濒危植物大别山五针松枝叶NPK化学计量特征季节动态
2021-03-22闫景明张世航
闫景明 张世航 汪 漫 杭 伟 钱 申 邱 东 陶 冶*
(1. 安庆师范大学生命科学学院/皖西南生物多样性研究与利用安徽省重点实验室,安庆 246133;2. 中国科学院新疆生态与地理研究所,荒漠与绿洲生态国家重点实验室,乌鲁木齐 830011)
生态化学计量学为研究生物体内的化学元素平衡提供了重要的手段[1],在反映植物器官的内稳定性、养分利用的高低以及限制性元素判断等方面具有重要的作用[2~3]。生态化学计量学中各元素具有不同的意义,其中C 在植物体中起结构性元素的作用,N、P 对判断植物生产力的限制因素起到重要的指示作用[4~5]。N、P含量存在明显的植物系统发育阶段(针阔叶)和功能群(乔灌草)间的差异性[6],而季节变化多强烈反映在叶片上。此外,养分元素间的异速生长关系能反映植物体不同元素之间及不同器官之间养分获取与分配速率的差异性,且异速生长指数也是衡量不同植物功能群养分利用趋同与趋异的重要指标[7]。就同一植物而言,研究植物叶片与不同器官N、P、K 计量特征的季节变化及其异速生长关系对理解植物的养分利用策略和生存机制具有重要意义[8~10]。
植物末端小枝(包括支撑构件枝和光合构件叶)是植物最活跃的器官,能反映植物生长过程中光合产物及养分资源分配策略[11]。植物叶片通常比枝具有更高的养分含量,以支持叶片高的生理活性。落叶阔叶植物枝叶化学计量特征在不同生长期(季节)有显著变化,其叶片N、P 含量一般随植物生长发育而逐渐降低,这在落叶裸子植物华北落叶松(Larix principis-rupprechtii)叶N、P 含量变化中也发现了类似趋势[12]。相比之下,季节变化对常绿植物叶片化学计量特征的影响较弱[13],如祁连圆柏(Sabina przewalskii)叶N、P化学计量特征在海拔上的变异明显大于季节[14],侧柏(Platycladus orientalis)人工林不同植物器官对C、N、P 及其化学计量比均有显著影响,而季节仅对N和P有显著影响[15]。但常绿针叶裸子植物是否也具有与上述柏科植物(磷刺型叶)类似的特征,目前尚不清楚。
大别山五针松(Pinus dabeshanensis)是中国特有物种,仅分布于安徽省岳西、金寨等大别山局地,属国家二级珍稀濒危保护植物。学者们在大别山五针松花粉传播与萌发、种子特性、种内种间关系、群落结构及人工培育等方面开展了一系列研究[16~18],但对其末端小枝N、P、K 化学计量特征及其季节动态还缺乏了解。本研究以大别山五针松末端小枝的枝和叶为研究对象,通过12次(每月1 次)连续调查取样:①分析大别山五针松枝和叶N、P、K 含量及其计量比的差异性及其季节变化特征;②揭示枝和叶N、P、K 化学计量的异速生长关系;③探究降水和温度对大别山五针松枝、叶化学计量特征的影响。研究结果将有助于从化学计量学角度了解该物种的生长特点和养分利用策略,为进一步揭示其濒危机制提供科学依据。
1 研究方法
1.1 研究区概况
