三角梅自然越冬过程的生理生化响应
2021-03-26马晓华胡青荻张旭乐刘洪见钱仁卷
马晓华,胡青荻,郑 坚,张旭乐,刘洪见,钱仁卷
(浙江省亚热带作物研究所,浙江温州325005)
0 引言
三角梅(Bougainvillea speetabilis)为紫茉莉科(Nyctaginaceae)叶子花属(Bougainvillea),又名三角花等,原产巴西,大多数为热带、亚热带木质藤本花卉[1]。三角梅的种类繁多,且花色艳丽、花期长,因此深受广大消费者的喜爱,在园艺园林中应用十分广泛[1-2]。目前,三角梅已成为园林造景中用途最广、形式最多样的美化素材之一。但三角梅性喜温暖潮湿的环境,怕冷不耐寒,对温度的要求极高。大多数品种对4℃以下的低温较为敏感,还有部分品种仅在国内南方部分地区露地栽培,其他地区都需温室培养或室内栽培,这导致三角梅的推广与栽培受到极大限制。研究低温胁迫对三角梅生长发育的影响显得尤为重要,有助于探索三角梅在低温条件下的表型、生理生化及分子的响应及低温适应能力,对三角梅的耐寒品种选育和推广应用具有重要意义。
三角梅不但具有很高的观赏价值,还具有重要的药用价值及抗病毒、抗氧化等效果[3]。大量研究表明,三角梅的叶和根部组织含有抗病毒蛋白[4-5],可抑制多种植物病毒的活性[6]。Bates[7]和Narayanan[8]等也曾从三角梅植株中提取出松醇,其具有与胰岛素类似的功能,对糖尿病有较好的疗效。近年来,关于三角梅的研究开始慢慢集中在温度方面,马若晨等[9]对4个三角梅品种的可溶性蛋白质含量进行研究,发现随着低温胁迫的加深,4个三角梅品种叶片的可溶性蛋白质含量均出现先减小后增加的趋势。邢海盈等[10]研究表明,在越冬期间,随着温度先下降后上升,3个三角梅品种叶片中ABA的含量呈先上升后下降的趋势。
目前三角梅的相关研究逐年增多,但多数集中在水肥管理、品种调查等方面[11-12],关于低温胁迫方面的系统研究仍然较少。且三角梅种类多样,不同的三角梅品种间差异十分显著,探索不同三角梅品种的越冬能力及其在低温条件下的生理生化反应是选育耐寒种质的基础。本试验选用7个2年生的三角梅品种,主要研究其在温州地区越冬过程中的生理生化变化,探索三角梅在越冬过程中的生理调节响应,以期为三角梅耐寒品种的选择及其推广提供理论基础。
1 材料与方法
1.1 试验时间、地点
试验于2019年10月—2020年2月在浙江温州浙江省亚热带作物研究所花卉资源圃(28°00′N,120°64′E)内进行,室内试验在浙江省亚热带作物研究所生化实验室进行。试验过程中正常进行水肥管理,且试验期间温州地区气候变化与往年差异不显著,无极端气候事件出现,表现为正常冬季越冬期间的气温变化。
1.2 试验材料
供试材料为引进自厦门千日红园艺公司2年生生长健壮的三角梅,分别为‘巴西紫’、‘花叶大红’、‘绿樱’、‘云南紫’、‘珍珠白’、‘柠檬黄’和‘中国红’7个品种,苗高(55±0.5)cm,地径(1.65±0.2)cm。
1.3 试验设计
2019年9月,选择长势良好、健康无病虫害的2年生正常生长的三角梅盆栽,放置在温州景山省亚作所资源圃内,土地平整,自然光线无遮挡。每个品种3个重复,每个重复10盆,共210盆。每个品种摆放在一起,采用3×10的排列方式,共7个品种,7个品种摆放位置十分接近,光线方位一致。研究7个品种三角梅在自然降温过程中(2019年10月—2020年2月)的脯氨酸、可溶性蛋白、丙二醛(MDA)、过氧化氢含量及超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)等的变化。期间进行正常的水肥管理,每隔20~25天采一次样,每次采样时间为6:30—7:30,所有品种均选取从顶端开始第3~4叶片作为测试材料。越冬期间的温度变化采用气象传感器(托普云农TPWST-IIIB-3,中国)获得,变化如图1所示。

