夜间人工照明对两种常见城市杂草光合特性及生长的影响
2021-03-22王欣宇
黄 莉,王欣宇,陈 斌
夜间人工照明对两种常见城市杂草光合特性及生长的影响
黄 莉,王欣宇,陈 斌*
(南京林业大学南方现代林业协同创新中心,南京林业大学生物与环境学院,南京 210037)
为明晰城市夜间人工照明对于植物光合作用特征和生长表现的调控作用,以典型城市杂草牛筋草和蒲公英为对象,研究在夜间灯光(光合有效辐射约为1 μmol·m-2·s-1的路灯落地面光强水平)和无夜间灯光两种处理条件下植物的光合生理和生物量积累。结果表明:(1)牛筋草和蒲公英叶片在夜间低水平光照条件下仍可进行光合作用。此外,夜间灯光对牛筋草叶片的夜间胞间CO2浓度、蒸腾速率和气孔导度具有抑制作用,而对蒲公英的相应指标无影响。(2)夜间灯光可显著抑制牛筋草叶片的白天净光合速率,显著抑制蒲公英叶片的白天胞间CO2浓度。(3)总体而言,夜间灯光对牛筋草的生物量积累无显著影响,但能显著增加蒲公英的地上生物量和总生物量水平。研究结果表明夜间人工照明可通过调节植物的昼、夜光合特征影响植物的生长速率,且植物对夜间光照的响应具有种间差异性。
夜间灯光;牛筋草;蒲公英;光合特性;生长
光不仅能为植物光合作用提供能量,也参与调节植物生长发育和开花结果等形态建成,并控制其生长过程[1]。与其他信号相比,光照的昼夜节律是地球历史上最为稳定的环境信号[2]。然而,自人类进入工业社会后,随着电力的广泛应用,城市夜间亮化工程飞速发展,人类聚集地及其周边的昼夜光照格局发生巨大改变。过去60年来,全球人造光源以约6%的年均速度持续增加[3]。尽管全球已有近五分之一的陆地受到人工光照的影响,但人们对夜间人工光照生态学效应的认识较为有限[4]。
到目前为止,关于夜间光照如何影响植物生长的研究主要集中在园艺学领域,核心关注高强度夜间光照对园艺(或农业)作物的促生长作用[5-6]。相对园艺用高强度人工光源(光合有效辐射范围约为300 ~ 600 μmol·m-2·s-1[7-8]),室外夜间照明用的人工光源的光强水平十分有限。例如,街道高压钠灯落地照度区间为32 ~ 35 Lx,LED灯落地照度区间为15 ~ 20 Lx[9];城市行道树夜晚所受光强普遍为1~10 μmol·m-2·s-1[10]。有关此类低水平夜间光照的生态学效应的研究较为缺乏,只有有限的研究案例报道:种植于道路两侧路灯正下方的香樟()行道树较种植于相邻路灯之间的香樟具有更高的生长率[11];路灯照射亦可促进杏树()新枝生长量的增加[12]。夜间照明还可改变草本植物群落中优势物种的群落生物量占比[13],甚至促进外来植物的入侵过程[14]。值得注意的是,上述研究案例主要关注植物形态结构和生物量积累的响应特征,而作为影响植物生长和生物量积累的关键生理性状——光合作用,其对夜间人工照明的生理生态学响应特征尚缺乏研究。
鉴于此,本研究分别选取常见城市杂草牛筋草()和蒲公英()开展模拟城市夜间人工照明实验,分析夜间灯光和无夜间灯光两种光照处理条件下植物昼、夜光合生理指标及生物量积累的差异性,旨在揭示弱光型城市夜间照明对于植物光合作用和生长表现的调控作用。
1 材料与方法
1.1 试验材料与设计
1.1.1 供试材料 牛筋草为禾本科穇属植物,蒲公英为菊科蒲公英属植物,两者在我国分布广泛,皆为常见的城市杂草,多生长于草地和路边[15]。
1.1.2 试验设计 2020年5月中旬对牛筋草和蒲公英进行育苗。育苗时选取大小均一、颗粒饱满的种子,用10%的次氯酸钠溶液浸泡消毒5 min后,用无菌水充分冲洗。洗净后的种子点播于育苗盘上,置于22 ℃恒温培养室进行黑暗处理。待种子出苗3 d后,对2种植物分别足量选取大小一致的健康小苗移入穴盘。2周后再挑选长势一致且健康的幼苗,以1盆1株的方式移入花盆(上直径12.0 cm、下直径8.6 cm、高10.8 cm,栽培基质为蛭石),并转移至室外半开放温室(仅顶部有透明塑料板)开展盆栽控制试验。