研究地点位于大别山腹地金寨县天堂寨镇马鬃岭(115.69°E,31.28°N,海拔1 130 m)。该地区属于北亚热带湿润气候区北缘,年降水量1 350~1 400 mm,年平均气温14~15℃,无霜期212 d。土壤为山地棕壤,pH5.0~5.5,呈酸性。植物资源丰富,区系起源古老,地理成分复杂,珍稀濒危和特有植物丰富。本地区特有的大别山五针松是中国二级珍稀濒危保护植物,其自然种群稀少,且主要分布于阴坡近山脊处[18]。
1.2 样品采集与处理
在多年野外调查及资料收集的基础上,选择交通便利的金寨县天堂寨镇马鬃岭作为研究地点。设置30 m×30 m 固定样地,选定6 株长势良好、大小均一的野生大别山五针松成株作为标准木(胸径20~27 cm)。样品采集时间从2017 年12月15 日开始至2018 年11 月15 日止,每月15 日左右采集。用高枝剪在每棵树的树冠中部按东南西北4 个方向采集4 个完整小枝,装入信封做好标记后放入装有冰袋的泡沫保温箱内,尽快运回实验室。同时采集0~10 cm混合土壤样品。
在实验室内将枝条冲洗干净并将枝、叶分离,分别装入信封,置于80℃烘箱内烘干24 h。野外观察及室内处理时发现,该地区大别山五针松新叶萌生量极少,甚至有些植株的枝条无新生叶。因此,本研究没有考虑针叶年龄差异的影响,即在样品采集过程中不区分新叶与老叶(新叶生物量比例0~20%),将其混合后使用。烘干后的样品经盘式震动研磨仪(RS200)研磨成粉末后装入封口袋避光保存。
1.3 植物和土壤元素含量测定
大别山五针松枝和叶全N 含量通过碳氮分析仪(multi N/C 3100,耶拿,德国)测定,全P 采用钼锑抗比色法测定,全K 采用火焰光度计法测定[19]。最后计算N∶P、N∶K和P∶K。采用土壤农化分析常规方法[19],测定土壤有机质,全量氮、磷、钾,速效氮、磷、钾含量,及pH和电导率(见表1)。
1.4 地理地形因子及气象数据收集与处理
首次野外调查时,用GPS 定位仪及电子罗盘测量并记录样地坡向、坡度、经度、纬度、海拔等环境参 数。2017 年10 月至2018 年11 月1 年期气 象数据来自县气象局的逐日气象数据,10 年(2009~2018)、30 年(1981~2010)和68 年(1951~2018)气象数据来源于国家气象局网站和中国资源环境数据云平台(见图1)。通过计算整合,选择采样前3、5、7、10、15和30 d(30 d表示1个月)的日平均气温(T3、T5、T7、T10、T15 和T30)和多日累积降水量(P3、P5、P7、P10、P15 和P30),以及10、30 和68 年的月平均气温(T10y、T30y 和T68y)和月平均降水量(P10y、P30y 和P68y)共9 个时间尺度18 个温度和降水量参数作为气象因子,分析其与大别山五针松枝和叶化学计量特征的关系。

表1 大别山五针松样地土壤理化特性Table 1 Soil physical-chemical properties in P.dabeshanensis plot
1.5 统计分析方法
