大胚突变体N2-52的研究,图位克隆了GE基因。N2-52中GE基因的单碱基突变,导致巨大胚和垩白的表型。在之前的研究中,发现GE基因编码CYP78A家族的细胞色素P450,抑制水稻盾片细胞生长,维持茎顶端分生组织的生理状态[8];而对于胚中表达的GE与胚乳中的淀粉合成途径之间的联系尚未见报道。对野生型和N2-52籽粒的理化性质分析发现,突变体的淀粉颗粒结构发生改变;扫描电镜和半薄切片观察发现,N2-52垩白部位的淀粉颗粒排列松散、分布稀疏。在N2-52成熟籽粒的垩白部位,多个淀粉合成酶的表达发生变化,包括AGPS Ⅱb、BEⅠ、BEⅡb、SSⅡa、GBSSⅠ。水稻胚乳的发育成熟是一个连续的过程[10],由多种酶共同调控完成[11-14]。AGPS Ⅱb作为AGPase的小亚基,负责胚乳发育中后期淀粉的合成;GBSSⅠ负责催化直链淀粉的长葡聚链的延伸[15];BEⅠ催化产生DP≥16的长链支链淀粉;BEⅡb负责产生支链淀粉的同时降解具有分支的水溶性多聚糖,BEⅡb的基因突变会产生淀粉颗粒变小的表型;SSⅡa主要负责合成水稻中长链支链淀粉[11]。因此,这些淀粉合成酶在籽粒灌浆中对淀粉的合成和积累起不可或缺的作用,推测这些酶的表达变化,使N2-52垩白部位的淀粉合成途径受阻、淀粉颗粒结构改变从而产生垩白。N2-52成熟籽粒的直链淀粉含量相对于野生型显著增加,与GBSSⅠ表达上调的结果相一致。
CYP78A家族的细胞色素P450主要表达于胚和胚的小叶上皮中[16],而GE功能的丧失可以改变盾片的细胞壁结构[8]。由于盾片为胚的发育提供营养,盾片结构的改变使更多的营养物质进入胚中,产生巨大胚的表型[8]。在本研究中,N2-52成熟籽粒的垩白位于胚下方和腹部下方,胚下方的垩白可能由于胚的异常发育,导致进入灌浆籽粒的碳源被过度消耗,使胚乳淀粉的合成受阻而产生。腹部下方的垩白除淀粉合成受阻外还可能由于其距胚较远[10],碳源运输不足,导致淀粉填充不足进而产生。蔗糖作为水稻籽粒淀粉合成的碳源物质,直接影响库源器官之间的装卸[17]。突变体中蔗糖合酶活性的上升,反映了淀粉合成的一种补偿性上升。综上所述,GE基因突变可能造成胚乳淀粉合成受阻,从而改变淀粉颗粒的结构,产生了垩白。
Nagasawa等[5]的研究未从淀粉合成的角度来解析GE突变体垩白形成的原因。本研究对N2-52的淀粉合成途径进行了初步分析,发现GE突变导致淀粉合成酶的表达变化,但其如何影响淀粉合成,还有待进一步研究。N2-52的发现为进一步探究垩白形成的分子机制奠定了基础,对理解水稻淀粉合成和改良水稻品种具有一定意义。