控制试验分为无夜间灯光(CK)和夜间灯光(NL)两种处理,每种植物每个处理设置12个重复。白天利用自然光源(阳光);夜晚,对于NL处理下的植物使用LED灯带作为夜间人工光源(光谱信息如图1所示)。利用HR-350光谱分析仪(中国台湾Hipoint公司),通过调节LED灯带的高度与密度,使花盆处夜间光照保持在强度约为60 Lx,光合有效辐射(PAR)约为1 μmol·m-2·s-1的水平。通过在温室顶部外侧设置光照传感器,自动控制夜间光照的开关,以模拟城市照明路灯的运行时间周期。具体为,当环境光强低于50 Lx时,照明自动开启;当环境光强高于此临界值时,照明自动关闭。2种处理间通过设置遮光帘以避免NL处理对CK处理产生干扰。试验过程中,每天早晚对植物进行适当浇水,保证培养基质始终保持湿润状态。通过每周对每棵植物浇灌50 mL 30%标准强度的霍格兰营养液(Hoagland solution,青岛海博生物技术有限公司),提供植物生长的必须养分。试验期间,每晚进行光照处理,运行2个月后,收获植物。

图1 LED光源的光谱组成
Figure 1 Light profile of photosynthetic photon flux density (PPFD) between 400 and 750 nm of LED lights
1.2 指标测定
1.2.1 植物叶片白天快速光响应曲线的测定 试验运行40 d后,对于每种植物的每个处理组各随机选取5个重复(即5株植物)进行光合指标的测量。具体而言,对选定的每株植物,挑选叶龄叶位(牛筋草自上而下第5片叶,蒲公英自内而外第4片叶)一致的健康叶片,在天气晴朗的上午(9:00—11:30)利用Li-6800(美国Li-cor公司)测定光响应曲线。测量过程中的相关参数设置分别为:气体流速500 μmol·s-1,叶室中CO2浓度400 μmol·mol-1,空气相对湿度55%,叶片温度27 ℃。正式测量前采用1 800 μmol · m-2· s-1光强诱导植物叶片30 min。正式测量时采用自动程序测量,设置快速光响应曲线的光合有效辐射(PAR)强度分别为1 800、1 500、1 200、900、700、500、400、300、200、175、150、125、100、75、50、25和0 μmol·m-2·s-1共17个梯度,不同光强值设置数据采集时间为120 ~150 s。光合测定仪自动记录净光合速率、气孔导度、胞间CO2浓度、蒸腾速率等光合参数。
1.2.2 夜间植物光合参数的测定 在测定白天光曲线当天的夜间,对植物进行夜间光合指标的测量。具体而言,为确保露天环境完全进入黑暗状态(即对照组环境光强为0 μmol·m-2·s-1),测量工作于21:00后进行。选取用于白天测量光曲线的植物叶片,利用Li-6800测定植物夜间光合。测量过程中的相关参数设置分别为:气体流速500 μmol·s-1,叶室中CO2浓度400 μmol·mol-1,空气相对湿度55%,叶片温度27 ℃。植物叶片在正式测量前,先于夜间环境光强下诱导至少20 min,待净光合速率和气孔导度数值稳定后开始记录。每个叶片记录5次,每次间隔1 min,计算平均值作为这一叶片的相关光合指标数据。
1.2.3 植物生物量的测定 试验处理60 d后对植物进行收获,收获时将植物按地上和地下部分分类收集,并将植物根系洗净,一起置于65 ℃烘箱烘干72 h,称量干重。
1.3 数据分析