对大别山五针松枝和叶N、P、K含量及N∶P、N∶K和P∶K 进行描述统计分析,对比各参数月平均值及变异系数(CV)的大小。采用双因素方差分析检验器官和月份对化学计量特征的影响作用,随后对不同月份枝和叶元素含量及其化学计量比进行单因素方差分析,用Tukey’s HSD 法进行多重比较(0.05 水平),用T-检验分析各月份不同器官间的元素含量及其化学计量比的差异性。采用Pearson相关性分析探讨不同时间尺度降水量、温度与枝、叶化学计量特征的关系。N、P、K 之间的异速生长关系用幂函数表征:
式中:Y 和X 为任意2 个元素,b 为标准化常数,a 为异速生长指数(a=1 为等速关系,a≠1 为异速关系)。将幂函数对数化后利用RMA 回归计算相关参数、95%置信区间及R2[20],并对比枝和叶之间a的差异性。
2 结果与分析
2.1 大别山五针松枝和叶N、P、K 含量及化学计量比的月动态特征
双因素方差分析表明(见表2),大别山五针松末端小枝N、P、K 及其计量比受器官的影响最大,其次为月份。除N 和N∶P 外,器官与月份对其余4个指标有显著交互效应。
从12 个月的平均值来看,叶N、P、K 含量(10.40±1.97、1.43±0.17 及5.15±1.27 mg·g-1)及N∶P、N∶K、P∶K 平 均 值(7.27±1.23、2.02±0.56 及0.28±0.06)均显著(P<0.05)或极显著(P<0.01)高于枝(见表3)。除枝(26.75%)和叶N∶K(28.00%)的变异系数>25%属于中等变异外,其余指标的变异系数均在25%以下(属于弱变异),其中P 的变异系数最小(叶和枝分别为11.89%和15.08%)。叶和枝的N∶P∶K平均值分别为7∶1∶4和5∶1∶4。
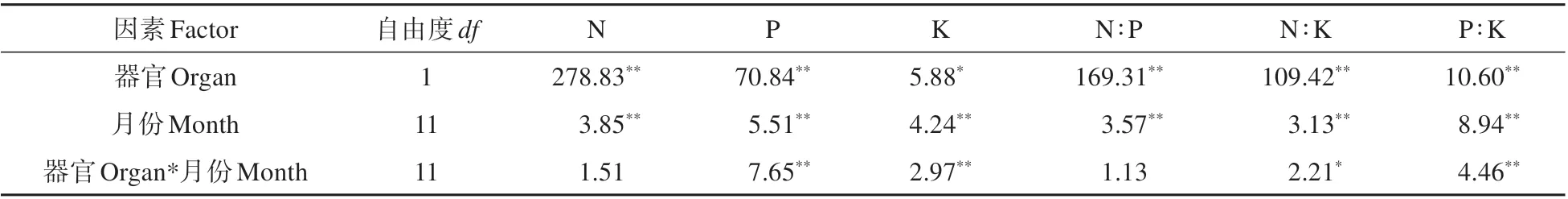
表2 大别山五针松枝叶N、P、K含量及计量比的双因素方差分析(F值)Table 2 Two-way ANOVA of N,P,K contents and the stoichiometric ratios in twigs and needles of P.dabeshanensis(F value)

表3 大别山五针松枝叶N、P、K含量及计量比的年平均值与变异系数Table 3 Annual means and variation coefficients of N,P,K contents and the stoichiometric ratios in twigs and needles of P.dabeshanensis
枝和叶间的T-检验表明(见图2),除个别月份的个别指标外(N,4 月;N∶P,1 月和4 月;N∶K,1月、4 月和11 月),大别山五针松叶N 含量、N∶P 及N∶K 在大部分月份均显著高于枝。除8 月份枝P含量显著高于叶外,叶P 含量在多数月份高于枝。叶和枝K 含量随月份变化的变异幅度较大,互有高低,但大部分表现为叶高于枝。枝和叶P∶K 在前5 个月较为接近,随后5~8 月叶显著高于枝,但10和11月又显著低于枝。