图1 三角梅越冬期间温度随日期的变化
1.4 试验方法
采用紫外吸收法测定过氧化物酶(CAT)活性[13],愈创木酚法测定过氧化物酶(POD)活性[14],超氧化物歧化酶(SOD)的活性测定采用SOD试剂盒(南京建成生物公司),丙二醛含量采用硫代巴比妥法[15]。采用考马斯亮蓝染色法测定植株的可溶性蛋白[16]。脯氨酸采用磺基水杨酸和茚三酮的方法测定[16]。超氧自由基的产生速率参考文献[17]的方法测定,过氧化氢含量参考文献[18]的方法测定。
1.5 数据分析
数据分析采用SPSS 16.0软件统计处理,运用Duncan’s检验法进行多重比较。
2 结果与分析
2.1 叶片内MDA、脯氨酸和可溶性蛋白含量变化
丙二醛是植物细胞膜脂过氧化的终产物,因而常被当作了解逆境对生物膜系统伤害程度的重要指标[19]。由图2可知,自然降温条件下,不同三角梅品种的MDA含量变化差异十分显著。‘巴西紫’、‘绿樱’的变化趋势相似,其MDA含量均随着温度变化出现先升高后降低又升高的趋势;而‘柠檬黄’、‘中国红’和‘云南紫’的MDA含量均在温度急速降低时显著上升。不同品种三角梅叶片脯氨酸含量随着自然降温的变化趋势差异十分显著。‘柠檬黄’的脯氨酸含量表现为随着温度降低而逐渐降低;‘绿樱’和‘巴西紫’表现为随着自然温度变化出现先降低后升高的现象;‘云南紫’、‘花叶大红’和‘珍珠白’的脯氨酸含量表现为先降低后升高再降低的变化趋势。不同三角梅品种可溶性蛋白的含量也显著不同。‘巴西紫’和‘绿樱’的可溶性蛋白含量随着自然温度变化先升高后降低;‘花叶大红’、‘珍珠白’和‘柠檬黄’的可溶性蛋白含量随着温度变化表现为先升高后降低再升高的变化趋势。

图2 自然降温条件下不同品种三角梅的MDA、脯氨酸和可溶性蛋白含量的变化
2.2 叶片H2O2含量和超氧自由基产生速率(O2·-)变化
H2O2和O2·-是植物体内常见的活性氧,广泛地存在于线粒体、叶绿体以及酶促反应中。如图3所示,三角梅越冬期间,不同品种之间的O2·-产生速率存在显著差异,‘巴西紫’、‘珍珠白’和‘柠檬黄’的O2·-产生速率随着自然降温先升高后降低;而‘中国红’的O2·-产生速率则随着自然降温逐渐升高;‘花叶大红’和‘绿樱’的O2·-产生速率则表现为随着自然降温先降低后升高再降低然后又升高的趋势。不同三角梅品种间的H2O2含量在自然降温下的变化趋势也存在显著差异。‘巴西紫’、‘花叶大红’、‘绿樱’和‘云南紫’的H2O2含量在越冬期间均随着温度降低出现先升高后降低再升高然后又降低的变化趋势;而‘柠檬黄’和‘中国红’则表现出先降低后升高再降低的变化趋势。

图3 自然降温条件下不同品种三角梅的超氧自由基产生速率(O2·-)和过氧化氢(H2O2)含量的变化
2.3 叶片POD、CAT和SOD活性变化
CAT、SOD和POD是植物体内主要的抗氧化酶,在维持细胞内活性氧的动态平衡过程中起十分重要的作用。如图4所示,在三角梅越冬期间,‘巴西紫’、‘花叶大红’和‘绿樱’的POD活性表现为先升高后降低再升高的变化趋势;‘云南紫’的POD活性则是先升高后降低的趋势;‘中国红’的POD活性表现为随着温度的降低而逐渐降低。‘绿樱’和‘柠檬黄’的CAT活性随着越冬期间的温度变化表现为先升高后降低的变化趋势;‘花叶大红’和‘中国红’的变化趋势表现为先降低后升高再降低的变化趋势。‘巴西紫’的SOD活性变化在越冬期间表现为先升高后降低的变化趋势,‘绿樱’的SOD活性的变化趋势则与之相反,表现为先降低后升高;‘珍珠白’、‘柠檬黄’和‘中国红’的SOD活性在越冬期间随着温度变化则表现为先降低后升高再降低的变化趋势。