对每种植物在白天和夜晚的光合参数以及生物量指标,采用检验进行NL和CK处理组的比较分析。此外,根据Ye等[16]的直角双曲线修正模型对两种植物的白天光响应曲线进行方程拟合和计算,得出相应的光合特征参数,采用检验开展NL和CK处理间的差异性分析。其模型的数学表达式为:

式中,是光响应曲线的初始斜率,和为系数,为光合有效辐射,d为暗呼吸。数理统计分析用SPSS 22.0。
2 结果与分析
2.1 夜间光照对植物白天光合特征的影响
与CK相比,NL处理可显著抑制牛筋草在PAR为175和300 ~ 900 μmol·m-2·s-1时的净光合速率(图2(a)),但可显著促进蒲公英在PAR为25 μmol·m-2·s-1时的净光合速率。值得注意的是,当> 700 μmol·m-2· s-1时,NL处理下的蒲公英净光合速率在均值水平上低于CK处理下的个体(图2(b))。
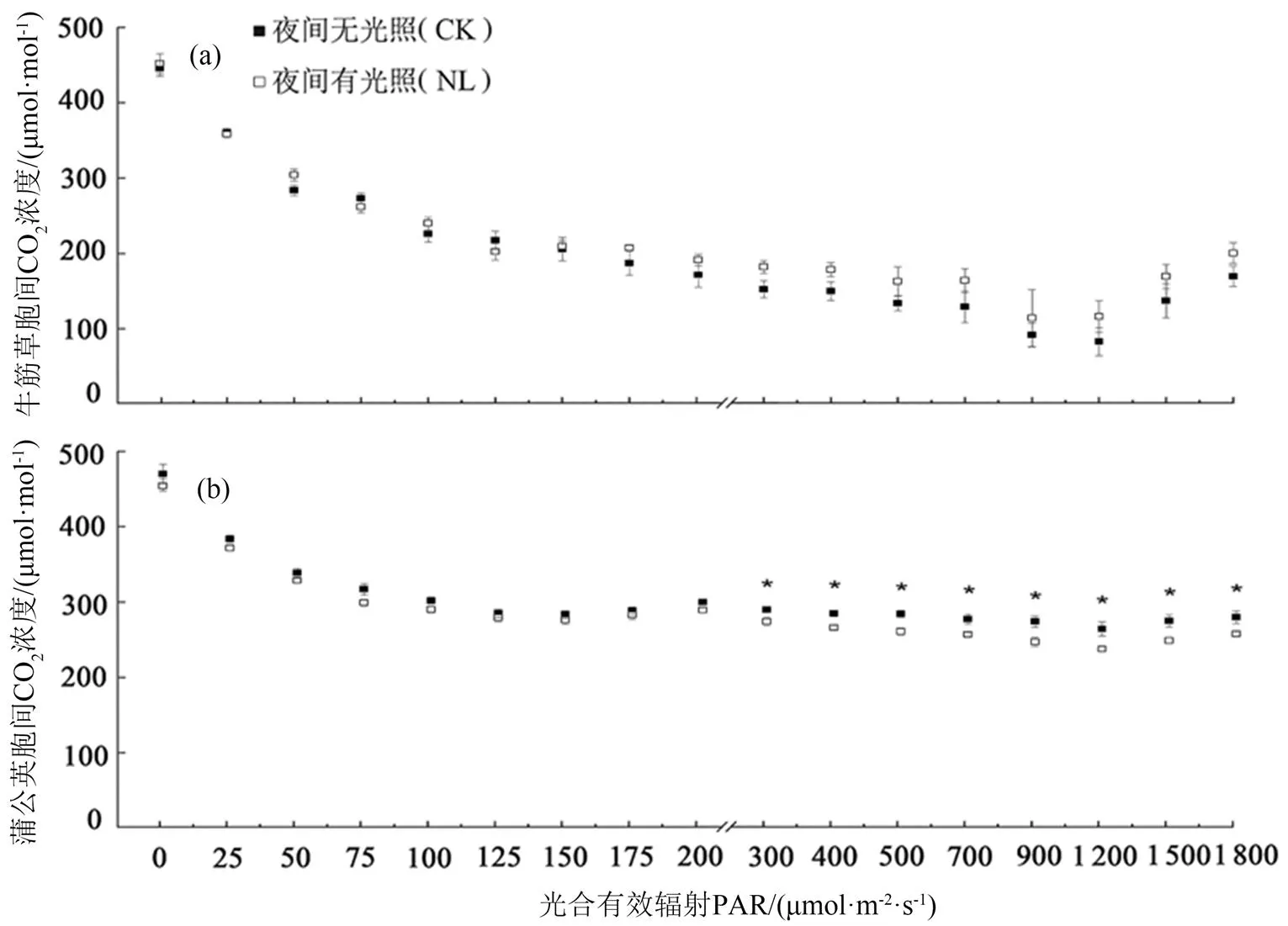
与CK相比,当> 300 μmol·m-2·s-1时,NL处理下的牛筋草胞间CO2浓度在均值水平上高于CK处理下的个体(图3(a))。另一方面,在>300 μmol·m-2·s-1时NL处理下的蒲公英叶片胞间CO2浓度显著下降(图3(b))。此外,与CK相比,NL处理对牛筋草和蒲公英的气孔导度和蒸腾速率均无显著作用。

注:* P < 0.05。误差线采用标准误表示;n = 20(2物种 × 2处理 × 5植株)。
Figure 2 Effect of artificial lighting at night on the responses of daytime net photosynthetic rate (n) of (a)and (b)leaves to the strength of photosynthetically active radiation (PAR)
注:* P < 0.05。误差线采用标准误表示;n = 20(2物种 × 2处理 × 5植株)。
Figure 3 Effect of artificial lighting at night on the responses of daytime intercellular CO2concentration (i) of (a)and (b)leaves to the strength of photosynthetically active radiation (PAR)

表1 夜间人工光照对牛筋草和蒲公英白天光响应曲线参数的影响
注:不同字母表示种内组间数据具有显著差异(< 0.05);数据采用均值±标准误表示;= 20(2物种 × 2处理 × 5植株)。

注:*代表组间存在显著性差异(P< 0.05);误差线采用标准误表示;n = 20(2物种 × 2处理 × 5植株)。(a)净光合速率Pn;(b)胞间CO2浓度Ci;(c)蒸腾速率Tr;(d)气孔导度Gs。
Figure 4 Effects of artificial light at night on the night-time photosynthetic parameters ofand
2.2 夜间光照对植物白天光响应曲线参数的影响
如表1所示,两种植物在NL与CK处理下的光响应曲线拟合程度的决定系数均大于0.99。与CK相比,NL可显著降低牛筋草的最大净光合速率,但对其光饱和点、光补偿点、初始斜率和暗呼吸速率无显著影响;另一方面,NL可显著增加蒲公英的初始斜率,显著降低它的光补偿点、光饱和点和暗呼吸速率,但对其最大净光合速率无显著影响。