大别山五针松各化学计量指标在12 个月份之间均具有显著差异(P<0.05),但变化趋势不同(见图2)。针叶N 含量在1 年中有3 次先升高趋势,分别在2 月和6 月和10~11 月;叶N 含量的低点分别在1 月、4 月和8~9 月。枝N 含量总体波动性较低,其最高值出现在1 月和11 月,而其余10 个月之间无显著差异。叶P 含量呈“下降—升高—下降—升高”的变化趋势,其含量在8月最低,10~11 月回升。枝P 含量在12 月至次年6 月间小幅波动,但月份间无显著差异;枝P 含量在7 月最低,10 月最高。叶和枝K 含量季节波动较大,尤其是叶(CV=24.66%);叶K 含量2月开始下降,在5 月到达最低点,6 月快速升高,随后波动性下降;枝K 含量在8 月最高,1 月和11 月最低。
枝和叶化学计量比也存在显著的季节波动(见图2)。针叶N∶P 在1 月最低、3 月最高,4 月则快速下降,随后呈现缓慢升高(4~8月)和缓慢下降并稳定的变化过程(8~11月)。枝N∶P的波动性高于针叶,规律性不明显。针叶N∶K 与P∶K 均在5月达到最高值,5 月前后分别为上升和下降过程。除5月份枝和叶P∶K 绝对值差距最大外,枝叶P∶K的变化趋势相似。
2.2 大别山五针松枝和叶化学计量特征的相关性
相关性分析表明(见表4),叶N、P、K之间均存在显著正相关(P<0.05),其中P与K 间相关系数最大(0.50);枝N 与P 及P 与K 之间均存在显著正相关(P<0.05),也是P 与K 之间的相关系数最大(0.49)。叶和枝N∶P 均与N 含量具有最大的相关系数(正相关),即N∶P 主要受到N 元素的调控;同理,枝和叶N∶K、P∶K 主要受到K 元素调控(负相关)。进一步分析发现,相同元素在叶和枝之间均无显著相关性,表现出相对独立性。
2.3 大别山五针松枝和叶N、P、K 含量的异速生长关系
由表5 可知,大别山五针松叶N 与P(N-P1.61,P=0.00)、P 与K(P-K0.52,P=0.00)及N 与K 之间(NK0.84,P=0.01)均具有极显著的异速生长关系,枝N与P(N-P1.19,P=0.00)及P 与K 之间(P-K0.76,P=0.00)也具有呈极显著的异速生长关系,而枝N 与K 的异速生长关系不显著。从异速生长指数来看,叶N 与P 间的异速生长指数显著高于枝,而叶P 与K间的异速生长指数显著低于枝。这表明,大别山五针松叶具有比枝更高的N 分配速率和较低的P分配速率。
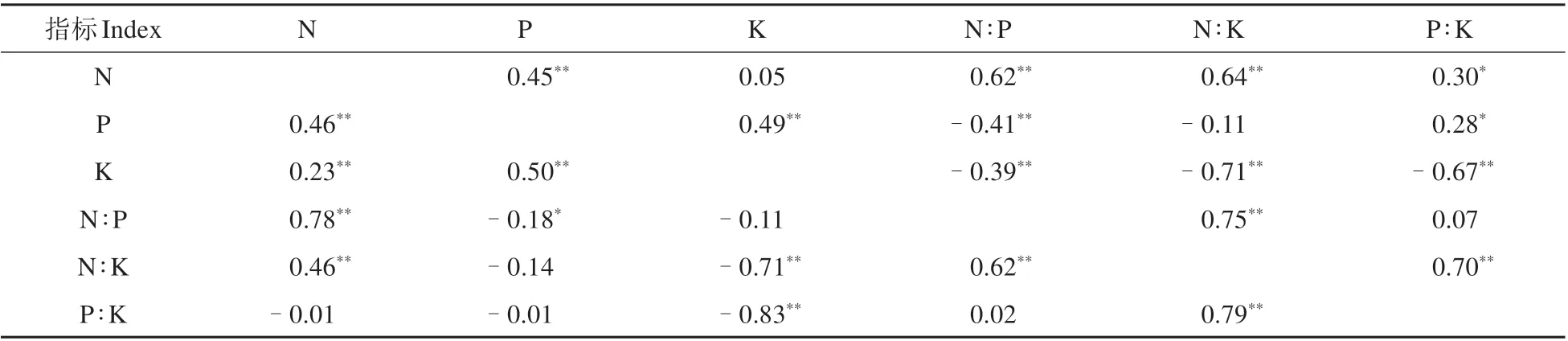
表4 大别山五针松枝叶N、P、K含量及计量比的相关系数Table 4 Correlation coefficients between N,P and K contents and the stoichiometric ratios in twigs and needles of P.dabeshanensis

表5 大别山五针松枝叶N、P、K间的异速生长关系Table 5 Allometric relationships between N,P,K contents in twigs and needles of P.dabeshanensis
2.4 大别山五针松枝和叶化学计量特征与温度和降水的关系