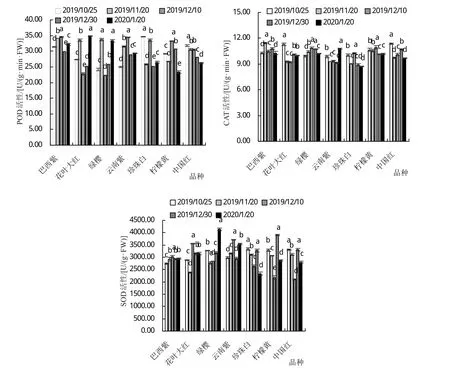
图4 自然降温条件下不同品种三角梅的POD、CAT和SOD活性的变化
3 结论
本研究对不同三角梅品种温州地区越冬期间的生理生化进行研究,结果表明不同的三角梅品种间的低温适应力存在极大的差异,‘中国红’、‘巴西紫’和‘柠檬黄’表现出较强的低温适应能力;‘绿樱’、‘云南紫’低温抵抗能力稍弱,但也正常完成越冬;而‘花叶大红’和‘珍珠白’适应低温能力较差,随着温州地区冬季温度降低,可溶性蛋白含量降低,代谢越来越弱,可能会通过休眠来完成越冬活动,但仍需进一步验证。
4 讨论
低温显著影响植物的生长发育,也是限制植物观赏价值及花卉品质的重要环境因素之一[20],特别是对低温极为敏感的原产于热带和亚热带地区的观赏植物,低温极大地限制了其推广和产业化发展。植物本身通常有一定的适应逆境的能力,但不同物种间差异较大,而同一物种的不同品种之间也存在显著不同[21]。本研究发现,不同品种的三角梅在相同的自然条件下越冬表现差异极大,表明三角梅不同品种间的低温适应能力显著不同,这可能与品种本身的生物学特性或者其内部的生理生化及分子的调节能力有关。本研究发现,在越冬期间,随着温度变化,‘巴西紫’、‘柠檬黄’的MDA含量和O2·-的产生速率虽有上调但依然维持在较为稳定的水平,且随着温度的变化其可溶性蛋白的含量及POD、CAT活性均出现了较大幅度的上升,表明‘巴西紫’在越冬期间可通过体内正常的代谢活动及抗氧化酶活动的上调来维持自身内环境的稳定,完成正常的越冬过程[22-23]。
植物体内的脯氨酸通常被认为是渗透调节小分子,而可溶性蛋白则包含了植物体内各种酶促反应过程的酶和小分子,在某种程度上体现了植物体内的生理活跃水平[24-25]。本研究发现‘云南紫’和‘绿樱’在越冬期间虽然表现为O2·-产生速率的显著上调,但其可溶性蛋白和脯氨酸含量及抗氧化酶POD和SOD活性维持了较高的水平,表明‘绿樱’和‘云南紫’在温州地区也具有较为良好的适应能力,可以正常完成越冬。然而,‘花叶大红’和‘珍珠白’则表现较低的脯氨酸、可溶性蛋白含量,较高的O2·-的产生速率和H2O2含量,且‘珍珠白’的抗氧化酶SOD、CAT和POD活性在后期表现出了明显的下降趋势,表明‘花叶大红’和‘珍珠白’的低温适应能力稍弱于其他品种,具体表现为可溶性蛋白含量降低,代谢越来越弱,因此可能会通过休眠来减少养分消耗以完成越冬活动,但仍需进一步验证。而‘中国红’的MDA、脯氨酸、可溶性蛋白、H2O2含量和O2·-的产生速率均表现为虽有变化但幅度较小,整体表现为较为稳定的状态,表明‘中国红’在越冬期间植物自身的内环境较为稳定,对自然温度的变化并不是极为敏感,具有较强的低温适应能力。
不同三角梅品种在温州地区越冬期间表现出不同的适应性,主要体现在完全不同的MDA、脯氨酸含量、O2·-的产生速率、H2O2含量及抗氧化酶 SOD、CAT 和POD活性的变化趋势,因此本研究从生理生化角度分析三角梅品种的越冬能力存在显著品种差异性,这可能是由品种间的生理特异性所引起的,也可能是低温环境与遗传角度共同作用的结果。因此,仍需从分子及遗传角度进一步研究不同三角梅品种对在低温环境的适应性机制。