注:不同字母代表植物的地上(或地下)生物量存在组间显著差异;*,总生物量有显著差异;ns,总生物量差异不显著;数据采用均值±标准误表示;n = 48(2物种 × 2处理 × 12植株)。
Figure 5 Effects of artificial light on the biomass ofand
2.3 夜间光照对植物夜间光合生理指标的影响
与CK相比,NL处理可诱导牛筋草和蒲公英积极响应夜间光照,触发夜间光合作用(图4(a))。此外,NL能显著抑制牛筋草的胞间CO2浓度(图4(b))、蒸腾速率(图4(c))和气孔导度(图4(d));但对蒲公英的相应指标无显著影响。
2.4 夜间光照对植物生物量的影响
牛筋草的地上、地下及总生物量在CK和NL处理间均无显著差异。而蒲公英的地上生物量和总生物量在NL处理下显著增加,其地下生物量也有增加的趋势(= 0.068)(图5)。
3 讨论
园艺工作中,常在夜间利用高强度灯光(如200 μmol·m-2·s-1)[21]给植物补充光能以增加植物光合作用时长,提高其生长速率。但对于室外照明用的夜间人工光源(如,路灯)而言,其落地照度往往处于极低水平[9]。传统认为此类夜间弱光照生态效应的主要作用机制是在光信号传递,而非光能量供给方面[13]。例如,PAR仅为0.02 μmol·m-2·s-1水平的夜间光照能延迟菊花()的开花时间[18]。然而,近年来有研究发现城市夜间人工照明亦能诱导紫丁香()[19]和二球悬铃木()[9]进行夜间光合作用,甚至使它们的净光合速率均达到正值。这可能是因为夜间光照辐射影响了植物气孔的开闭,叶片气孔限制值减小,气孔导度增加,导致叶片的气孔阻力减小,气孔开张度增大,环境中更多CO2分子能通过扩散进入叶片光合组织,使得叶片内胞间CO2浓度上升,光合作用提高[19]。当然,值得注意的是,上述研究的观测对象均为具有一定高度的木本植物,它们的叶片高度与近地面位置相比更加接近路灯光源,接收到的路灯光照强度更高。然而,本研究结果似乎表明牛筋草和蒲公英在PAR仅为1 μmol·m-2·s-1水平的弱夜间光照下即可进行光合作用。对这一结果的可能解释是夜间弱度光照抑制了植物的暗呼吸速率[20]。
本研究结果同时表明,夜间照明对植物生长的调控不仅仅局限于对植物夜间生理活动的影响,更重要地反映在它对植物白天光合生理活动的影响。已有大量园艺学研究表明,对植物进行夜间高强度补光可以影响植物白天的光合作用过程。例如,与夜间无光照处理相比,6 h夜间强光照处理可以显著提高小麦()旗叶在白天的净光合速率[21]。然而,随着夜间强光照时长的增加,烟草()叶片的白天光合参数表现出先升高后降低的趋势[22]。而本研究观察到牛筋草和蒲公英白天净光合速率在不同程度上受夜间弱光照所抑制的现象,这一结果同样也是基于长时间(彻夜)光照的实验处理方式。上述研究结果说明,过长的夜间光照时间可能是抑制植物叶片白天光合机能的重要原因[22]。
大量园艺研究已明确,夜间强光照可提高植物生物量水平[23-24]。目前,在关于夜间弱光照的有限研究中亦发现了类似效应。如陈良华等[25]发现夜间路灯照射可显著增加凤仙花()的叶、茎干重和夏菊()的茎干重。本研究发现在夜间弱光照处理下,即使牛筋草的白天光合机能受到显著抑制,其生物量积累并未显著降低,而蒲公英在白天光合机能未受明显影响的条件下,其生物量积累水平依然能够显著增加。产生这一现象的可能原因之一是由于植物夜间进行微弱光合作用(或夜间呼吸受抑制)的补偿效应所致。此外,夜间光照也可能通过改变植物昼夜节律[25]影响植物个体生长发育过程,进而对其生物量积累产生作用。