温度和降水与3 个元素含量的相关性分析表明(见表6),3 个时间尺度的降水量(P30、P30y 及P68y)与枝N 含量为显著负相关(P<0.05),除此以外,任何时间尺度的温度和降水参数对叶和枝N含量均无显著影响。除降水量参数P10、P10y 及P68y 与叶P 含量无显著相关外,任何时间尺度的温度和降水量对叶和枝N 含量均呈显著(P<0.05)或极显著(P<0.01)负相关,即温度和降水增加均会显著降低枝和叶P含量。叶K含量与温度无关,但与10、15 及1 个月间隔的降水量呈显著或极显著负相关;枝K 含量与所有时间尺度的温度均呈显著或极显著正相关,而与多年的月均降水量(P10y、P30y及P68y)呈显著或极显著正相关。
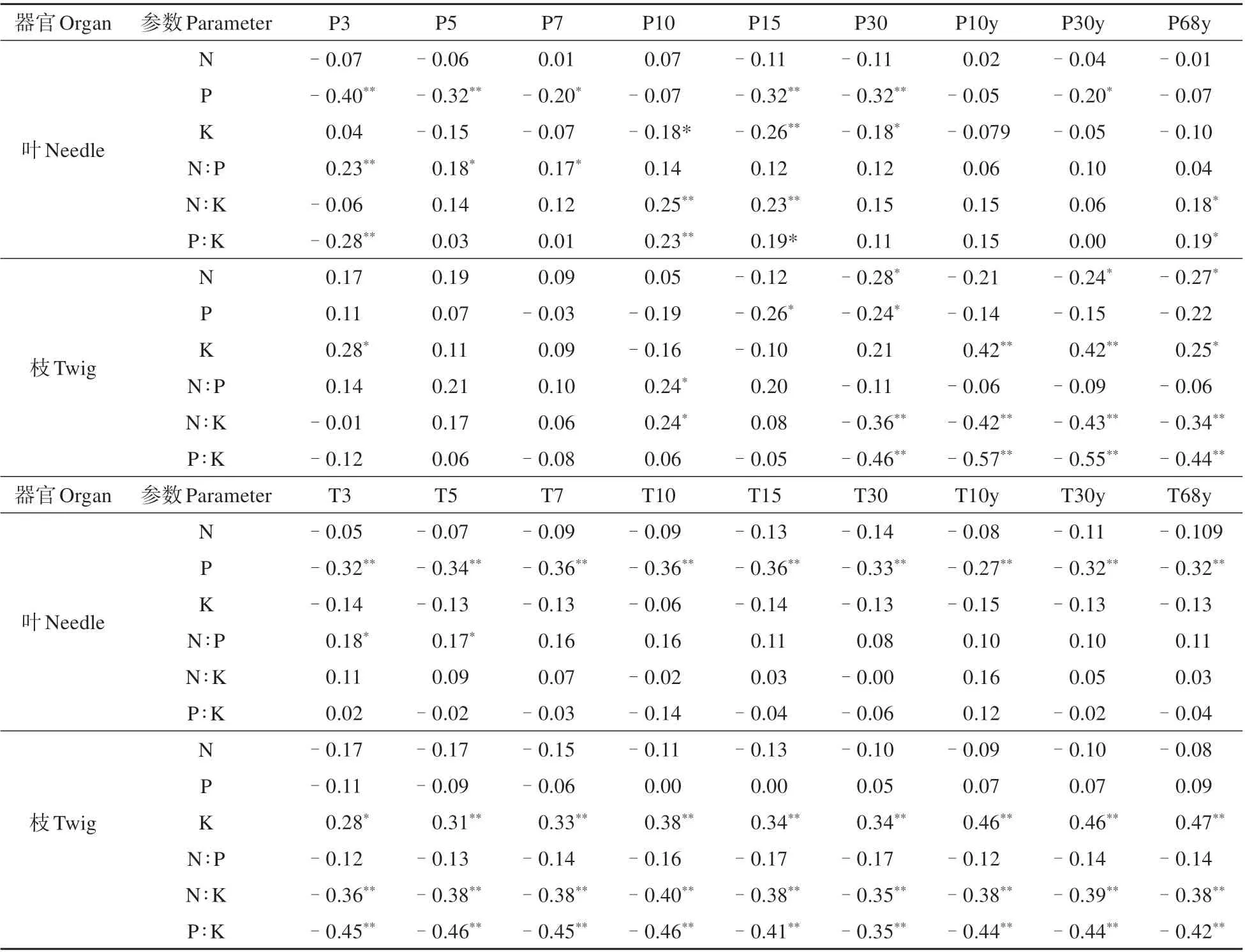
表6 大别山五针松枝叶N、P、K化学计量特征与温度和降水之间的相关系数Table 6 Correlation coefficients between N,P and K stoichiometric characteristics and temperature and precipitation of twigs and needles of P.dabeshanensis
枝和叶N、P 和K 化学计量比对温度和降水的响应有较大差异(见表6)。叶N∶P与短时间尺度3个降水量参数(P3、P5 和P7)显著正相关,与各温度参数均无显著相关性;除枝N∶P 与P10 显著相关外,叶和枝N:P 与其他任何时间尺度的温度和降水量均无显著相关性。叶N∶K 和P∶K 与P10、P15 及P68y 呈显著或极显著正相关,而与各温度参数均无显著相关性;枝N∶K 和P∶K 与4 个长时间尺度(P30 和3 个多年平均月降水量)降水量呈极显著负相关,而与9 个温度参数均无显著相关性。可见,大别山五针松N、P、K 化学计量指标之间以及枝和叶2 个器官之间对温度和降水变化的响应具有明显差异性。
3 讨论
3.1 大别山五针松枝叶N、P、K 含量及其化学计量比
与已有研究结果对比发现,大别山五针松针叶N 含量低于全球植被[21]、中国陆地植被[22]、亚热带(福建万木林和黔中地区)常绿植物和落叶植物的N 含量平均值[23~24](见表7)。相比于其他裸子植物,大别山五针松叶N 含量高于云南松(Pinus yunnanensis)[25]和山东黑松(Pinus thunbergii)[26],与湖南会同杉木(Cunninghamia lanceolata)[2]以及雪岭云杉(Picea schrenkiana)[27]接近,但低于浙江天童山马尾松(Pinus massoniana)[28]、三峡马尾松(Pinus massoniana)[29]、华北落叶松(Larix principis rupprechtii)[12]、广 西 大 青 山 杉 木(Cunninghamia lanceolata)[30]尤其是沙地樟子松(Pinus sylvestris var.mongolica)[31],体现出明显的种间差异性。大别山五针松枝条N 含量低于其针叶,也低于米槠(Castanopsis carlesii)[32]和 刺 槐(Robina pseudoacacia)[33]等被子植物,但高于杨梅(Myrica nana)[34];与其他裸子植物相比,大别山五针松枝N 含量远高于黔中亚热带针叶树种[35],但低于同气候区 的 杉 木(Cunninghamia lanceolata)[2]和 雪 岭云杉[27]。
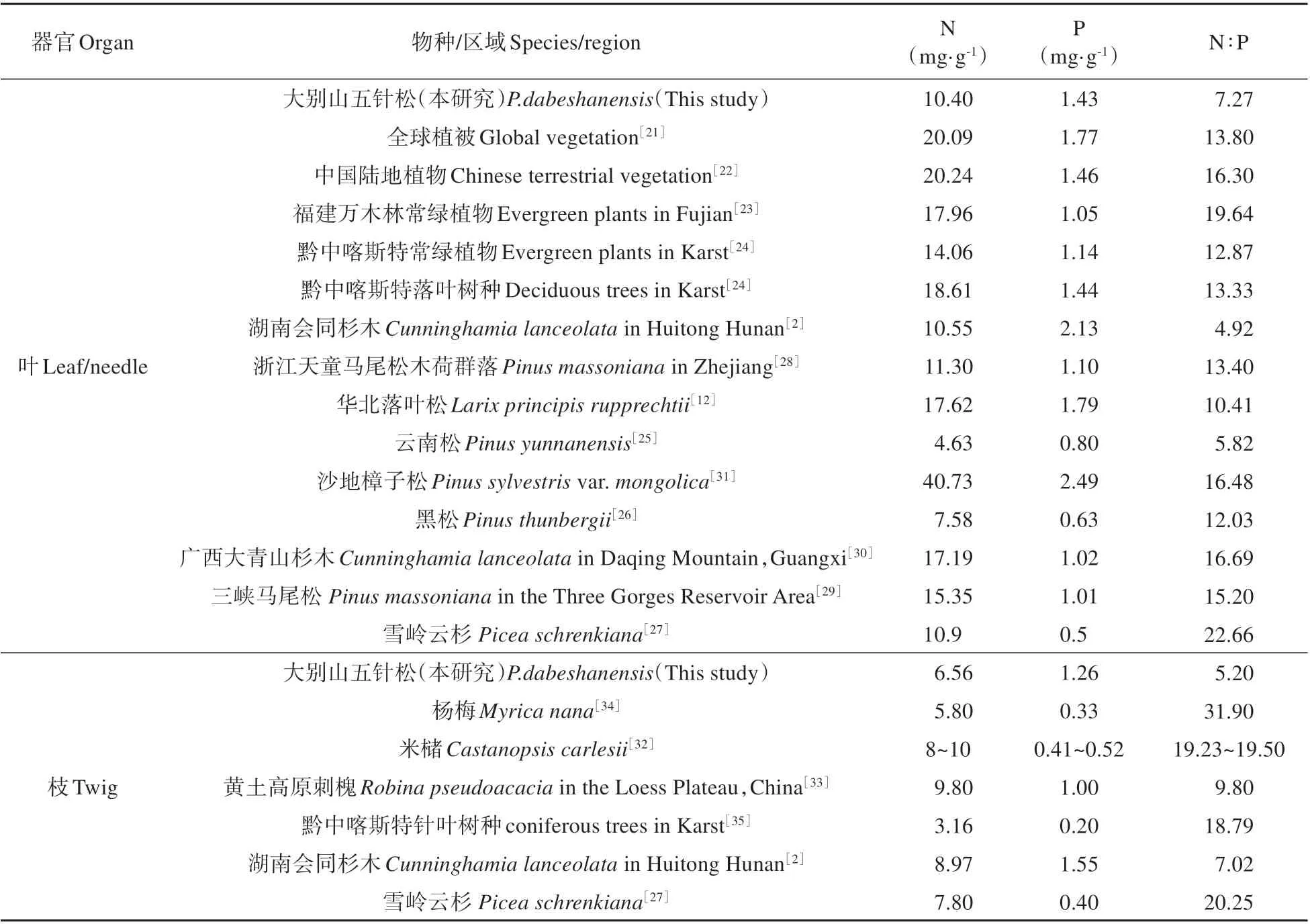
表7 大别山五针松N、P含量与其他研究对比Table 7 Comparison of N and P contents of P.dabeshanensis with other studies
对于叶P 含量而言,大别山五针松低于全球植被平均值[21],与我国陆地植被[22]和黔中亚热带落叶树种[24]含量接近,但高于福建及黔中分布的常绿植物[23-24](见表7)。在已知裸子植物中,大别山五针松针叶P 含量处于中等水平,其高于广西大青山杉木人工林[30]、山东黑松[26]、云南松[25]、浙江天童马尾松[28]、三峡马尾松[29]、雪岭云杉[27],但低于湖南会同杉木[2]、华北落叶松[12]和沙地樟子松[31]。此外,大别山五针松枝P 含量处于较高水平,仅低于湖南会同杉木[2]。
大别山五针松叶平均N:P(7.27)明显低于全球植被(13.8)[21]、中国陆地植物(16.3)[22]以及其他常绿和落叶树种的叶片N:P 平均值[23-24](见表7)。与其他裸子植物相比,大别山五针松叶N:P 高于会同杉木[2]和云南松[25],但低于多数研究结果,可能归因于前者较高的枝P 含量。N:P 通常作为植物养分限制的判断依据,即当叶N:P>16 时,植物生长表现为P 限制,当N:P<14 时,植物生长表现为N 限制,当16>N:P>14 时,为同时受N、P 限制或者N、P 均不限制[36]。大别山五针松叶平均N:P 远小于14,表现为严重的N 限制,这也是大部分裸子植物(沙地樟子松除外)所共有的特征(见表7)。
植物K 含量与植物的生长、抗旱以及抗病害能力有关,K 含量高的植物其抗性更强[37]。大别山五针松叶K 平均含量(5.15 mg·g-1)高于枝(4.80 mg·g-1),但低于黔中喀斯特植物叶片K 含量(7.22 mg·g-1)[24]。参考Venterink 等关于K 元素阈值的划分标准(N∶K>2.1,K∶P<3.4 时,植物的生长受K 元素限制)[38],大别山五针松针叶N∶K(2.02)<2.1 而K∶P(3.6)>3.4,表明其生长基本不受K的限制。尽管如此,大别山五针松相对较低的K 含量可能表明其面临外界环境胁迫时的抗性较低。
3.2 大别山五针松枝叶化学计量关系
尽管枝和叶N、P、K 含量随生长季有显著变化,但大别山五针松养分元素之间具有显著的相关性,且元素间的异速生长关系也较为稳定,这与多数研究结果相似,是植物种群稳定性维持的重要保证[1]。植物叶片N-P 间异速生长指数的理论值为2/3或者3/4[39],低于本研究中叶N-P异速生长指数1.61,表明大别山五针松叶具有更快的N 分配和积累速率。这可能是因为该值在不同植物功能群、不同科、不同地点和区域间本来就具有较大波动性(范围可达0.37~1.93),而全球植物的理论值是各类植物混合计算得到的,其弱化了类群间的差异性[6]。