牛筋草和蒲公英响应夜间光照处理的种间差异性暗示了夜间人工照明可能对植物群落的结构与动态具有潜在影响。Bennie等[13]对英国康沃尔郡野外典型草本植物群落的控制实验发现,夜间人工照明可以通过改变群落中细弱剪股颖()、黄花茅()和绒毛草()3个优势种的物候特征、生物量水平及其在群落中的生物量占比,进而改变这些优势种在群落中的相对盖度,最终引起整个草本植物群落结构组成的变化。Speier等[14]在研究外来植物入侵本土植物群落的实验中发现,入侵植物对夜间光照反应敏感,在夜间光照条件下更容易入侵:夜间人工照明可以通过改变入侵植物狭叶一枝黄花()和反枝苋()等与本土植物长叶车前()和草地早熟禾()等之间的竞争平衡,进而改变外来—本土植物群落中的物种生物量占比,从而影响整个植物群落结构组成。本研究在植物生理生态水平上为上述研究结果的解释提供了一个可能机制,即夜间光照可通过影响植物的昼夜光合特征进而改变植物的生长速率,从而影响植物群落中的种间竞争关系。
[1] SVECHKINA A, PORTNOV B A, TROP T. The impact of artificial light at night on human and ecosystem health: a systematic literature review[J]. Landsc Ecol, 2020, 35(8): 1725-1742.
[2] RICH C, LONGCORE T. Ecological Consequences of Artificial Night Lighting[M]. Washington DC: Island Press, 2006.
[3] SMITH M. Time to turn off the lights[J]. Nature, 2009, 457(7225): 27.
[4] GASTON K J, VISSER M E, HÖLKER F. The biological impacts of artificial light at night: the research challenge[J]. Phil Trans R Soc B, 2015, 370(1667): 20140133.
[5] ISLAM N, PATIL G G, GISLERØD H R. Effect of photoperiod and light integral on flowering and growth of(Raf.) Shinn[J]. Sci Hortic, 2005, 103(4): 441-451.
[6] 李海云, 韩国徽, 任秋萍, 等. 不同光周期对黄瓜幼苗生长的影响[J]. 西北农业学报, 2009, 18(3): 201-203.
[7] FU W G, LI P P, WU Y Y. Effects of different light intensities on chlorophyll fluorescence characteristics and yield in lettuce[J]. Sci Hortic, 2012, 135: 45-51.
[8] 丁娟娟, 杨振超, 王鹏勃, 等. LED光强对不结球小白菜生长与光合特性的影响[J]. 西北农林科技大学学报(自然科学版), 2015, 43(3): 113-118.
[9] 王旭东, 许晶, 徐慧, 等. 城市道路夜间照明与行道树相互影响探析[J]. 中国园林, 2019, 35(9): 120-123.
[10] 陈芳, 彭少麟. 城市夜晚光污染对行道树的影响[J]. 生态环境学报, 2013, 22(7): 1193-1198.
[11] 李小婷, 陈斌, 王海军, 等. 城市夜间灯光对香樟生长的影响[J]. 应用生态学报, 2019, 30(7): 2284-2290.