同时,本研究中N-P 异速生长指数也不支持针叶树种N 吸收利用率小于阔叶树种的结论[24]。另外,植物叶片N-P 异速生长指数存在明显的纬度格局,即热带最高,寒温带最低[6]。研究表明,中国57 种荒漠植物叶片N-P 异速生长指数为1.47[40],阿拉善荒漠草本植物为1.17[41],中国栎属植物(从北到南贯穿北温带和南亚热带)为1.88[42],本 地 香 樟 叶 片N-P 异 速 生 长 指 数 为1.48[43]。相比之下,大别山位于亚热带北缘,其较高的N-P 异速生长指数符合异速生长指数由热带向寒温带递减这一论断。
在裸子植物其他物种中,会同杉木叶N-P 异速生长指数(0.54)低于枝(0.62)[2],雪岭云杉叶NP 异速生长指数(0.96)为等速,而枝N-P 之间无显著异速生长关系[27]。此外,雪岭云杉针叶N-K、PK 及枝N-K 异速生长指数均为负(分别为-0.25、-1.57 和-0.33)[27],与大别山五针松N、P、K 间正的异速生长关系完全不同,体现了种间的差异性。
尽管枝和叶N-P 异速生长指数有差异,但枝和叶化学计量指标间的相关性相似。叶和枝中N∶P 都受到N 元素的控制,N∶K 和P∶K 主要受到K元素的控制。本研究与本地香樟[43]及湖南会同杉木[2]叶片N:P受到N控制的结论一致,但与福建万木林植物叶片N:P受到P控制不同[23]。
3.3 大别山五针松枝叶化学计量特征季节变化及其影响因素
植物在不同生长季其N、P、K 含量及计量比往往会有显著变化。研究发现侧柏在不同的生长期N∶P 保持稳定[15],体现了较强的内稳性。华北落叶松N∶P 随植物生长而先增加后下降[44],这是因为落叶植物生长旺盛期对养分需求量大导致N∶P升高,而生长后期生长速率减慢,进而导致N∶P 下降。杉木N∶P 随植物生长呈现不规则波动现象,其在10 月达到最低值[2]。本研究中大别山五针松样品采集地固定,标准木不变,也不存在空间异质性,因此器官和月份是其末端小枝化学计量特征变异的两个主要因素。云南普洱季风常绿阔叶林同一生境下植物体N、P、K含量及计量比主要受生长阶段的影响[45],而大别山五针松化学计量特征受到器官的影响最大,这种差异可能是由于生境及物种的不同引起的养分吸收和利用的差异性导致的。
大别山五针松针叶N、P 含量在整个生长季中会出现3 次先升高后降低的趋势,最后在10~11月含量再次增加,这与杉木叶片N、P 含量的变化基本相似[2]。针叶N、P、K 及N:P 在经过冬季停滞期后,于2 月迅速升高;可能是因为1 月末突然降温后,2月温度逐渐升高(见图1),植物生长发育逐渐恢复旺盛,进而导致N、P、K及N∶P逐渐升高。3~5月气温波动大且波动频繁,针叶养分含量又逐渐降低,其中N 在4 月最低,P 在4 月和5 月均较低,而K 在5 月最低。随着5 月后气温逐步提升且变幅变小,针叶养分含量升高且逐步趋于稳定。8月和9 月,针叶N、P、K 及枝N∶P 均明显降低,这可能是因为此时大别山五针松的生长基本停滞[46],植物不需要太多的营养元素。相比之下,针叶K 含量自2月开始下降,至5月到达最低,比N和P最低点有所延迟,这可能是金属与非金属元素的敏感性不同导致的。
温度对植物叶生理生态特征影响显著,主要是因为叶片薄且暴露在空中,温度变化会直接影响叶片。有研究发现,植物叶片N、P 含量与温度呈显著负相关,而N∶P 则随温度增加而增加,但也有研究认为植物N∶P 随温度增加而减少[47-49]。降水对植物化学元素的影响也随研究的时空尺度及研究对象的不同而呈现不同规律。有研究认为降水与N、P 呈显著负相关[50],有研究发现二者为正相关而与P 没有一致性关系[51],但也有研究认为降水对植物化学计量特征无显著影响[52]。本研究表明,大别山五针松叶N 与不同时间尺度的温度和降水均无显著相关性,而其N:P 则与短时间尺度的温度(3 和5 d)和降水(3、5 和7 d)有显著正相关;这表明,温度和降水在短时间内能显著影响大别山五针松N∶P,超过5 和7 d 后温度和降水的效应就减弱消失。除个别时间尺度降水量外,大别山五针松叶P 含量与各时间尺度下的温度和降水均为极显著负相关关系。这表明,降水会导致土壤淋溶增强、降低植物对P 的吸收[22,24],而高温不利于光合作用和植物生长,使得大量消耗P 的RNA 需求量降低[8,40],进而叶P 含量也显著降低。也符合全球植物P 含量由高纬度(低温)向低纬度(高温)递减的趋势[3]。因为依据“环境温度驱动植物氮磷计量关系”和“土壤母质养分条件决定氮磷限制”假说,随温度升高土壤N 和P 的可利用性降低,比如南方土壤普遍缺P[6]。
对于大别山五针松小枝而言,其K、N:K 和P:K 与各时间尺度下的温度均有显著相关性,即随温度升高枝的K 含量上升但其化学计量比下降。K 含量与植物抗性密切相关,温度较高的生长季有利于枝K 含量提高,对提升大别山五针松抗性有潜在益处[37]。降水对大别山五针松枝N、K 含量及N:P 和N:K 的影响具有时间尺度效应,表现为短时间尺度(1个月内)影响不显著,而长时间尺度(P30 及P10y、P30y 及P68y)影响显著。这可能是因为枝相比于叶,对环境的敏感性较低的缘故。
总的来说,降水和温度对大别山五针松不同生长期枝和叶化学计量特征具有一定影响。降水对叶和枝化学计量特征的影响具有不同的时间尺度效应,其中叶片主要受短时间尺度降水影响,而枝主要受长时间尺度降水影响。温度升高不利于大别山五针松叶P 积累,但有利于枝中K 含量提高,有助于提升枝条抗性。
4 结论
本研究表明,大别山五针松末端小枝N、P、K含量及其化学计量比的变异主要来源于器官,其次是月份。针叶养分元素含量明显高于枝;各化学计量指标的变异性多以中等偏弱为主(CV<25%)。枝或叶N、P、K 之间多具有显著的异速生长关系,枝和叶N-P及P-K 之间异速生长指数有显著差异,但枝和叶化学计量特征的相互关系类似,均表现为N∶P、N∶K 及P∶K 分别受N、K 和K 的控制。总体上,大别山五针松的生长表现出较严重的N限制,而P和K则相对充足。降水量对大别山五针松枝和叶化学计量特征的影响具有时间尺度效应,即短时间尺度影响大多数叶化学计量指标,而长时间尺度则显著影响枝化学计量特征。各时间尺度下的温度均显著影响针叶P 含量(负相关),但可以显著提升枝K 含量,并降低N∶K 和P∶K。