[12] 范春楠, 孙艳昭, 王艺萌, 等. 路灯照明对杏树行道树生长及光合的影响[J]. 北华大学学报(自然科学版), 2017, 18(1): 101-105
[13] BENNIE J, DAVIES T W, CRUSE D, et al. Artificial light at night alters grassland vegetation species composition and phenology[J]. J Appl Ecol, 2018, 55(1): 442-450.
[14] SPEIER B, LIU Y J, VAN KLEUNEN M. Biomass responses of widely and less-widely naturalized alien plants to artificial light at night[J]. J Ecol, 2021, 109(4): 1819-1827.
[15] 中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京: 科学出版社, 1959.
[16] YE Z P, ROBAKOWSKI P, SUGGETT D J. A mechanistic model for the light response of photosynthetic electron transport rate based on light harvesting properties of photosynthetic pigment molecules[J]. Planta, 2013, 237(3): 837-847.
[17] 马艳, 喻晨, 王瑞, 等. 不同LED补光时间对日光温室番茄生长发育及光合特性的影响[J]. 新疆农业科学, 2019, 56(8): 1469-1475.
[18] VINCE P D, CANHAM A E. Encyclopedia of Plant Physiology[M]. Berlin: Springer-Verlag, 1983.
[19] 高明辉, 吴汇元, 周兆平, 等. 夜间照明对紫丁香光合作用的影响[J]. 吉林林业科技, 2012, 41(5): 7-10, 22.
[20] CHEN Z Y, PENG Z S, YANG J, et al. A mathematical model for describing light-response curves inL[J]. Photosynthetica, 2011, 49(3): 467-471.
[21] 冉午玲, 郭家萌, 马野, 等. 夜间补光对小麦叶片激素含量及光合特性的影响[J]. 麦类作物学报, 2017, 37(9): 1181-1186.
[22] 徐超华, 李军营, 崔明昆, 等. 延长光照时间对烟草叶片生长发育及光合特性的影响[J]. 西北植物学报, 2013, 33(4): 763-770.
[23] KIM Y J, LEE H J, KIM K S. Night interruption promotes vegetative growth and flowering of[J]. Sci Hortic, 2011, 130(4): 887-893.
[24] 刘淑艳, 于振良, 陶延怀, 等. LED补光对番茄幼苗生长的影响[J]. 北方园艺, 2013(23): 58-60.
[25] 陈良华, 曹艺, 杨万勤, 等. 三种园林植物对夜间光照的响应与适应特征[J]. 生态学报, 2017, 37(2): 549-556.
Effects of artificial lighting at night on the photosynthesis and growth of two typical urban weed species
HUANG Li, WANG Xinyu, CHEN Bin
(Co-Innovation Center for the Sustainable Forestry in Southern China, Nanjing Forestry University, College of Biology and the Environment, Nanjing Forestry University, Nanjing 210037)
To investigate the effects of urban artificial lighting at night (ALAN) on the photosynthesis and growth of plants, we conducted a controlled experiment that two typical urban weed speciesandgrew under nighttime lighting (NL, with PAR around 1 μmol·m-2·s-1to simulate the ground-level light intensity of street lamps) and no nighttime lighting (CK) conditions, and measured their photosynthetic parameters and biomass. The results showed that: (1) Bothandcan photosynthesize under low-light NL treatment at night. Meanwhile, NL significantly reduced the nighttime intercellular CO2concentration, transpiration rate and stomata conductance of, but it had no effects on those parameters of. (2) NL treatment significantly reduced the daytime photosynthetic activity ofand significantly reduced the daytime intercellular CO2concentration of. (3) Overall, NL treatment had no effect on the biomass production of, while it significantly increased the shoot mass and total mass of. The results suggested that ALAN can influence the growth of plants via its effects on both day- and night-time photosynthesis of plants, and the responses of plants to ALAN are highly species-specific.
ALAN;;; photosynthesis; growth
S688.4
A
1672-352X (2021)06-0934-06
10.13610/j.cnki.1672-352x.20220106.014
2022-1-7 7:29:38
[URL] https://kns.cnki.net/kcms/detail/34.1162.S.20220106.1254.028.html
2021-03-17
国家自然科学基金项目(32071526)和江苏省高校优势学科建设工程项目(PAPD)资助。
黄 莉,硕士研究生。E-mail:954047670@qq.com
通信作者:陈 斌,副教授。E-mail:bin.chen@njfu.edu.cn
